
Assessing the Productivity of the Matang Mangrove Forest Reserve: Review of One of the Best-Managed Mangrove Forests

 , , , , , ,
, , , , , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Productivity
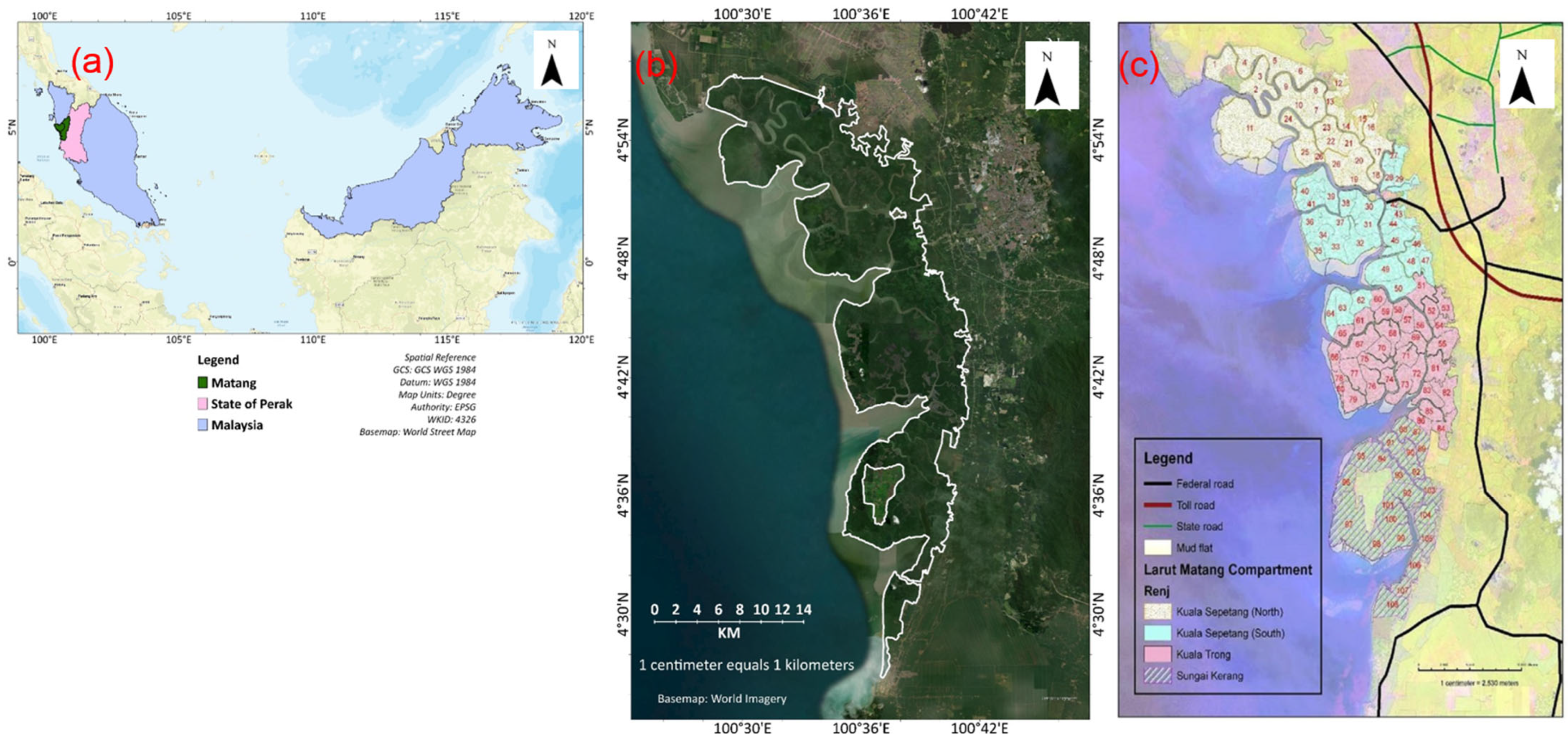
3. Study Area
- Avicennia-Sonneratia tree stands: These stands typically comprise young Avicennia trees that colonise new mud flats near river mouths. The dominant species in these stands are Avicennia alba Blume and A. officinalis L. However, patches of Sonneratia alba can also be found in A. alba and A. officinalis clumps. These stands are submerged during high tides and cover an area of 3299 hectares.
- Rhizophora apiculata tree stands: Found within the protected zone, these consist of Rhizophora apiculata and Rhizophora mucronata forms that are not subject to exploitation. While Rhizophora apiculata predominates, Rhizophora mucronata can also be found along estuaries and riverbanks. This stand covers an area of 1665 hectares.
- Dryland forest stands: These stands represent a transition to the local forest. They are characterised by a forest floor containing high densities of Acrostichum aureum Linnaeus and dry deciduous trees [42]. Dryland forest stands have 30 different tree species [45], including four main mangrove species such as stripe mangrove. These dryland forest stands are submerged by the equinoctial tides and are located at higher elevations inland. The total volume of dryland forests spans 2291 hectares [42].
4. Productivity of MMFR
5. Factors Affecting Productivity
5.1. Nutrients
5.2. The Gradient Effect of Flooding
5.3. Pollution
5.4. Climate Change
5.5. Management of Mangrove Forest
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Sahu, S.K.; Kathiresan, K. The age and species composition of mangrove forest directly influence the net primary productivity and carbon sequestration potential. Biocatal. Agric. Biotechnol. 2019, 20, 101235. [Google Scholar] [CrossRef]
- Kumari, P.; Singh, J.K.; Pathak, B. Potential contribution of multifunctional mangrove resources and its conservation. In Biotechnological Utilization of Mangrove Resources; Academic Press: Cambridge, MA, USA, 2020; pp. 1–26. [Google Scholar]
- Wang, Y.S.; Gu, J.D. Ecological responses, adaptation and mechanisms of mangrove wetland ecosystem to global climate change and anthropogenic activities. Int. Biodeterior. Biodegrad. 2021, 162, 105248. [Google Scholar] [CrossRef]
- Pradisty, N.A.; Amir, A.A.; Zimmer, M. Plant species-and stage-specific differences in microbial decay of mangrove leaf litter: The older the better? Oecologia 2021, 195, 843–858. [Google Scholar] [CrossRef]
- Chong, V.C. Sustainable utilization and management of mangrove ecosystems of Malaysia. Aquat. Ecosyst. Health Manag. 2006, 9, 249–260. [Google Scholar] [CrossRef]
- Soeprobowati, T.R.; Anggoro, S.; Puryono, S.; Purnaweni, H.; Sularto, R.B.; Mersyah, R. Species composition and distribution in the mangrove ecosystem in the city of Bengkulu, Indonesia. Water 2022, 14, 3516. [Google Scholar] [CrossRef]
- Praveena, S.M.; Radojevic, M.; Abdullah, M.H.; Aris, A.Z. Application of sediment quality guidelines in the assessment of mangrove surface sediment in Mengkabong lagoon, Sabah, Malaysia. J. Environ. Health Sci. Eng. 2008, 5, 35–42. [Google Scholar]
- Seca Gandaseca, S.G.; Noraini Rosli, N.R.; Johin Ngayop, J.N.; Arianto, C.I. Status of water quality based on the physico-chemical assessment on river water at Wildlife Sanctuary Sibuti Mangrove Forest, Miri Sarawak. Am. J. Environ. Sci. 2011, 7, 269–275. [Google Scholar] [CrossRef]
- Das, S. Evaluation of mangrove ecosystem services: Methodological and data challenges. In Energy, Environment and Globalization: Recent Trends, Opportunities and Challenges in India; Springer Nature: Berlin, Germany, 2020; pp. 157–174. [Google Scholar]
- Tengku Hashim, T.M.Z.; Suratman, M.N. Mangroves as a carbon sink/stocks. In Mangroves: Ecology, Biodiversity and Management; Springer Nature: Berlin, Germany, 2021; pp. 157–175. [Google Scholar]
- Alongi, D. The Energetics of Mangrove Forests; Springer Science & Business Media: Berlin, Germany, 2009. [Google Scholar]
- Mitra, A.; Mitra, A. Ecosystem services of mangroves: An overview. In Mangrove Forests in India: Exploring Ecosystem Services; Springer Nature: Berlin, Germany, 2020; pp. 1–32. [Google Scholar]
- Mumby, P.J.; Edwards, A.J.; Ernesto Arias-González, J.; Lindeman, K.C.; Blackwell, P.G.; Gall, A.; Gorczynska, M.I.; Harborne, A.R.; Pescod, C.L.; Renken, H.; et al. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 2004, 427, 533–536. [Google Scholar] [CrossRef]
- Laffoley, D.; Grimsditch, G.D. The Management of Natural Coastal Carbon Sinks; IUCN: Gland, Switzerland, 2009. [Google Scholar]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Dinilhuda, A.; Akbar, A.A.; Herawaty, H. Potentials of mangrove ecosystem as storage of carbon for global warming mitigation. Biodivers. J. Biol. Divers. 2020, 21. [Google Scholar] [CrossRef]
- Zhu, J.J.; Yan, B. Blue carbon sink function and carbon neutrality potential of mangroves. Sci. Total Environ. 2022, 822, 153438. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.C.; Pendleton, L.; Jenkins, W.A.; Sifleet, S. Green Payments for Blue Carbon: Economic Incentives for Protecting Threatened Coastal Habitats; CABI: Wallingford, UK, 2011. [Google Scholar]
- Khan, W.R.; Zulkifli, S.Z.; Kasim, M.R.B.M.; Pazi, A.M.; Mostapa, R.; Nazre, M. Mangrove Productivity Estimation using Mod-elling Approach and Tree Parameters Assessment. Trop. Conserv. Sci. 2019, 12, 1940082919872137. [Google Scholar]
- Ong, J.-E.; Gong, K.W.; Clough, B. Structure and productivity of a 20-year-old stand of Rhizophora apiculata Bl. mangrove forest. J. Biogeogr. 1995, 417–424. [Google Scholar]
- Alongi, D.M.; Sasekumar, A.; Chong, V.C.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Brunskill, G.J. Sediment accumulation and organic material flux in a managed mangrove ecosystem: Estimates of land–ocean–atmosphere exchange in peninsular Malaysia. Mar. Geol. 2004, 208, 383–402. [Google Scholar] [CrossRef]
- Friess, D.A.; Rogers, K.; Lovelock, C.E.; Krauss, K.W.; Hamilton, S.E.; Lee, S.Y.; Lucas, R.; Primavera, J.; Rajkaran, A.; Shi, S. The state of the world’s mangrove forests: Past, present, and future. Annu. Rev. Environ. Resour. 2019, 44, 89–115. [Google Scholar] [CrossRef]
- Odum, W.E.; McIvor, C.C.; Smith, T.J. The Ecology of the Mangroves of South Florida: A Community Profile; The Service; U.S. Department of the Interior: Washington, DC, USA, 1982.
- Twilley, R.R.; Day, J.W. The productivity and nutrient cycling of mangrove ecosystem. In Ecosistemas de manglar en América Tropical; Instituto de Ecología, AC: Xalapa, México; UICN/ORMA: San José, Costa Rica; NOAA/NMFS: Silver Spring, MD, USA, 1999; pp. 127–151. [Google Scholar]
- Krauss, K.W.; Lovelock, C.E.; McKee, K.L.; López-Hoffman, L.; Ewe, S.M.; Sousa, W.P. Environmental drivers in mangrove establishment and early development: A review. Aquat. Bot. 2008, 89, 105–127. [Google Scholar] [CrossRef]
- Kathiresan, K.; Moorthy, P.; Rajendran, N. Mangroves and its associate’s diversity. In Mangroves of Indiae Their Biology and Uses. Zoological Survey of India, Prani Vigyan Bhawan, MeBlock, New Alipore, Kol-kata; Zoological Survey of India: Kolkata, India, 2013; Volume 700, p. 2013. [Google Scholar]
- Collins, D.S.; Avdis, A.; Allison, P.A.; Johnson, H.D.; Hill, J.; Piggott, M.D.; Hassan, M.H.A.; Damit, A.R. Tidal dynamics and mangrove carbon sequestration during the Oligo–Miocene in the South China Sea. Nat. Commun. 2017, 8, 15698. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. Climate Regulation by Capturing Carbon in Mangroves; The Wetland Book; Springer: Dordrecht, Switzerland, 2016; pp. 1–7. [Google Scholar]
- Odum, W.E.; Heald, E.J. The detritus-based food web of an estuarine mangrove community. In Estuarine Research: Chemistry, Biology, and the Estuarine System; Academic Press: Cambridge, MA, USA, 1975; Volume 1, p. 265. [Google Scholar]
- Matsui, N. Estimated stocks of organic carbon in mangrove roots and sediments in Hinchinbrook Channel, Australia. Mangroves Salt Marshes 1998, 2, 199–204. [Google Scholar] [CrossRef]
- Sahu, S.K.; Singh, R.; Kathiresan, K. Multi-gene phylogenetic analysis reveals the multiple origin and evolution of mangrove physiological traits through exaptation. Estuar. Coast. Shelf Sci. 2016, 183, 41–51. [Google Scholar] [CrossRef]
- Amarasinghe, M.D.; Balasubramaniam, S. Net primary productivity of two mangrove forest stands on the northwestern coast of Sri Lanka. Hydrobiologia 1992, 247, 37–47. [Google Scholar] [CrossRef]
- Mehmood, K.; Anees, S.A.; Luo, M.; Akram, M.; Zubair, M.; Khan, K.A.; Khan, W.R. Assessing Chilgoza Pine (Pinus gerardiana) Forest Fire Severity: Remote Sensing Analysis, Correlations, and Predictive Modeling for Enhanced Management Strategies. Trees For. People 2024, 16, 100521. [Google Scholar] [CrossRef]
- Mehmood, K.; Anees, S.A.; Rehman, A.; Tariq, A.; Zubair, M.; Liu, Q.; Rabbi, F.; Khan, K.A.; Luo, M. Exploring spatiotemporal dynamics of NDVI and climate-driven responses in ecosystems: Insights for sustainable management and climate resilience. Ecol. Inform. 2024, 80, 102532. [Google Scholar] [CrossRef]
- Okimoto, Y.; Nose, A.; Katsuta, Y.; Tateda, Y.; Agarie, S.; Ikeda, K. Gas exchange analysis for estimating net CO2 fixation capacity of mangrove (Rhizophora stylosa) forest in the mouth of river Fukido, Ishigaki Island, Japan. Plant Prod. Sci. 2007, 10, 303–313. [Google Scholar] [CrossRef]
- Okamura, K.; Tanaka, K.; Siow, R.; Man, A.; Kodama, M.; Ichikawa, T. Spring tide hypoxia with relation to chemical properties of the sediments in the Matang mangrove estuary, Malaysia. Jpn. Agric. Res. Q. 2010, 44, 325–333. [Google Scholar] [CrossRef]
- Muda, A. A Working Plan for the Matang Mangrove Forest Reserves, Perak: The Third 10-Year Period (2000–2009) of the Second Rotation; State Foresty Department of Perak Darul Rizuan: Ipoh, Malaysia, 2003. [Google Scholar]
- Khan, W.R.; Aljahdali, M.O. Elemental composition of above and belowground mangrove tissue and sediment in managed and unmanaged compartments of the Matang Mangrove Forest Reserve. Plants 2022, 11, 2916. [Google Scholar] [CrossRef] [PubMed]
- Dahlan, Z.A.; Othman, N.A.; Idris, M.F.I.M.; Kamaruzaman, J.H. Development of Individual Learners: Perspective on the Uncertain Future Contribution of E-Learning. Comput. Inf. Sci. 2009, 2, 182–189. [Google Scholar] [CrossRef]
- Malaysian Meteorological Department. Available online: https://www.met.gov.my/en/penerbitan/laporan-tahunan/ (accessed on 28 September 2017).
- Chan, H.T. A note on tree species and productivity of a natural dryland mangrove forest in Matang, Peninsular Malaysia. J. Trop. For. Sci. 1989, 1, 399–400. [Google Scholar]
- Ariffin, R.; Mustafa, N.M.S.N. A Working Plan for the Matang Mangrove Forest Reserve, Perak; State Forestry Department of Perak: Ipoh, Malaysia, 2013. [Google Scholar]
- Chen, D.; Satyanarayana, B.; Wolswijk, G.; Abd Rahim, N.H.; Amir, A.A.; Hugé, J.; Dahdouh-Guebas, F. Long-term monitoring and appraisal of forest management for extractive uses and other ecological values–The case of Matang mangrove forest Reserve, Malaysia. J. Nat. Conserv. 2024, 126582. [Google Scholar] [CrossRef]
- National Hydrography Centre Malaysia. Malaysian Tide Table 2017; Royal Malaysian Navy: Lumut, Malaysia, 2017. [Google Scholar]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Shaharuddin, M.I.; Azahar, M.; Razani, U.; Kamaruzaman, A.B.; Lim, K.L.; Rosli, S.; Jalil Md, S.; Latiff, A. Sustainable Management of Matang Mangroves: 100 Years and Beyond; Forestry Department Peninsular Malaysia: Kuala Lumpur, Malaysia, 2005.
- Ong, J.E. Mangroves-a carbon source and sink. Chemosphere 1993, 27, 1097–1107. [Google Scholar] [CrossRef]
- Ammar, A.A.; Dargusch, P.; Shamsudin, I. Can the Matang Mangrove Forest Reserve provide perfect teething ground for a blue carbon based REDD+ pilot project? J. Trop. For. Sci. 2014, 26, 371–381. [Google Scholar]
- Sioh, M. Colonial Trauma and Postcolonial Anxieties: The Haunted Choices of Economic Development; Taylor & Francis: Abingdon-on-Thames, UK, 2024. [Google Scholar]
- Gong, W.K.; Ong, J.E. Plant biomass and nutrient flux in a managed mangrove forest in Malaysia. Estuar. Coast. Shelf Sci. 1990, 31, 519–530. [Google Scholar] [CrossRef]
- Goessens, A.; Satyanarayana, B.; Van der Stocken, T.; Quispe Zuniga, M.; Mohd-Lokman, H.; Sulong, I.; Dahdouh-Guebas, F. Is Matang Mangrove Forest in Malaysia sustainably rejuvenating after more than a century of conservation and harvesting management? PLoS ONE 2014, 9, e105069. [Google Scholar] [CrossRef]
- Putz, F.E.; Chan, H.T. Tree growth, dynamics, and productivity in a mature mangrove forest in Malaysia. For. Ecol. Manag. 1986, 17, 211–230. [Google Scholar] [CrossRef]
- Khoon, G.W.; Eong, O.J. The use of demographic studies in mangrove silviculture. In Asia-Pacific Symposium on Mangrove Ecosystems: Proceedings of the International Conference Held at The Hong Kong University of Science & Technology, Hong Kong, 1–3 September 1993; Springer: Dordrecht, The Netherlands, 1995; pp. 255–261. [Google Scholar]
- Ong, J.E.; Gong, W.K.; Wong, C.H. Seven years of productivity studies in a Malaysian managed mangrove forest then what. In Coastal and Tidal Wetlands of the Australian Monsoon Region; National Library of Australia: Canberra, Australia, 1985; pp. 213–223. [Google Scholar]
- Adame, M.F.; Zakaria, R.M.; Fry, B.; Chong, V.C.; Then, Y.H.A.; Brown, C.J.; Lee, S.Y. Loss and recovery of carbon and nitrogen after mangrove clearing. Ocean. Coast. Manag. 2018, 161, 117–126. [Google Scholar] [CrossRef]
- Hamdan, O.; Khairunnisa, M.R.; Ammar, A.A.; Hasmadi, I.M.; Aziz, H.K. Mangrove carbon stock assessment by optical satellite imagery. J. Trop. For. Sci. 2013, 25, 554–565. [Google Scholar]
- Otero, V.; Van De Kerchove, R.; Satyanarayana, B.; Martínez-Espinosa, C.; Fisol, M.A.B.; Ibrahim, M.R.B.; Sulong, I.; Mohd-Lokman, H.; Lucas, R.; Dahdouh-Guebas, F. Managing mangrove forests from the sky: Forest inventory using field data and Unmanned Aerial Vehicle (UAV) imagery in the Matang Mangrove Forest Reserve, peninsular Malaysia. For. Ecol. Manag. 2018, 411, 35–45. [Google Scholar] [CrossRef]
- Veettil, B.K.; Ward, R.D.; Quang, N.X.; Trang, N.T.T.; Giang, T.H. Mangroves of Vietnam: Historical development, current state of research and future threats. Estuar. Coast. Shelf Sci. 2019, 218, 212–236. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Kangkuso, A.; Jamili, J.; Septiana, A.; Raya, R.; Sahidin, I.; Rianse, U.; Rahim, S.; Alfirman, A.; Sharma, S.; Nadaoka, K. Allometric models and aboveground biomass of Lumnitzera racemosa Willd. forest in Rawa Aopa Watumohai National Park, Southeast Sulawesi, Indonesia. For. Sci. Technol. 2016, 12, 43–50. [Google Scholar]
- Boto, K.G.; Wellington, J.T. Soil characteristics and nutrient status in a northern Australian mangrove forest. Estuaries 1984, 7, 61–69. [Google Scholar] [CrossRef]
- Guannel, G.; Arkema, K.; Ruggiero, P.; Verutes, G. The power of three: Coral reefs, seagrasses and mangroves protect coastal regions and increase their resilience. PLoS ONE 2016, 11, e0158094. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, J.; Spalding, M.; Zu Ermgassen, P. The role of mangroves in fisheries enhancement. Nat. Conserv. Wetl. Int. 2014, 54, 434. [Google Scholar]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Snedaker, S.C.; Lahmann, E.J. Mangrove understorey absence: A consequence of evolution? J. Trop. Ecol. 1988, 4, 311–314. [Google Scholar] [CrossRef]
- Bayen, S. Occurrence, bioavailability and toxic effects of trace metals and organic contaminants in mangrove ecosystems: A review. Environ. Int. 2012, 48, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Akhand, A.; Chanda, A.; Dutta, S.; Hazra, S.; Sanyal, P. Comparative study of heavy metáis in selected mangroves of Sundarban ecosystem, India. J. Environ. Biol. 2012, 33, 1045–1049. [Google Scholar] [PubMed]
- MacFarlane, G.R.; Pulkownik, A.; Burchett, M.D. Accumulation and distribution of heavy metals in the grey mangrove, Avicennia marina (Forsk.) Vierh.: Biological indication potential. Environ. Pollut. 2003, 123, 139–151. [Google Scholar] [CrossRef]
- Ismail, A.; Jusoh, N.R.; Ghani, I.A. Trace metal concentrations in marine prawns off the Malaysian coast. Mar. Pollut. Bull. 1995, 31, 108–110. [Google Scholar] [CrossRef]
- Rahman, F.; Ismail, A.; Yusof, S. Metals contamination in the foraging area of Milky Stork: Evidence of anthropogenic inputs in the aquatic environment of Kuala Gula, Malaysia. Toxicol. Environ. Chem. 2013, 95, 1499–1505. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Kellner, J.R.; Forde, A.J.; Gruner, D.S.; Parker, J.D.; Rodriguez, W.; Feller, I.C. Poleward expansion of mangroves is a threshold response to decreased frequency of extreme cold events. Proc. Natl. Acad. Sci. USA 2014, 111, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Cunha-Lignon, M.; Kampel, M.; Menghini, R.P.; Schaeffer-Novelli, Y.; Cintrón, G.; Dahdouh-Guebas, F. Mangrove forests submitted to depositional processes and salinity variation investigated using satellite images and vegetation structure surveys. J. Coast. Res. 2011, 344–348. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Range | Compartment | Protective Zone (ha) | Restrictive Zone (ha) | Productive Zone (ha) | Unproductive Zone (ha) | Total (ha) |
|---|---|---|---|---|---|---|
| Kuala Sepetang | 1–50 62–64 | 5164 | 1369 | 14,357 | 179 | 21,069 |
| Kuala Trong | 51–61 65–86 | 834 | 480 | 9560 | 84 | 10,958 |
| Sungai Kerang | 87–108 | 1362 | 1043 | 5877 | 157 | 8439 |
| Total | 7360 (18.2%) | 2892 (7.2%) | 29,794 (73.6%) | 420 (1%) | 40,466 (100%) |
| Years | Age | Species | ABG t/ha | Sources |
|---|---|---|---|---|
| 1950 | - | Rhizophora apiculata | 270 | [53] |
| 1980 | 5 | Rhizophora apiculata | 72 | [54] |
| 1980 | 13 | Rhizophora apiculata | 131 | [54] |
| 1980 | 23 | Rhizophora apiculata | 155 | [54] |
| 1980 | 28 | Rhizophora apiculata | 153 | [54] |
| 1981 | Seedling to mature | Rhizophora apiculata | 460 | [53] |
| 1984 | 1–10 (5) | Rhizophora apiculata | 100 | [55] |
| 1984 | 11–20 (10) | Rhizophora apiculata | 200 | [55] |
| 1984 | 21–30 (15) | Rhizophora apiculata | 300 | [55] |
| 1985 | 10 | Rhizophora apiculata | 18 | [55] |
| 1985 | 15 | Rhizophora apiculata | 13 | [55] |
| 1985 | 25 | Rhizophora apiculata | 12 | [55] |
| 1986 | >80 | Rhizophora apiculata | 270 | [53] |
| 1986 | >80 | Rhizophora apiculata | 460 | [53] |
| 1986 | >80 | Rhizophora apiculata | 400 | [53] |
| 1986 | 30 | Rhizophora apiculata | 150 | [53] |
| 1986 (1950–1981) | Seedling to mature | Rhizophora apiculata | 409 averages | [53] |
| 1990 | - | Rhizophora apiculata | 185.30 | [51] |
| 1990 | 1–30 | Rhizophora apiculata | 202.53 | [51] |
| 1993 | 5 | Rhizophora apiculata | 8 | [54] |
| 1993 | 10 | Rhizophora apiculata | 90 | [54] |
| 1993 | 15 | Rhizophora apiculata | 100 | [54] |
| 1993 | 25 | Rhizophora apiculata | 150 | [54] |
| 1993 | 30 | Rhizophora apiculata | 300 | [54] |
| 1995 | 20 | Rhizophora apiculata | 228 | [54] |
| 2011 | 15 | Rhizophora apiculata | 216 | [52] |
| 2011 | 20 | Rhizophora apiculata | 217 | [52] |
| 2011 | 30 | Rhizophora apiculata | 372 | [52] |
| 2014 | >80 | Rhizophora apiculata Rhizophora mucronata Bruguiera parviflora Avicennia officinalis Sonneratia alba Excoecaria agallocha Bruguiera gymnorrhiza Bruguiera cylindrical | 415 | [52] |
| 2014 | 15 | Rhizophora apiculata | 216 | [52] |
| 2014 | 20 | Rhizophora apiculata | 217 | [52] |
| 2014 | 30 | Rhizophora apiculata | 372 | [52] |
| 2016 (1980–1989) | - | All species in the area | 177 | [56] |
| 2016 (1990–1999) | - | All species in the area | 175 | [56] |
| 2016 (2000–2009) | - | All species in the area | 179 | [56] |
| 2019 | 15 | Rhizophora apiculata | 168 | [20] |
| 2019 | 15 | Rhizophora apiculata | 235 | [20] |
| 2019 | 25 | Rhizophora apiculata | 241 | [20] |
| 2019 | 25 | Rhizophora apiculata | 283 | [20] |
| 2019 | 30 | Rhizophora apiculata | 266 | [20] |
| Through remote sensing base | ||||
| 1991 | All age class species | All species in MMFR | 153 t/ha | [57] |
| 2011 | All age class species | All species in MMFR | 140 t/ha | [57] |
| 2017 | 90 (protective forest) | All species in MMFR | 210 Mg ha−1 UAV data | [58] |
| 2017 | 90 (protective forest) | All species in MMFR | 143 Mg ha−1 ground inventory data | [58] |
| 2017 | 15 productive forests | All species in MMFR | 217 Mg ha−1 UAV data | [58] |
| 2017 | 15 productive forests | All species in MMFR | 238 Mg ha−1 ground inventory data | [58] |
| Years | Difference between ABG t/ha | Estimated ABG t/ha Difference per/year | Indication Increase/Decrease |
|---|---|---|---|
| 1950–1980 | 270–153 | 117 | Decreased |
| 1980–1981 | 153–460 | 307 | Increased |
| 1981–1984 | 460–300 | 160 | Decreased |
| 1984–1986 | 300–400 | 100 | Increased |
| 1986–1990 | 400–202 | 198 | Decreased |
| 1990–1993 | 202–300 | 98 | Increased |
| 1993–1995 | 300–228 | 72 | Decreased |
| 1995–2011 | 228–372 | 144 | Increased |
| 2011–2014 | 372–415 | 43 | Increased |
| 2014–2016 | 372–177 | 195 | Decreased |
| 2016–2019 | 177–168 | 9 | Same |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, W.R.; Nazre, M.; Akram, S.; Anees, S.A.; Mehmood, K.; Ibrahim, F.H.; Edrus, S.S.O.A.; Latiff, A.; Fitri, Z.A.; Yaseen, M.; et al. Assessing the Productivity of the Matang Mangrove Forest Reserve: Review of One of the Best-Managed Mangrove Forests. Forests 2024, 15, 747. https://doi.org/10.3390/f15050747
Khan WR, Nazre M, Akram S, Anees SA, Mehmood K, Ibrahim FH, Edrus SSOA, Latiff A, Fitri ZA, Yaseen M, et al. Assessing the Productivity of the Matang Mangrove Forest Reserve: Review of One of the Best-Managed Mangrove Forests. Forests. 2024; 15(5):747. https://doi.org/10.3390/f15050747
Chicago/Turabian StyleKhan, Waseem Razzaq, Mohammad Nazre, Seemab Akram, Shoaib Ahmad Anees, Kaleem Mehmood, Faridah Hanum Ibrahim, Syeed SaifulAzry Osman Al Edrus, Abdul Latiff, Zohari Ahmad Fitri, Muhammad Yaseen, and et al. 2024. "Assessing the Productivity of the Matang Mangrove Forest Reserve: Review of One of the Best-Managed Mangrove Forests" Forests 15, no. 5: 747. https://doi.org/10.3390/f15050747