

Colletotrichum Species Causing Cyclocarya paliurus Anthracnose in Southern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Molecular Identification
2.2.1. DNA Extraction
2.2.2. Multigene Amplification and Sequencing
2.2.3. Phylogenetic Analyses
2.3. Phenotypic Analysis
2.4. Pathogenicity Tests
2.5. Data Analyses
3. Results
3.1. Symptomatology and Fungal Isolation
3.2. Molecular Identification and Phylogenetic Analyses
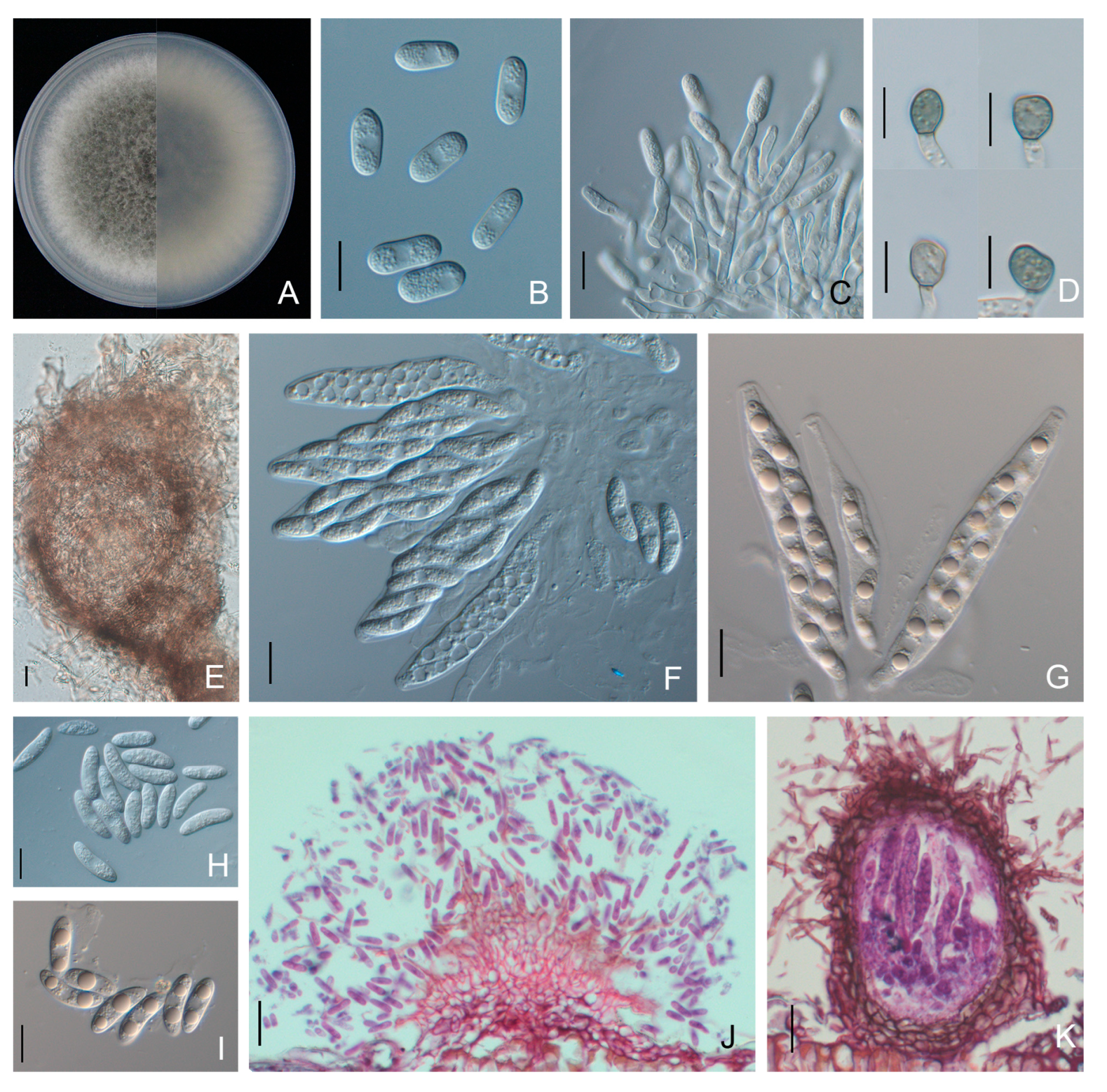
3.3. Morphological Characteristics
3.4. Pathogenicity Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Culture/Isolate a | Host | Location | GenBank Accession Number b | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ITS | GAPDH | CHS | ACT | TUB | CAL | GS | ApMat | ||||
| Colletotrichum aenigma | ICMP 18608 | Persea americana | Israel | JX010244 | JX010044 | JX009774 | JX009443 | JX010389 | JX009683 | JX010078 | KM360143 |
| C. aeschynomenes | ICMP 17673 | Aeschynomene virginica | USA | JX010176 | JX009930 | JX009799 | JX009483 | JX010392 | JX009721 | JX010081 | KM360145 |
| C. alatae | CBS 304.67 | Dioscorea alata | India | JX010190 | JX009990 | JX009837 | JX009471 | JX010383 | JX009738 | JX010065 | KC888932 |
| C. alienum | ICMP 12071 | Malus domestica | New Zealand | JX010251 | JX010028 | JX009882 | JX009572 | JX010411 | JX009654 | JX010101 | KM360144 |
| C. aotearoa | ICMP 18537 | Coprosma sp. | New Zealand | JX010205 | JX010005 | JX009853 | JX009564 | JX010420 | JX009611 | JX010113 | KC888930 |
| C. asianum | CBS 130418 | Coffea arabica | Thailand | FJ972612 | JX010053 | JX009867 | JX009584 | JX010406 | FJ917506 | JX010096 | FR718814 |
| C. camelliae | CGMCC 3.14925 | Camellia sinensis | China | KJ955081 | KJ954782 | MZ799255 | KJ954363 | KJ955230 | KJ954634 | KJ954932 | KJ954497 |
| C. chrysophilum | CMM 4268 | Musa sp. | Brazil | KX094252 | KX094183 | KX094083 | KX093982 | KX094285 | KX094063 | KX094204 | KX094325 |
| C. clidemiae | ICMP 18658 | Clidemia hirta | USA | JX010265 | JX009989 | JX009877 | JX009537 | JX010438 | JX009645 | JX010129 | KC888929 |
| C. conoides | CGMCC 3.17615 | Chili pepper | China | KP890168 | KP890162 | KP890156 | KP890144 | KP890174 | KP890150 | - | - |
| C. cordylinicola | ICMP 18579 | Cordyline fruticosa | Thailand | JX010226 | JX009975 | JX009864 | HM470235 | JX010440 | HM470238 | JX010122 | JQ899274 |
| C. fructicola | CBS 130416 | Coffea arabica | Thailand | JX010165 | JX010033 | JX009866 | FJ907426 | JX010405 | FJ917508 | JX010095 | JQ807838 |
| NFUCF-4 | Cyclocarya paliurus | Sichuan, China | OR056200 | OR069484 | OR073817 | OR096449 | OR073835 | OR096522 | OR098645 | OR105821 | |
| NFUCF-5 | Cy. paliurus | Guizhou, China | OR056201 | OR069485 | OR073818 | OR096450 | OR073836 | OR096523 | OR098646 | OR105822 | |
| NFUCF-12 | Cy. paliurus | Sichuan, China | OR056202 | OR069486 | OR073819 | OR096451 | OR073837 | OR096524 | OR098647 | OR105823 | |
| NFUCF-15 c | Cy. paliurus | Guangxi, China | OR056203 | OR069487 | OR073820 | OR096452 | OR073838 | OR096525 | OR098648 | OR105824 | |
| NFUCF-21 | Cy. paliurus | Fujian, China | OR056204 | OR069488 | OR073821 | OR096453 | OR073839 | OR096526 | OR098649 | OR105825 | |
| NFUCF-28 | Cy. paliurus | Hubei, Chna | OR056205 | OR069489 | OR073822 | OR096454 | OR073840 | OR096527 | OR098650 | OR105826 | |
| NFUCF-35 | Cy. paliurus | Jiangxi, China | OR056206 | OR069490 | OR073823 | OR096455 | OR073841 | OR096528 | OR098651 | OR105827 | |
| NFUCF-41 | Cy. paliurus | Guangxi, China | OR056207 | OR069491 | OR073824 | OR096456 | OR073842 | OR096529 | OR098652 | OR105828 | |
| NFUCF-43 | Cy. paliurus | Fujian, China | OR056208 | OR069492 | OR073825 | OR096457 | OR073843 | OR096530 | OR098653 | OR105829 | |
| NFUCF-51 | Cy. paliurus | Jiangxi, China | OR056209 | OR069493 | OR073826 | OR096458 | OR073844 | OR096531 | OR098654 | OR105830 | |
| NFUCF-59 | Cy. paliurus | Guizhou, China | OR056210 | OR069494 | OR073827 | OR096459 | OR073845 | OR096532 | OR098655 | OR105831 | |
| NFUCF-62 c | Cy. paliurus | Hunan, China | OR056211 | OR069495 | OR073828 | OR096460 | OR073846 | OR096533 | OR098656 | OR105832 | |
| NFUCF-74 | Cy. paliurus | Guangxi, China | OR056212 | OR069496 | OR073829 | OR096461 | OR073847 | OR096534 | OR098657 | OR105833 | |
| NFUCF-95 | Cy. paliurus | Zhejiang, China | OR056213 | OR069497 | OR073830 | OR096462 | OR073848 | OR096535 | OR098658 | OR105834 | |
| NFUCF-118 | Cy. paliurus | Guizhou, China | OR056214 | OR069498 | OR073831 | OR096463 | OR073849 | OR096536 | OR098659 | OR105835 | |
| NFUCF-154 | Cy. paliurus | Hunan, China | OR056215 | OR069499 | OR073832 | OR096464 | OR073850 | OR096537 | OR098660 | OR105836 | |
| NFUCF-179 | Cy. paliurus | Jiangxi, China | OR056216 | OR069500 | OR073833 | OR096465 | OR073851 | OR096538 | OR098661 | OR105837 | |
| NFUCF-214 c | Cy. paliurus | Zhejiang, China | OR056217 | OR069501 | OR073834 | OR096466 | OR073852 | OR096539 | OR098662 | OR105838 | |
| C. gloeosporioides | CBS 112999 | Citrus sinensis | Italy | JX010152 | JX010056 | JX009818 | JX009531 | JX010445 | JX009731 | JX010085 | JQ807843 |
| NFUCl-3 | Cy. paliurus | Guangxi, China | OR064046 | OR069502 | OR073853 | OR096419 | OR096467 | OR096540 | OR098663 | OR105839 | |
| NFUCl-5 c | Cy. paliurus | Jiangxi, China | OR064047 | OR069503 | OR073854 | OR096420 | OR096468 | OR096541 | OR098664 | OR105840 | |
| NFUCl-11 c | Cy. paliurus | Guizhou, China | OR064048 | OR069504 | OR073855 | OR096421 | OR096469 | OR096542 | OR098665 | OR105841 | |
| NFUCl-28 | Cy. paliurus | Guizhou, China | OR064049 | OR069505 | OR073856 | OR096422 | OR096470 | OR096543 | OR098666 | OR105842 | |
| NFUCl-34 c | Cy. paliurus | Hunan, China | OR064050 | OR069506 | OR073857 | OR096423 | OR096471 | OR096544 | OR098667 | OR105843 | |
| C. grevilleae | CBS 132879 | Grevillea sp. | Italy | KC297078 | KC297010 | KC296987 | KC296941 | KC297102 | KC296963 | KC297033 | - |
| C. hebeiense | MFLUCC13–0726 | Vitis vinifera | China | KF156863 | KF377495 | KF289008 | KF377532 | KF288975 | - | - | - |
| C. henanense | CGMCC 3.17354 | Ca. sinensis | China | KJ955109 | KJ954810 | MZ799256 | KM023257 | KJ955257 | KJ954662 | KJ954960 | KJ954524 |
| C. horii | ICMP 10492 | Diospyros kaki | Japan | GQ329690 | GQ329681 | JX009752 | JX009438 | JX010450 | JX009604 | JX010137 | JQ807840 |
| C. jiangxiense | CGMCC 3.17361 | Ca. sinensis | China | KJ955149 | KJ954850 | MZ799257 | KJ954427 | OK236389 | KJ954701 | KJ955000 | KJ954561 |
| C. kahawae subsp. ciggaro | ICMP 18539 | Olea europaea | Australia | JX010230 | JX009966 | JX009800 | JX009523 | JX010434 | JX009635 | JX010132 | - |
| C. musae | CBS 116870 | Musa sp. | USA | JX010146 | JX010050 | JX009896 | JX009433 | HQ596280 | JX009742 | JX010103 | KC888926 |
| C. nupharicola | CBS 470.96 | Nuphar lutea | USA | JX010187 | JX009972 | JX009835 | JX009437 | JX010398 | JX009663 | JX010088 | JX145319 |
| C. perseae | CBS 141365 | Avocado | Israel | KX620308 | KX620242 | MZ799260 | KX620145 | KX620341 | KX620206 | KX620275 | KX620177 |
| C. proteae | CBS 132882 | Protea sp. | South Africa | KC297079 | KC297009 | KC296986 | KC296940 | KC297101 | KC296960 | KC297032 | - |
| C. psidii | CBS 145.29 | Psidium sp. | Italy | JX010219 | JX009967 | JX009901 | JX009515 | JX010443 | JX009743 | JX010133 | KC888931 |
| C. queenslandicum | ICMP 1778 | Carica papaya | Australia | JX010276 | JX009934 | JX009899 | JX009447 | JX010414 | JX009691 | JX010104 | KC888928 |
| C. salsolae | ICMP 19051 | Salsola tragus | Hungary | JX010242 | JX009916 | JX009863 | JX009562 | JX010403 | JX009696 | JX010093 | KC888925 |
| C. siamense | CBS 130417 | Coffea arabica | Thailand | JX010171 | JX009924 | JX009865 | FJ907423 | JX010404 | FJ917505 | JX010094 | JQ899289 |
| C. tainanense | CBS 143666 | Capsicum annuum | China | MH728818 | MH728823 | MH805845 | MH781475 | MH846558 | - | MH748259 | MH728836 |
| C. theobromicola | CBS 124945 | Theobroma cacao | Panama | JX010294 | JX010006 | JX009869 | JX009444 | JX010447 | JX009591 | JX010139 | KC790726 |
| C. ti | ICMP 4832 | Cordyline sp. | New Zealand | JX010269 | JX009952 | JX009898 | JX009520 | JX010442 | JX009649 | JX010123 | KM360146 |
| C. tropicale | CBS 124949 | Theobroma cacao | Panama | JX010264 | JX010007 | JX009870 | JX009489 | JX010407 | JX009719 | JX010097 | KC790728 |
| C. wuxiense | CGMCC 3.17894 | Camellia sinensis | China | KU251591 | KU252045 | KU251939 | KU251672 | KU252200 | KU251833 | KU252101 | KU251722 |
| C. xanthorrhoeae | CBS 127831 | Xanthorrhoea preissii | Australia | JX010261 | JX009927 | JX009823 | JX009478 | JX010448 | JX009653 | JX010138 | KC790689 |
| C. yulongense | CFCC 50818 | Vaccinium dunalianum | China | MH751507 | MK108986 | MH793605 | MH777394 | MK108987 | MH793604 | MK108988 | - |
| Monilochaetes infuscans | CBS 869.96 | Ipomoea batatas | South Africa | JQ005780 | JX546612 | JQ005801 | JQ005843 | JQ005864 | - | - | - |
| Species | Culture/Isolate a | Clade | Host | Location | GenBank Accession Number b | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ITS | GAPDH | CHS-1 | HIS3 | ACT | TUB2 | |||||
| C. abscissum | COAD 1877 | Acutatum | Citrus sinensis cv. Pera | Brazil | KP843126 | KP843129 | KP843132 | KP843138 | KP843141 | KP843135 |
| C. acerbum | CBS 128530 | Acutatum | Malus domestica | New Zealand | JQ948459 | JQ948790 | JQ949120 | JQ949450 | JQ949780 | JQ950110 |
| C. acutatum | CBS 112996 | Acutatum | Carica papaya | Australia | JQ005776 | JQ948677 | JQ005797 | JQ005818 | JQ005839 | JQ005860 |
| C. australe | CBS 116478 | Acutatum | Trachycarpus fortunei | South Africa | JQ948455 | JQ948786 | JQ949116 | JQ949446 | JQ949776 | JQ950106 |
| C. brevisporum | BCC 38876 | Magnum | Neoregalia sp. | Thailand | JN050238 | JN050227 | MZ799287 | MZ673841 | JN050216 | JN050244 |
| NFUCB-2 c | Magnum | Cyclocarya paliurus | Hunan, China | OR064061 | OR069517 | OR073868 | OR096507 | OR096434 | OR096482 | |
| NFUCB-6 c | Magnum | Cy. Paliurus | Hunan, China | OR064062 | OR069518 | OR073869 | OR096508 | OR096435 | OR096483 | |
| NFUCB-9 | Magnum | Cy. Paliurus | Hunan, China | OR064063 | OR069519 | OR073870 | OR096509 | OR096436 | OR096484 | |
| NFUCB-11 | Magnum | Cy. Paliurus | Hunan, China | OR064064 | OR069520 | OR073871 | OR096510 | OR096437 | OR096485 | |
| NFUCB-12 c | Magnum | Cy. Paliurus | Guizhou, China | OR064065 | OR069521 | OR073872 | OR096511 | OR096438 | OR096486 | |
| C. brisbanense | CBS 292.67 | Acutatum | Capsicum annuum | Australia | JQ948291 | JQ948621 | JQ948952 | JQ949282 | JQ949612 | JQ949942 |
| C. cacao | CBS 119297 | Magnum | Theobroma cacao | Costa Rica | MG600772 | MG600832 | MG600878 | MG600916 | MG600976 | MG601039 |
| C. cairnsense | BRIP 63642 | Acutatum | Capsicum annuum | Australia | KU923672 | KU923704 | KU923710 | KU923722 | KU923716 | KU923688 |
| C. cattleyicola | CBS 170.49 | Orchidearum | Cattleya sp. | Belgium | MG600758 | MG600819 | MG600866 | MG600905 | MG600963 | MG601025 |
| C. chrysanthemi | IMI 364540 | Acutatum | Chrysanthemum coronarium | China | JQ948273 | JQ948603 | JQ948934 | JQ949264 | JQ949594 | JQ949924 |
| C. cliviicola | CBS 125375 | Orchidearum | Clivia miniata | China | MG600733 | MG600795 | MG600850 | MG600892 | MG600939 | MG601000 |
| C. cosmi | CBS 853.73 | Acutatum | Cosmos sp. | Netherlands | JQ948274 | JQ948604 | JQ948935 | JQ949265 | JQ949595 | JQ949925 |
| C. costaricense | CBS 330.75 | Acutatum | Coffea arabica, cv. Typica | Costa Rica | JQ948180 | JQ948510 | JQ948841 | JQ949171 | JQ949501 | JQ949831 |
| C. cuscutae | IMI 304802 | Acutatum | Cuscuta sp. | Dominica | JQ948195 | JQ948525 | JQ948856 | JQ949186 | JQ949516 | JQ949846 |
| C. eriobotryae | GLMC 1935 | Acutatum | Eriobotrya japonica | China | MF772487 | MF795423 | MN191653 | MN191658 | MN191648 | MF795428 |
| C. fioriniae | CBS 128517 | Acutatum | Fiorinia externa | USA | JQ948292 | JQ948622 | JQ948953 | JQ949283 | JQ949613 | JQ949943 |
| C. godetiae | CBS 133.44 | Acutatum | Clarkia hybrida cv. Kelvon Glory | Denmark | JQ948402 | JQ948733 | JQ949063 | JQ949393 | JQ949723 | JQ950053 |
| NFUCo-1 c | Acutatum | Cy. paliurus | Guizhou, China | OR064051 | OR069507 | OR073858 | OR096497 | OR096424 | OR096472 | |
| NFUCo-4 c | Acutatum | Cy. paliurus | Jiangxi, China | OR064052 | OR069508 | OR073859 | OR096498 | OR096425 | OR096473 | |
| NFUCo-5 c | Acutatum | Cy. paliurus | Hunan, China | OR064053 | OR069509 | OR073860 | OR096499 | OR096426 | OR096474 | |
| NFUCo-8 | Acutatum | Cy. paliurus | Hunan, China | OR064054 | OR069510 | OR073861 | OR096500 | OR096427 | OR096475 | |
| NFUCo-11 | Acutatum | Cy. paliurus | Hunan, China | OR064055 | OR069511 | OR073862 | OR096501 | OR096428 | OR096476 | |
| C. guajavae | IMI 350839 | Acutatum | Psidium guajava | India | JQ948270 | JQ948600 | JQ948931 | JQ949261 | JQ949591 | JQ949921 |
| C. indonesiense | CBS 127551 | Acutatum | Eucalyptus sp. | Indonesia | JQ948288 | JQ948618 | JQ948949 | JQ949279 | JQ949609 | JQ949939 |
| C. javanense | CBS 144963 | Acutatum | Capsicum annuum | Indonesia | MH846576 | MH846572 | MH846573 | MH846571 | MH846575 | MH846574 |
| C. johnstonii | CBS 128532 | Acutatum | Solanum lycopersicum | New Zealand | JQ948444 | JQ948775 | JQ949105 | JQ949435 | JQ949765 | JQ950095 |
| C. kinghornii | CBS 198.35 | Acutatum | Phormium sp. | UK | JQ948454 | JQ948785 | JQ949115 | JQ949445 | JQ949775 | JQ950105 |
| C. laticiphilum | CBS 112989 | Acutatum | Hevea brasiliensis | India | JQ948289 | JQ948619 | JQ948950 | JQ949280 | JQ949610 | JQ949940 |
| C. limetticola | CBS 114.14 | Acutatum | Citrus aurantifolia | USA, Florida | JQ948193 | JQ948523 | JQ948854 | JQ949184 | JQ949514 | JQ949844 |
| C. lobatum | IMI 79736 | Magnum | Piper catalpaefolium | Trinidad | MG600768 | MG600828 | MG600874 | MG600912 | MG600972 | MG601035 |
| C. lupini | CBS 109225 | Acutatum | Lupinus albus | Ukraine | JQ948155 | JQ948485 | JQ948816 | JQ949146 | JQ949476 | JQ949806 |
| C. magnum | CBS 519.97 | Magnum | Citrullus lanatus | USA | MG600769 | MG600829 | MG600875 | MG600913 | MG600973 | MG601036 |
| C. melonis | CBS 159.84 | Acutatum | Cucumis melo | Brazil | JQ948194 | JQ948524 | JQ948855 | JQ949185 | JQ949515 | JQ949845 |
| C. merremiae | CBS 124955 | Magnum | Merremia umbellata | Panama | MG600765 | MG600825 | MG600872 | MG600910 | MG600969 | MG601032 |
| C. monsterae | LC13871 | Orchidearum | Monstera deliciosa | China | MZ595897 | MZ664121 | MZ799351 | MZ673917 | MZ664195 | MZ674015 |
| C. musicola | CBS 132885 | Orchidearum | Musa sp. | Mexico | MG600736 | MG600798 | MG600853 | MG600895 | MG600942 | MG601003 |
| C. nymphaeae | CBS 515.78 | Acutatum | Nymphaea alba | Netherlands | JQ948197 | JQ948527 | JQ948858 | JQ949188 | JQ949518 | JQ949848 |
| NFUCN-2 c | Acutatum | Cy. paliurus | Guangxi, China | OR064071 | OR069527 | OR073878 | OR096517 | OR096444 | OR096492 | |
| NFUCN-5 c | Acutatum | Cy. paliurus | Hunan, China | OR064072 | OR069528 | OR073879 | OR096518 | OR096445 | OR096493 | |
| NFUCN-7 | Acutatum | Cy. paliurus | Hunan, China | OR064073 | OR069529 | OR073880 | OR096519 | OR096446 | OR096494 | |
| NFUCN-11 c | Acutatum | Cy. paliurus | Guizhou, China | OR064074 | OR069530 | OR073881 | OR096520 | OR096447 | OR096495 | |
| NFUCN-14 | Acutatum | Cy. paliurus | Jiangxi, China | OR064075 | OR069531 | OR073882 | OR096521 | OR096448 | OR096496 | |
| C. orchidearum | CBS 135131 | Orchidearum | Dendrobium nobile | Netherlands | MG600738 | MG600800 | MG600855 | MG600897 | MG600944 | MG601005 |
| C. panamense | CBS 125386 | Magnum | Merremia umbellata | Panama | MG600766 | MG600826 | MG600873 | MG600911 | MG600970 | MG601033 |
| C. paxtonii | IMI 165753 | Acutatum | Musa sp. | Saint Lucia | JQ948285 | JQ948615 | JQ948946 | JQ949276 | JQ949606 | JQ949936 |
| C. phormii | CBS 118194 | Acutatum | Phormium sp. | Germany | JQ948446 | JQ948777 | JQ949107 | JQ949437 | JQ949767 | JQ950097 |
| C. piperis | IMI 71397 | Orchidearum | Piper nigrum | Malaysia | MG600760 | MG600820 | MG600867 | MG600906 | MG600964 | MG601027 |
| C. plurivorum | CBS 125474 | Orchidearum | Coffea sp. | Vietnam | MG600718 | MG600781 | MG600841 | MG600887 | MG600925 | MG600985 |
| NFUCP-3 c | Orchidearum | Cy. paliurus | Guizhou, China | OR064066 | OR069522 | OR073873 | OR096512 | OR096439 | OR096487 | |
| NFUCP-5 c | Orchidearum | Cy. paliurus | Hunan, China | OR064067 | OR069523 | OR073874 | OR096513 | OR096440 | OR096488 | |
| NFUCP-7 | Orchidearum | Cy. paliurus | Hunan, China | OR064068 | OR069524 | OR073875 | OR096514 | OR096441 | OR096489 | |
| NFUCP-8 | Orchidearum | Cy. paliurus | Hunan, China | OR064069 | OR069525 | OR073876 | OR096515 | OR096442 | OR096490 | |
| NFUCP-13 c | Orchidearum | Cy. paliurus | Jiangxi, China | OR064070 | OR069526 | OR073877 | OR096516 | OR096443 | OR096491 | |
| C. pyricola | CBS 128531 | Acutatum | Pyrus communis | New Zealand | JQ948445 | JQ948776 | JQ949106 | JQ949436 | JQ949766 | JQ950096 |
| C. reniforme | LC8230 | Orchidearum | Smilax cocculoides | China | MZ595847 | MZ664110 | MZ799290 | MZ673867 | MZ664145 | MZ673968 |
| C. rhombiforme | CBS 129953 | Acutatum | Olea europaea | Portugal | JQ948457 | JQ948788 | JQ949118 | JQ949448 | JQ949778 | JQ950108 |
| C. salicis | CBS 607.94 | Acutatum | Salix sp. | Netherlands | JQ948460 | JQ948791 | JQ949121 | JQ949451 | JQ949781 | JQ950111 |
| C. schimae | LC13880 | Acutatum | Schima sp. | China | MZ595885 | MZ664105 | MZ799347 | MZ673905 | MZ664183 | MZ674003 |
| C. scovillei | CBS 126529 | Acutatum | Capsicum sp. | Indonesia | JQ948267 | JQ948597 | JQ948928 | JQ949258 | JQ949588 | JQ949918 |
| C. simmondsii | CBS 122122 | Acutatum | Carica papaya | Australia | JQ948276 | JQ948606 | JQ948937 | JQ949267 | JQ949597 | JQ949927 |
| C. sloanei | IMI 364297 | Acutatum | Theobroma cacao | Malaysia | JQ948287 | JQ948617 | JQ948948 | JQ949278 | JQ949608 | JQ949938 |
| C. sojae | ATCC 62257 | Orchidearum | Glycine max | USA | MG600749 | MG600810 | MG600860 | MG600899 | MG600954 | MG601016 |
| NFUCS-1 c | Orchidearum | Cy. paliurus | Jiangxi, China | OR064056 | OR069512 | OR073863 | OR096502 | OR096429 | OR096477 | |
| NFUCS-3 c | Orchidearum | Cy. paliurus | Hunan, China | OR064057 | OR069513 | OR073864 | OR096503 | OR096430 | OR096478 | |
| NFUCS-6 | Orchidearum | Cy. paliurus | Hunan, China | OR064058 | OR069514 | OR073865 | OR096504 | OR096431 | OR096479 | |
| NFUCS-10 c | Orchidearum | Cy. paliurus | Guizhou, China | OR064059 | OR069515 | OR073866 | OR096505 | OR096432 | OR096480 | |
| NFUCS-15 | Orchidearum | Cy. paliurus | Fujian, China | OR064060 | OR069516 | OR073867 | OR096506 | OR096433 | OR096481 | |
| C. tamarilloi | CBS 129814 | Acutatum | Solanum betaceum | Colombia | JQ948184 | JQ948514 | JQ948845 | JQ949175 | JQ949505 | JQ949835 |
| C. vittalense | CBS 181.82 | Orchidearum | Theobroma cacao | India | MG600734 | MG600796 | MG600851 | MG600893 | MG600940 | MG601001 |
| C. walleri | CBS 125472 | Acutatum | Coffea sp. | Vietnam | JQ948275 | JQ948605 | JQ948936 | JQ949266 | JQ949596 | JQ949926 |
| Monilochaetes infuscans | CBS 869.96 | outgroup | Ipomoea batatas | South Africa | JQ005780 | JX546612 | JQ005801 | JQ005822 | JQ005843 | JQ005864 |
| Province | County/Location | Leaf Samples | Latitude (N) | Longitude (E) |
|---|---|---|---|---|
| Fujian | Xiapu | 4 | 27°03′08″ | 119°56′33″ |
| Jianyang | 6 | 27°33′08″ | 117°47′03″ | |
| Guangxi | Longsheng | 7 | 26°01′13″ | 109°55′08″ |
| Guizhou | Lipin | 12 | 26°06′50″ | 109°11′08″ |
| Hubei | Yidu | 6 | 30°26′04″ | 111°19′54″ |
| Sui | 6 | 32°11′43″ | 113°16′11″ | |
| Hunan | Jianghua Yao nationality | 13 | 24°54′01″ | 112°06′43″ |
| Jiangxi | Jinggangshan | 6 | 26°42′03″ | 114°17′47″ |
| Shangrao | 7 | 28°49′54″ | 118°11′07″ | |
| Sichuan | Xuyong | 5 | 28°09′11″ | 105°23′54″ |
| Zhejiang | Lanxi | 11 | 29°08′50″ | 119°23′28″ |
References
- Zhao, W.; Tang, D.; Yuan, E.; Wang, M.; Zhang, Q.; Liu, Y.; Shen, B.; Chen, J.; Yin, Z. Inducement and cultivation of novel red Cyclocarya paliurus callus and its unique morphological and metabolic characteristics. Ind. Crops Prod. 2020, 147, 30–33. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, J.; Li, J.; Xiong, L.; Chen, H.; Liu, X.; Wang, N.; Ouyang, K.; Wang, W. Antihyperlipidemic and hepatoprotective activities of polysaccharide fraction from Cyclocarya paliurus in high-fat emulsion-induced hyperlipidaemic mice. Carbohydr. Polym. 2018, 183, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, J.; Nie, Q.; Chang, X.; Fang, Q.; Xie, J.; Li, H.; Nie, S.P. Hypoglycemic mechanism of polysaccharide from Cyclocarya paliurus leaves in type 2 diabetic rats by gut microbiota and host metabolism alteration. Sci. China Life Sci. 2020, 64, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Peng, Y.; Zhu, X.; Li, H.; Zhang, L.; Kong, F.; Wang, J.; Yu, D. The phytochemicals and health benefits of Cyclocarya paliurus (Batalin) Iljinskaja. Front. Nutr. 2023, 10, 1158158. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Hu, W.B.; Yang, Z.W.; Chen, H.; Wang, N.; Liu, X.; Wang, W. Enzymolysis-ultrasonic assisted extraction of flavanoid from Cyclocarya paliurus (Batal) Iljinskaja: HPLC profile, antimicrobial and antioxidant activity. Ind. Crops Prod. 2019, 130, 615–626. [Google Scholar]
- Tu, W.C.; Luo, H.R.; Yuan, E.; Sakah, J.; Yang, Q.Y.; Xiao, W.L.; Zheng, Y.T.; Liu, M.F. Triterpene constituents from the fruits of Cyclocarya paliurus and their anti-HIV-1 IIIB activity. Nat. Prod. Res. 2022, 37, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Quek, S.Y.; Shang, X.; Fang, S. Geographical variations of triterpenoid contents in Cyclocarya paliurus leaves and their inhibitory effects on HeLa cells. Ind. Crops Prod. 2021, 162, 113314. [Google Scholar] [CrossRef]
- Li, X.; Fu, X.; Shang, X.; Yang, W.; Fang, S. Natural population structure and genetic differentiation for heterodicogamous plant: Cyclocarya paliurus (Batal.) Iljinskaja (Juglandaceae). Tree Genet. Genomes 2017, 13, 80. [Google Scholar] [CrossRef]
- Zheng, X.R.; Liu, C.L.; Zhang, M.J.; Shang, X.L.; Fang, S.Z.; Chen, F.M. First report of leaf blight of Cyclocarya paliurus caused by Nigrospora sphaerica in China. Crop Prot. 2020, 140, 105453. [Google Scholar] [CrossRef]
- Zheng, X.R.; Zhang, M.J.; Shang, X.L.; Fang, S.Z.; Chen, F.M. Etiology of Cyclocarya paliurus Anthracnose in Jiangsu Province, China. Front. Plant Sci. 2021, 11, 613499. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Xu, X.; Cheng, J.; Zheng, L.; Huang, J.; Li, D.W. Identification and Characterization of Colletotrichum Species Associated with Anthracnose Disease of Camellia oleifera in China. Plant Dis. 2020, 104, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.; Cai, L.; Cannon, P.; Crouch, J.A.; Crous, P.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.; Jones, E.; et al. Colletotrichum—Names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.; Diao, Y.; Wu, W.; Damm, U.; Song, S.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–56. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Weir, B.; Damm, U.; Crous, P.; Wang, Y.; Liu, B.; Wang, M.; Zhang, M.; Cai, L. Unravelling Colletotrichum species associated with Camellia: Employing ApMat and GS loci to resolve species in the C. gloeosporioides complex. Persoonia 2016, 35, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.; Taylor, P.; Weir, B.; Waller, J.; Abang, M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Gardes, M.; Bruns, T. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Guerber, J.C.; Liu, B.; Correll, J.C.; Johnston, P.R. Characterization of diversity in Colletotrichum acutatum sensu lato by sequence analysis of two gene introns, mtDNA and intron RFLPs, and mating compatibility. Mycologia 2003, 95, 872–895. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risede, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A Multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- Silva, D.N.; Talhinhas, P.; Varzea, V.; Cai, L.; Paulo, O.S.; Batista, D. Application of the Apn2/MAT locus to improve the systematics of the Colletotrichum gloeosporioides complex: An example from coffee (Coffea spp.) hosts. Mycologia 2012, 104, 396–409. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Zheng, X.R.; Zhang, M.J.; Qiao, Y.H.; Li, R.; Alkan, N.; Chen, J.Y.; Chen, F.M. Cyclocarya paliurus Reprograms the Flavonoid Biosynthesis Pathway Against Colletotrichum fructicola. Front. Plant Sci. 2022, 13, 933484. [Google Scholar] [CrossRef] [PubMed]
- He, L.F.; Li, X.X.; Gao, Y.Y.; Li, B.X.; Mu, W.; Liu, F. Characterization and Fungicide Sensitivity of Colletotrichum spp. from Different Hosts in Shandong, China. Plant Dis. 2019, 103, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum Species Responsible for Anthracnose Diseases of Various Fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.; Horowitz, S.; Sharon, A. Pathogenic and Nonpathogenic Lifestyles in Colletotrichum acutatum from Strawberry and other Plants. Phytopathology 2001, 91, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.; Seifert, K.; Huhndorf, S.M.; Robert, V.; Spouge, J.; Levesque, C.; Chen, W.; Janzen, D.; Consortium, A. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Nilsson, R.H.; Alias, S.A.; Ariyawansa, H.A.; Blair, J.E.; Cai, L.; Cock, A.W.A.M.; Dissanayake, A.J.; Glockling, S.L.; Goonasekara, I.D.; et al. One stop shop: Backbones trees for important phytopathogenic genera: I. Fungal Divers. 2014, 67, 21–125. [Google Scholar] [CrossRef]
- Tovar-Pedraza, J.M.; Mora-Aguilera, J.A.; Nava-Díaz, C.; Lima, N.B.; Michereff, S.J.; Sandoval-Islas, J.S.; Câmara, M.P.S.; Téliz-Ortiz, D.; Leyva-Mir, S.G. Distribution and Pathogenicity of Colletotrichum Species Associated with Mango Anthracnose in Mexico. Plant Dis. 2020, 104, 137–146. [Google Scholar] [CrossRef]
- Prihastuti, H.; McKenzie, E.; Hyde, K.; Cai, L.; Hu, M.; Hyde, E. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum Species Causing Anthracnose of Citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef]
- Eaton, M.; Edwards, S.; Inocencio, H.; Machado, F.; Nuckles, E.; Farman, M.; Gauthier, N.; Vaillancourt, L. Diversity and Cross-Infection Potential of Colletotrichum Causing Fruit Rots in Mixed-Fruit Orchards in Kentucky. Plant Dis. 2020, 105, 1115–1128. [Google Scholar] [CrossRef]
- Moreira, R.R.; Peres, N.A.; May, D.M.L.L. Colletotrichum acutatum and C. gloeosporioides Species Complexes Associated with Apple in Brazil. Plant Dis. 2019, 103, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Noor, N.M.; Zakaria, L. Identification and characterization of Colletotrichum spp. associated with chili anthracnose in peninsular Malaysia. Eur. J. Plant Pathol. 2018, 151, 961–973. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Zhou, Y.; Tan, G.; Zhang, L. Identification and characterization of Colletotrichum species associated with Camellia sinensis anthracnose in Anhui province, China. Plant Dis. 2020, 105, 2649–2657. [Google Scholar] [CrossRef]
- Jian, Y.; Li, Y.; Tang, G.; Zheng, X.; Khaskheli, M.I.; Gong, G. Identification of Colletotrichum Species Associated with Anthracnose Disease of Strawberry in Sichuan Province, China. Plant Dis. 2021, 105, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fu, D.; Wang, W.; Gleason, M.L.; Zhang, R.; Liang, X.; Sun, G. Diversity of Colletotrichum Species Causing Apple Bitter Rot and Glomerella Leaf Spot in China. J. Fungi 2022, 8, 740. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Schnabel, G.; Chaisiri, C.; Yin, L.; Yin, W.; Luo, C. Colletotrichum Species Associated with Peaches in China. J. Fungi 2022, 8, 313. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Zhao, G.; Li, Q.; Solangi, G.S.; Tang, L.; Guo, T.; Huang, S.; Hsiang, T. Identification and Characterization of Colletotrichum Species Associated with Mango Anthracnose in Guangxi, China. Plant Dis. 2018, 102, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Jin, G.; Li, D.; Wu, S.; Zhu, L. First report of Colletotrichum fructicola causing leaf spots on Liriodendron chinense × tulipifera in China. Forest Pathol. 2022, 52, e12779. [Google Scholar] [CrossRef]










| Loci | Product Name | Primer | Direction | Sequence (5′-3′) | Reference |
|---|---|---|---|---|---|
| ITS | Internal transcribed spacer | ITS1F | Forward | CTTGGTCATTTAGAGGAAGTAA | Gardes and Bruns [20] |
| ITS4 | Reverse | TCCTCCGCTTATTGATATGC | White et al. [21] | ||
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | GDF1 | Forward | GCCGTCAACGACCCCTTCATTGA | Guerber et al. [22] |
| GDR1 | Reverse | GGGTGGAGTCGTACTTGAGCATGT | Guerber et al. [22] | ||
| CHS-1 | Chitin synthase 1 | CHS-79F | Forward | TGGGGCAAGGATGCTTGGAAGAAG | Carbone and Kohn [23] |
| CHS-354R | Reverse | TGGAAGAACCATCTGTGAGAGTTG | Carbone and Kohn [23] | ||
| HIS3 | histone H3 | CYLH3F | Forward | AGGTCCACTGGTGGCAAG | Crous et al. [24] |
| CYLH3R | Reverse | AGCTGGATGTCCTTGGACTG | Crous et al. [24] | ||
| ACT | Actin | ACT-512F | Forward | ATGTGCAAGGCCGGTTTCGC | Carbone and Kohn [23] |
| ACT-783R | Reverse | TACGAGTCCTTCTGGCCCAT | Carbone and Kohn [23] | ||
| TUB | β-tubulin | T1 | Forward | AACATGCGTGAGATTGTAAGT | O’Donnell and Cigelnik [25] |
| Bt-2b | Reverse | ACCCTCAGTGTAGTGACCCTTGGC | Glass and Donaldson [26] | ||
| CAL | Calmodulin | CL1A | Forward | GATCAAGGAGGCCTTCTC | O’Donnell et al. [27] |
| CL2A | Reverse | TTTTTGCATCATGAGTTGGAC | O’Donnell et al. [27] | ||
| GS | Glutamine synthetase | GSLF2 | Forward | TACACGAGSAAAAGGATACGC | Liu et al. [17] |
| GSLR1 | Reverse | AGRCGCACATTGTCAGTATCG | Liu et al. [17] | ||
| ApMat | Apn2-Mat1-2 intergenic spacer | AM-F | Forward | TCATTCTACGTATGTGCCCG | Silva et al. [28] |
| AM-R | Reverse | CCAGAAATACACCGAACTTGC | Silva et al. [28] |
| Species | Colony Appearance | Growth Rate (mm/d) a | Conidia | Appressoria | Ascospores | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Length (μm) b | Width (μm) b | Shape | Length (μm) b | Width (μm) b | Shape | Length (μm) b | Width (μm) b | Shape | |||
| Colletotrichum brevisporum | Dense, dark-grey with the grey aerial mycelium and edges | 12.7 ± 0.2 B | 14.1 ± 1.2 (10.6–17.3) | 5.8 ± 0.3 (5.0–6.8) | Cylindrical | 10.5 ± 1.7 (7.5–17.5) | 8.9 ± 1.4 (5.6–13.2) | Globose, entire or lobed margin | / | / | / |
| C. fructicola | Dense, olive-grey with the white edge hyphae | 14.4 ± 0.2 A | 13.5 ± 1.8 (10.3–22.5) | 5.8 ± 0.5 (4.4–7.9) | Cylindrical | 9.5 ± 1.6 (6.5–16.0) | 7.0 ± 0.9 (4.5–9.1) | Ovoid to slightly irregular | 17.7 ± 1.7 (12.3–23.2) | 5.0 ± 0.6 (3.8–6.5) | Curved fusoid |
| C. gloeosporioides | Dense, white with whitish aerial mycelium and edges | 12.6 ± 0.5 B | 15.9 ± 1.1 (13.1–22.7) | 5.5 ± 0.4 (4.5–6.3) | Cylindrical | 9.6 ± 1.0 (7.2–12.5) | 7.2 ± 0.9 (6.0–10.3) | Ovoid to slightly irregular | / | / | / |
| C. godetiae | Dense, white hyphae, lack of aerial mycelium | 8.4 ± 0.2 E | 15.9 ± 1.3 (12.6–20.7) | 5.1 ± 0.4 (3.8–6.8) | Fusiform | 9.5 ± 1.0 (7.6–13.2) | 6.4 ± 0.7 (4.9–8.7) | Ovoid to globose | / | / | / |
| C. nymphaeae | Dense, olive-grey with white margin, lack of aerial mycelium | 9.8 ± 0.2 D | 14.5 ± 1.9 (11.1–18.0) | 5.5 ± 0.9 (4.0–6.9) | Fusiform | 9.1 ± 1.3 (7.0–11.9) | 7.0 ± 1.1 (5.0–8.9) | Ovoid, with smooth margin | / | / | / |
| C. plurivorum | Dense, olive-grey with the white edge hyphae | 11.1 ± 0.1 C | 14.9 ± 1.6 (12.1–20.2) | 6.2 ± 0.6 (5.0–7.7) | Cylindrical | 12.4 ± 2.2 (8.6–20.5) | 9.2 ± 1.2 (6.4–12.5) | Globose, entire or lobed margin | 18.0 ± 1.6 (13.6–23.0) | 7.0 ± 0.8 (5.0–9.3) | Fusiform to curved fusoid |
| C. sojae | Dense, light orange-red with the whitish aerial mycelium and edges | 14.7 ± 0.7 A | / | / | / | 11.1 ± 1.7 (7.2–18.0) | 7.4 ± 0.7 (5.7–9.4) | Ovoid, entire or lobed margin | 24.4 ± 4.4 (13.2–32.1) | 5.0 ± 0.7 (3.0–6.8) | Curved fusoid |
| Detached Leaves a | Intact Plant a | |||||
|---|---|---|---|---|---|---|
| Wounding | Nonwounding | Wounding | ||||
| Species | Disease Incidence (%) | Lesion Diameter (cm) | Disease Incidence (%) | Lesion Diameter (cm) | Disease Incidence (%) | Lesion Diameter (cm) |
| Colletotrichum brevisporum | 66.7 ± 10.3 | 5.4 ± 1.1 D | – | – | – | – |
| C. fructicola | 100.0 ± 0.0 | 25.3 ± 0.8 A | 100.0 ± 0.0 | 20.1 ± 1.3 a | 88.7 ± 7.2 | 9.4 ± 1.0 a |
| C. gloeosporioides | 100.0 ± 0.0 | 18.0 ± 1.0 B | 100.0 ± 0.0 | 17.7 ± 0.9 a | 66.7 ± 8.7 | 6.2 ± 1.1 ab |
| C. godetiae | 100.0 ± 0.0 | 21.8 ± 1.0 AB | 100.0 ± 0.0 | 16.9 ± 1.2 a | 66.3 ± 12.2 | 6.1 ± 1.1 ab |
| C. nymphaeae | 100.0 ± 0.0 | 19.2 ± 0.7 B | 55.2 ± 11.1 | 3.6 ± 0.8 b | 33.0 ± 12.0 | 2.3 ± 0.9 b |
| C. plurivorum | 66.2 ± 8.7 | 6.6 ± 1.2 CD | – | – | – | – |
| C. sojae | 100.0 ± 0.0 | 10.4 ± 0.7 C | 38.5 ± 13.2 | 2.3 ± 0.7 b | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.-R.; Zhang, M.-J.; Chen, F.-M. Colletotrichum Species Causing Cyclocarya paliurus Anthracnose in Southern China. Forests 2024, 15, 490. https://doi.org/10.3390/f15030490
Zheng X-R, Zhang M-J, Chen F-M. Colletotrichum Species Causing Cyclocarya paliurus Anthracnose in Southern China. Forests. 2024; 15(3):490. https://doi.org/10.3390/f15030490
Chicago/Turabian StyleZheng, Xiang-Rong, Mao-Jiao Zhang, and Feng-Mao Chen. 2024. "Colletotrichum Species Causing Cyclocarya paliurus Anthracnose in Southern China" Forests 15, no. 3: 490. https://doi.org/10.3390/f15030490