
Carbon Allocation to Leaves and Its Controlling Factors and Impacts on Gross Primary Productivity in Forest Ecosystems of Northeast China

Abstract
:1. Introduction
2. Materials and Methods
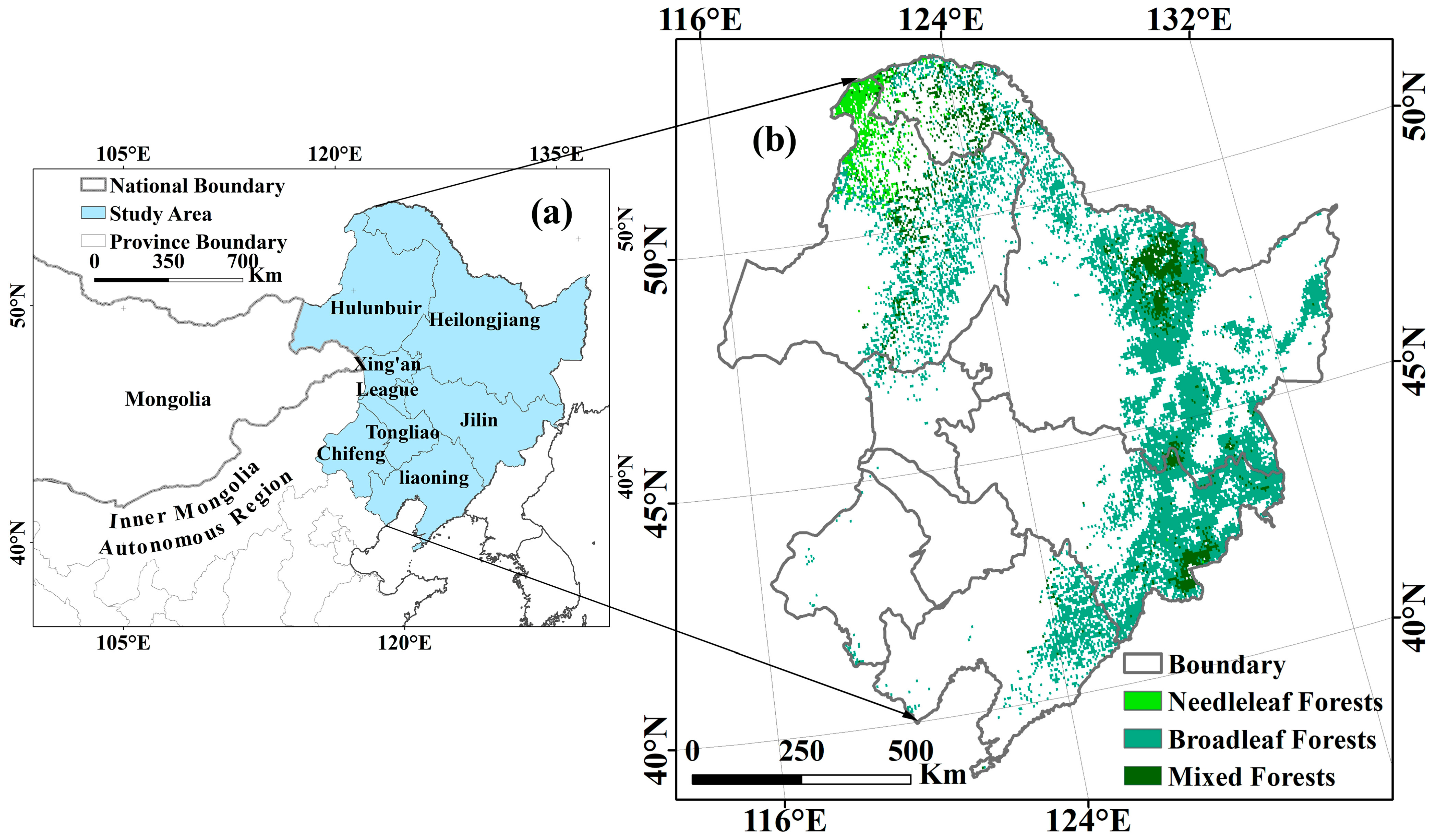
2.1. Overview of the Study Area and Data Sources
2.1.1. Study Area
2.1.2. Remote Sensing Data
2.2. Methods
2.2.1. Estimation of Carbon Allocation to Leaves
2.2.2. Forecast of Future Trends
2.2.3. Statistical Analyses
- (1)
- Pearson correlation analysis
- (2)
- Random forest (RF)
- (3)
- Structural equation model
3. Results
3.1. Spatiotemporal Distribution of Carbon Allocation to Leaves
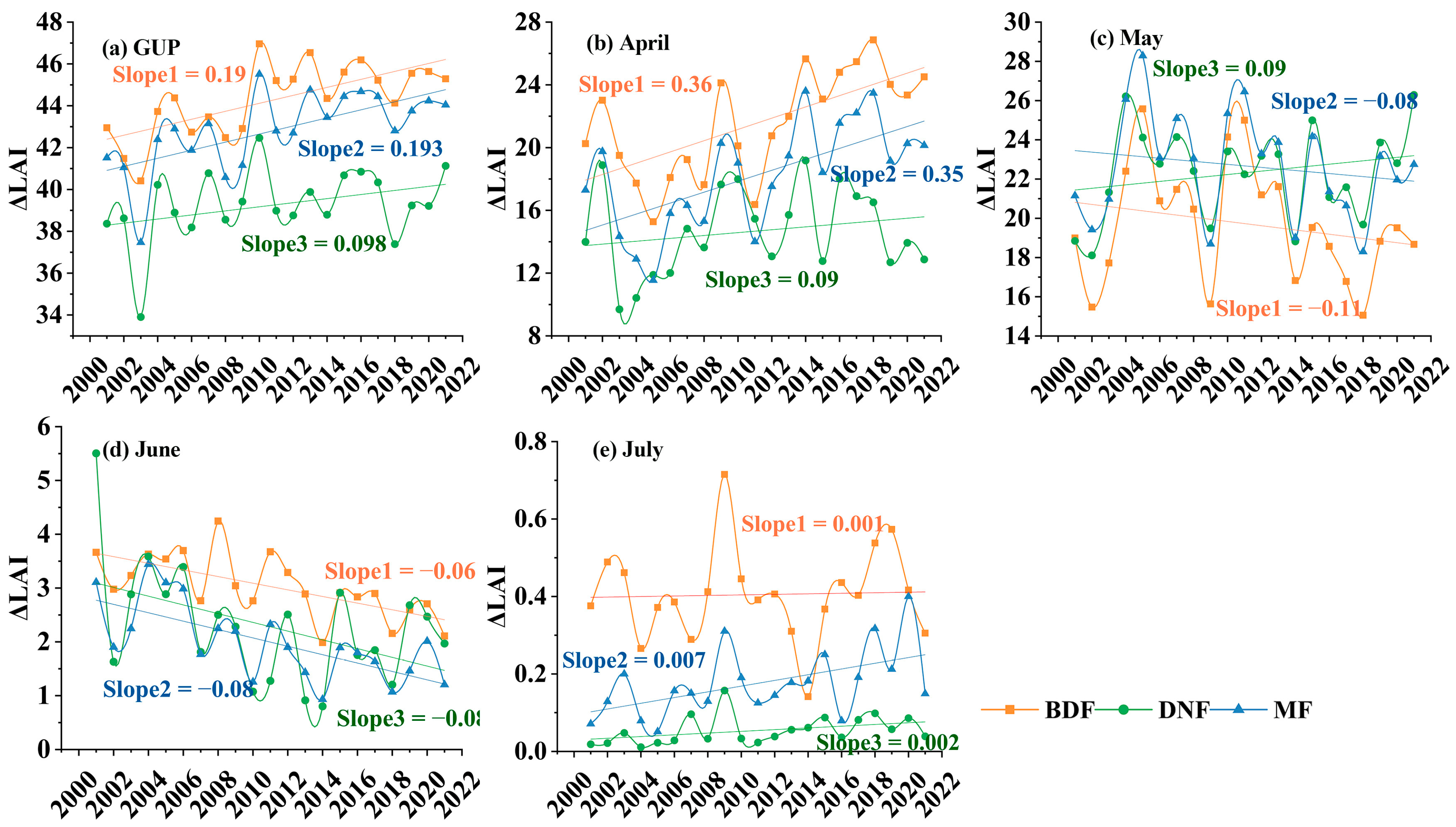
3.1.1. Interannual Variation Trend of Carbon Allocation to Leaves
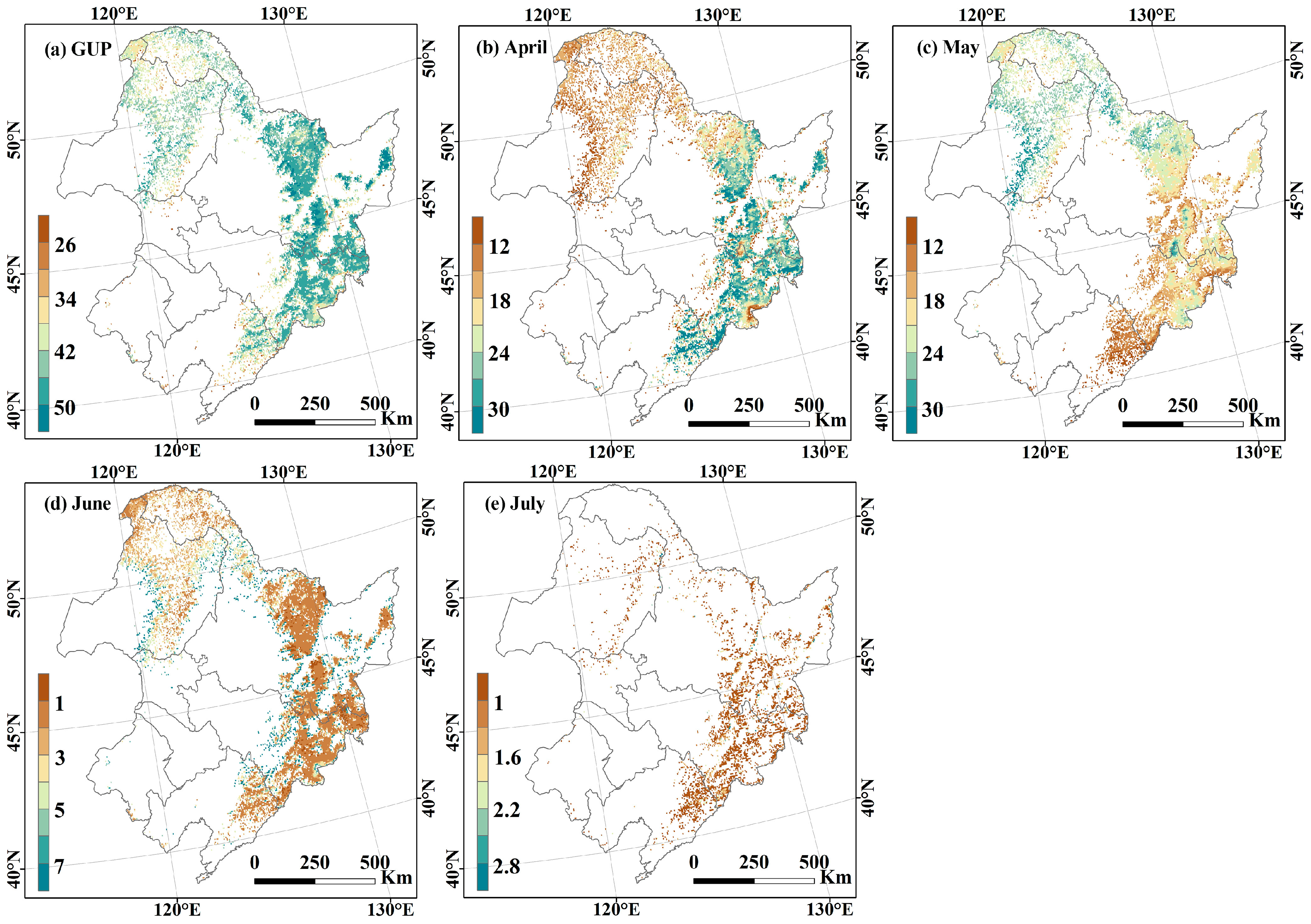
3.1.2. Spatial Distribution of Carbon Allocation to Leaves
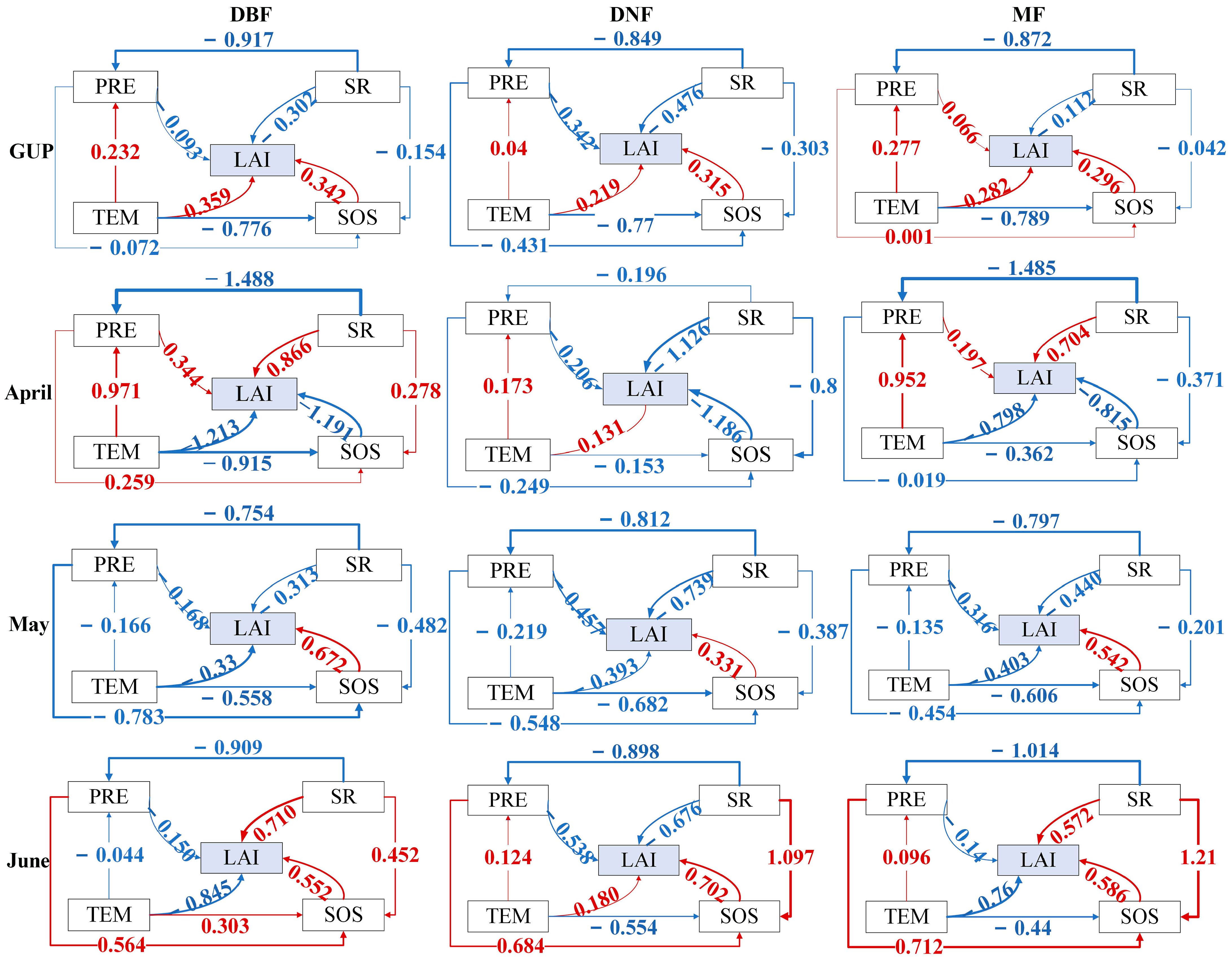
3.2. Analysis of Driving Factors of Carbon Allocation to Leaves
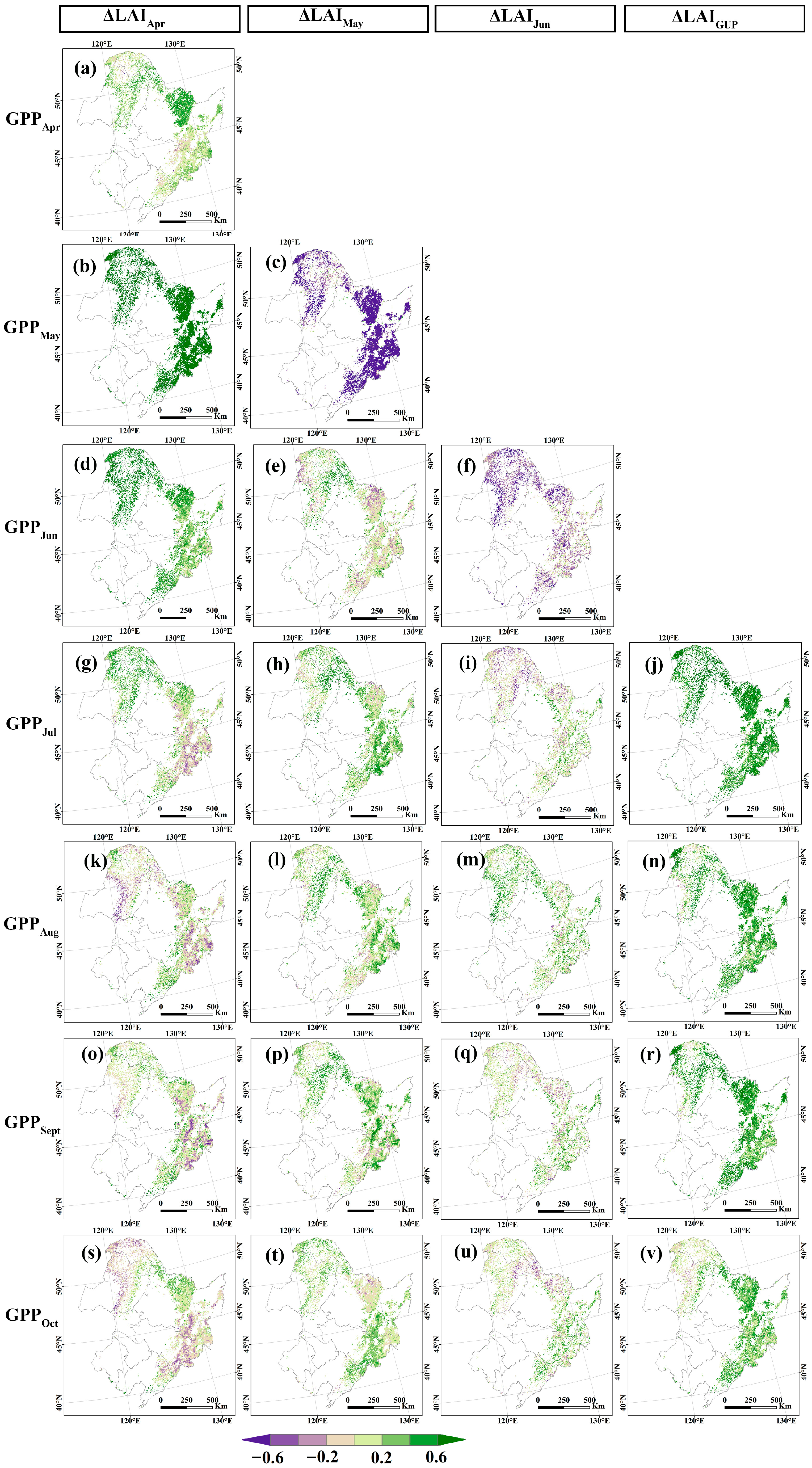
3.3. Effects of Carbon Allocation to Leaves on Gross Primary Productivity
4. Discussion
4.1. Spatiotemporal Distribution of Leaf Carbon Distribution
4.2. Driving Factors of Leaf Carbon Distribution
4.3. Effects of Carbon Allocation to Leaves on Gross Primary Productivity
4.4. Limitations and Prospects
5. Conclusions
- (1)
- Owing to the differences in physiological attributes, in the GUP, the ΔLAI values of DBF and MF are much higher than that of DNF, and all three show an insignificant increasing trend each year. The highest ΔLAI in DBF occurred in April and in DNF and MF it occurred in May. The ΔLAI of DBF and MF showed a significant year-by-year increasing trend in April, and DNF showed a significant increasing trend in most areas in May;
- (2)
- The main factors driving ΔLAI in GUP are TEM and SOS. The main driving factors in April and May were SR and SOS. The driving mechanism in June was the most complex, and the difference between different forestlands was the highest. Except for PRE in DBF and MF, all other factors had larger path coefficients. The coefficients of SR and SOS were the highest for DNF;
- (3)
- ΔLAI in GUP has a significant impact on the GPP. In the MF, the higher ΔLAI in May was most conducive to an increase in GPP. In DBF and DNF, the ΔLAI in April and May both promote the increase of GPP.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brüggemann, N.; Gessler, A.; Kayler, Z.; Keel, S.G.; Badeck, F.; Barthel, M.; Boeckx, P.; Buchmann, N.; Brugnoli, E.; Esperschütz, J.; et al. Carbon allocation and carbon isotope fluxes in the plant-soil-atmosphere continuum: A review. Biogeosciences 2011, 8, 3457–3489. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.; Matson, P.A.; Vitousek, P.M. Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer: New York, NY, USA, 2011; Volume XV, 529p. [Google Scholar]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Hartmann, H.; Bahn, M.; Carbone, M.; Richardson, A.D. Plant carbon allocation in a changing world–challenges and progress: Introduction to a Virtual Issue on carbon allocation. New Phytol. 2020, 227, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Bala, G.; Caldeira, K.; Nemani, R.; Ban-Weiss, G. Importance of carbon dioxide physiological forcing to future climate change. Proc. Natl. Acad. Sci. USA 2010, 107, 9513–9518. [Google Scholar] [CrossRef]
- Doughty, C.E.; Metcalfe, D.; Girardin, C.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.; Araujo-Murakami, A.; Da Costa, M.; Rocha, W. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef]
- Xia, J.; Yuan, W.; Wang, Y.-P.; Zhang, Q. Adaptive Carbon Allocation by Plants Enhances the Terrestrial Carbon Sink. Sci. Rep. 2017, 7, 3341. [Google Scholar] [CrossRef]
- Trugman, A.T.; Detto, M.; Bartlett, M.K.; Medvigy, D.; Anderegg, W.R.L.; Schwalm, C.; Schaffer, B.; Pacala, S.W.; Cameron, D. Tree carbon allocation explains forest drought-kill and recovery patterns. Ecol. Lett. 2018, 21, 1552–1560. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.N.; Walker, A.P.; Dietze, M.C.; Wang, Y.P.; Luo, Y.; Jain, A.K.; El-Masri, B.; Hickler, T. Where does the carbon go? A model–data intercomparison of vegetation carbon allocation and turnover processes at two temperate forest free-air CO2 enrichment sites. New Phytol. 2014, 203, 883–899. [Google Scholar] [CrossRef]
- Negrón-Juárez, R.I.; Koven, C.D.; Riley, W.J.; Knox, R.G.; Chambers, J.Q. Observed allocations of productivity and biomass, and turnover times in tropical forests are not accurately represented in CMIP5 Earth system models. Environ. Res. Lett. 2015, 10, 064017. [Google Scholar] [CrossRef]
- Zhou, Q.; Shi, H.; He, R.; Liu, H.; Zhu, W.; Yu, D.; Zhang, Q.; Dang, H. Prioritized carbon allocation to storage of different functional types of species at the upper range limits is driven by different environmental drivers. Sci. Total Environ. 2021, 773, 145581. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, J.; Francois, C.; Hmimina, G.; Dufrene, E.; Martin-StPaul, N.K.; Soudani, K.; Marie, G.; Ourcival, J.M.; Delpierre, N. Environmental control of carbon allocation matters for modelling forest growth. New Phytol. 2017, 214, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Braswell, B.H.; Schimel, D.S.; Under, E.; Moore, B., III. The response of global terrestrial ecosystems to interannual temperature variability. Science 1997, 278, 870–872. [Google Scholar] [CrossRef]
- Sun, P.; Zhe, Q.U.; Yuan, C.; Yuan, Y.; Wang, C.; Jia, Q. Meteorological Tower Observed CO2 Flux and Footprint in the Forest of Xiaoxing’an Mountains, Northeast China. J. Meteorol. 2023, 37, 15. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J. A Global, 0.05-Degree Product of Solar-Induced Chlorophyll Fluorescence Derived from OCO-2, MODIS, and Reanalysis Data. Remote Sens. 2019, 11, 517. [Google Scholar] [CrossRef]
- Pantin, F.; Simonneau, T.; Muller, B. Coming of leaf age: Control of growth by hydraulics and metabolics during leaf ontogeny. New Phytol. 2012, 196, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Aalen, O.O. A linear regression model for the analysis of life times. Stat. Med. 1989, 8, 907–925. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Huang, Y.; Chen, J.; Benesty, J.; Benesty, J.; Chen, J.; Huang, Y.; Cohen, I. Pearson correlation coefficient. In Noise Reduction in Speech Processing; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–4. [Google Scholar]
- Breiman, L. Random forest. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Flach, P. Machine Learning: The Art and Science of Algorithms that Make Sense of Data; Cambridge University Press: New York, NY, USA, 2012. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 23, 18–22. [Google Scholar]
- Dhaene, S.; Rosseel, Y. An Evaluation of Non-Iterative Estimators in the Structural after Measurement (SAM) Approach to Structural Equation Modeling (SEM). Struct. Equ. Model. A Multidiscip. J. 2023, 30, 926–940. [Google Scholar] [CrossRef]
- Rosseel, Y. lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Lefcheck, J.S. SEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, B.; Henderson, M.; Shen, X.; Su, Y.; Zhou, W. Changing Spring Phenology of Northeast China Forests during Rapid Warming and Short-Term Slowdown Periods. Forests 2022, 13, 2173. [Google Scholar] [CrossRef]
- Donnelly, A.; Yu, R.; Caffarra, A.; Hanes, J.; Liang, L.; Desai, A.R.; Liu, L.; Schwartz, M.D. Interspecific and interannual variation in the duration of spring phenophases in a northern mixed forest. Agric. For. Meteorol. 2017, 243, 55–67. [Google Scholar] [CrossRef]
- Leuschner, C.; Backes, K.; Hertel, D.; Schipka, F.; Schmitt, U.; Terborg, O.; Runge, M. Drought responses at leaf, stem and fine root levels of competitive Fagus sylvatica L. and Quercus petraea (Matt.) Liebl. trees in dry and wet years. For. Ecol. Manag. 2001, 149, 33–46. [Google Scholar] [CrossRef]
- López, B.C.; Sabate, S.; Gracia, C.A. Thinning effects on carbon allocation to fine roots in a Quercus ilex forest. Tree Physiol. 2003, 23, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, J.; Martin-StPaul, N.K.; Dufrêne, E.; François, C.; Soudani, K.; Ourcival, J.M.; Delpierre, N. The dynamic of the annual carbon allocation to wood in European tree species is consistent with a combined source–sink limitation of growth: Implications for modelling. Biogeosciences 2015, 12, 2773–2790. [Google Scholar] [CrossRef]
- Franklin, O.; Johansson, J.; Dewar, R.C.; Dieckmann, U.; McMurtrie, R.E.; Brännström, Å.; Dybzinski, R. Modeling carbon allocation in trees: A search for principles. Tree Physiol. 2012, 32, 648–666. [Google Scholar] [CrossRef]
- Chen, G.; Yang, Y.; Robinson, D. Allocation of gross primary production in forest ecosystems: Allometric constraints and environmental responses. New Phytol. 2013, 200, 1176–1186. [Google Scholar] [CrossRef]
- Meng, F.; Liu, D.; Wang, Y.; Wang, S.; Wang, T. Negative relationship between photosynthesis and late-stage canopy development and senescence over Tibetan Plateau. Glob. Change Biol. 2023, 29, 3147–3158. [Google Scholar] [CrossRef]
- Trugman, A.T.; Anderegg, L.D.L.; Wolfe, B.T.; Birami, B.; Ruehr, N.K.; Detto, M.; Bartlett, M.K.; Anderegg, W.R.L. Climate and plant trait strategies determine tree carbon allocation to leaves and mediate future forest productivity. Glob. Change Biol. 2019, 25, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Hong, S.; Wang, J.; Chen, A.; Zhang, Y.; Zhang, Y.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Penuelas, J.; et al. Climate change increases carbon allocation to leaves in early leaf green-up. Ecol. Lett. 2023, 26, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ran, J.; Hu, W.; Dong, L.; Ji, M.; Jia, X.; Lu, J.; Gong, H.; Aqeel, M.; Yao, S. Effects of biotic and abiotic factors on forest biomass fractions. Natl. Sci. Rev. 2021, 8, nwab025. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Nianpeng, H.E.; Hou, J. Spatial patterns and influencing factors of specific leaf area in typical temperate forests. Acta Ecol. Sin. 2022, 42, 872–883. [Google Scholar]
- Hoffmann, W.A.; Franco, A.C.; Moreira, M.Z.; Haridasan, M. Specific leaf area explains differences in leaf traits between congeneric savanna and forest trees. Funct. Ecol. 2005, 19, 932–940. [Google Scholar] [CrossRef]
- Nabeshima, E.; Kubo, T.; Hiura, T. Variation in tree diameter growth in response to the weather conditions and tree size in deciduous broad-leaved trees. For. Ecol. Manag. 2010, 259, 1055–1066. [Google Scholar] [CrossRef]
- Zhou, X.; Geng, X.; Yin, G.; Hänninen, H.; Fu, Y.H. Legacy effect of spring phenology on vegetation growth in temperate China. Agric. For. Meteorol. 2019, 281, 107845. [Google Scholar] [CrossRef]
- Huang, Z.; Zhou, L.; Chi, Y. Spring phenology rather than climate dominates the trends in peak of growing season in the Northern Hemisphere. Glob. Change Biol. 2023, 29, 4543–4555. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Chen, A.; Wang, K.; Li, X.; Buermann, W.; Huntingford, C.; Peñuelas, J.; Xu, H.; Myneni, R.B. Seasonal biological carryover dominates northern vegetation growth. Nat. Commun. 2021, 12, 983. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Li, L.Z.; Li, Y.; Huntingford, C.; Ciais, P.; Cescatti, A.; Janssens, I.A.; Peñuelas, J.; Buermann, W. Summer soil drying exacerbated by earlier spring greening of northern vegetation. Sci. Adv. 2020, 6, eaax0255. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef] [PubMed]
- Bo, Y.; Li, X.; Liu, K.; Wang, S.; Zhang, H.; Gao, X.; Zhang, X. Three Decades of Gross Primary Production (GPP) in China: Variations, Trends, Attributions, and Prediction Inferred from Multiple Datasets and Time Series Modeling. Remote Sens. 2022, 14, 2564. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, L.; Tian, X.; Zhang, J.; Yuan, W.; Zheng, Y.; Zhao, X.; Kato, T. Assimilation of remotely-sensed leaf area index into a dynamic vegetation model for gross primary productivity estimation. Remote Sens. 2017, 9, 188. [Google Scholar] [CrossRef]
- Chen, X.; Le, X.; Niklas, K.J.; Hu, D.; Zhong, Q.; Cheng, D. Divergent leaf nutrient-use strategies of coexistent evergreen and deciduous trees in a subtropical forest. J. Plant Ecol. 2023, 16, rtac093. [Google Scholar] [CrossRef]
- Zhao, W.; Tan, W.; Li, S. High leaf area index inhibits net primary production in global temperate forest ecosystems. Environ. Sci. Pollut. Res. 2021, 28, 22602–22611. [Google Scholar] [CrossRef]
- Chen, X.; Cai, A.; Guo, R.; Liang, C.; Li, Y. Variation of gross primary productivity dominated by leaf area index in significantly greening area. J. Geogr. Sci. 2023, 33, 1747–1764. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, G.; Chen, Z.; Zhu, X.; Han, L.; Liu, Z.; Lin, Y.; Han, S.; Sha, L.; Wang, H. Photosynthetic capacity dominates the interannual variation of annual gross primary productivity in the Northern Hemisphere. Sci. Total Environ. 2022, 849, 157856. [Google Scholar] [CrossRef]
- Chen, S.; Chen, Z.; Kong, Z.; Zhang, Z. The increase of leaf water potential and whole-tree hydraulic conductance promotes canopy conductance and transpiration of Pinus tabulaeformis during soil droughts. Trees 2023, 37, 41–52. [Google Scholar] [CrossRef]
- Liao, Z.; Zeng, H.; Fan, J.; Lai, Z.; Zhang, C.; Zhang, F.; Wang, H.; Cheng, M.; Guo, J.; Li, Z. Effects of plant density, nitrogen rate and supplemental irrigation on photosynthesis, root growth, seed yield and water-nitrogen use efficiency of soybean under ridge-furrow plastic mulching. Agric. Water Manag. 2022, 268, 107688. [Google Scholar] [CrossRef]
- Zhou, B.; Chen, Y.; Zeng, L.; Cui, Y.; Li, J.; Tang, H.; Liu, J.; Tang, J. Soil nutrient deficiency decreases the postharvest quality-related metabolite contents of tea (Camellia sinensis (L.) Kuntze) leaves. Food Chem. 2022, 377, 132003. [Google Scholar] [CrossRef]
- D’Andrea, E.; Scartazza, A.; Battistelli, A.; Collalti, A.; Proietti, S.; Rezaie, N.; Matteucci, G.; Moscatello, S. Unravelling resilience mechanisms in forests: Role of non-structural carbohydrates in responding to extreme weather events. Tree Physiol. 2021, 41, 1808–1818. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Veen, G.; Bonis, A.; Bradford, E.M.; Classen, A.T.; Cornelissen, J.H.C.; Crowther, T.W.; De Long, J.R.; Freschet, G.T.; Kardol, P. A test of the hierarchical model of litter decomposition. Nat. Ecol. Evol. 2017, 1, 1836–1845. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hurst | Slope | Future Trends |
|---|---|---|
| >0.5 | >0 | Continuous increase |
| <0 | Continuous decrease | |
| <0.5 | >0 | From increase to decrease |
| <0 | From decrease to increase | |
| =0.5 | Unpredictable |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Lai, Q.; Bao, Y.; Sude, B.; Bao, Z.; Liu, X. Carbon Allocation to Leaves and Its Controlling Factors and Impacts on Gross Primary Productivity in Forest Ecosystems of Northeast China. Forests 2024, 15, 129. https://doi.org/10.3390/f15010129
Li Z, Lai Q, Bao Y, Sude B, Bao Z, Liu X. Carbon Allocation to Leaves and Its Controlling Factors and Impacts on Gross Primary Productivity in Forest Ecosystems of Northeast China. Forests. 2024; 15(1):129. https://doi.org/10.3390/f15010129
Chicago/Turabian StyleLi, Zhiru, Quan Lai, Yuhai Bao, Bilige Sude, Zhengyi Bao, and Xinyi Liu. 2024. "Carbon Allocation to Leaves and Its Controlling Factors and Impacts on Gross Primary Productivity in Forest Ecosystems of Northeast China" Forests 15, no. 1: 129. https://doi.org/10.3390/f15010129