

Greenhouse Gas Fluxes from the Epiphytic Lichens: Incubation Experiments
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Site
2.2. Study Objects
Species Characteristics
2.3. EL Sampling and Probe Preparation
2.4. Methods for the Assessment of EL Diversity
2.5. EL Incubations
2.6. Data Examination
3. Results
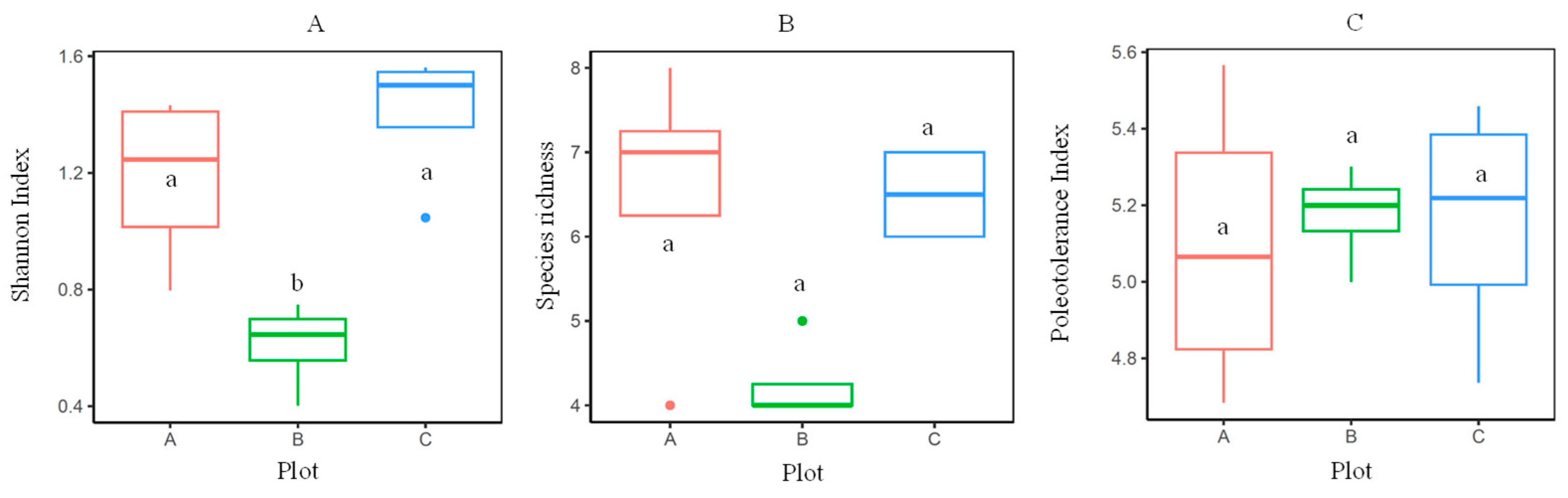
3.1. Assessment of the EL Biodiversity
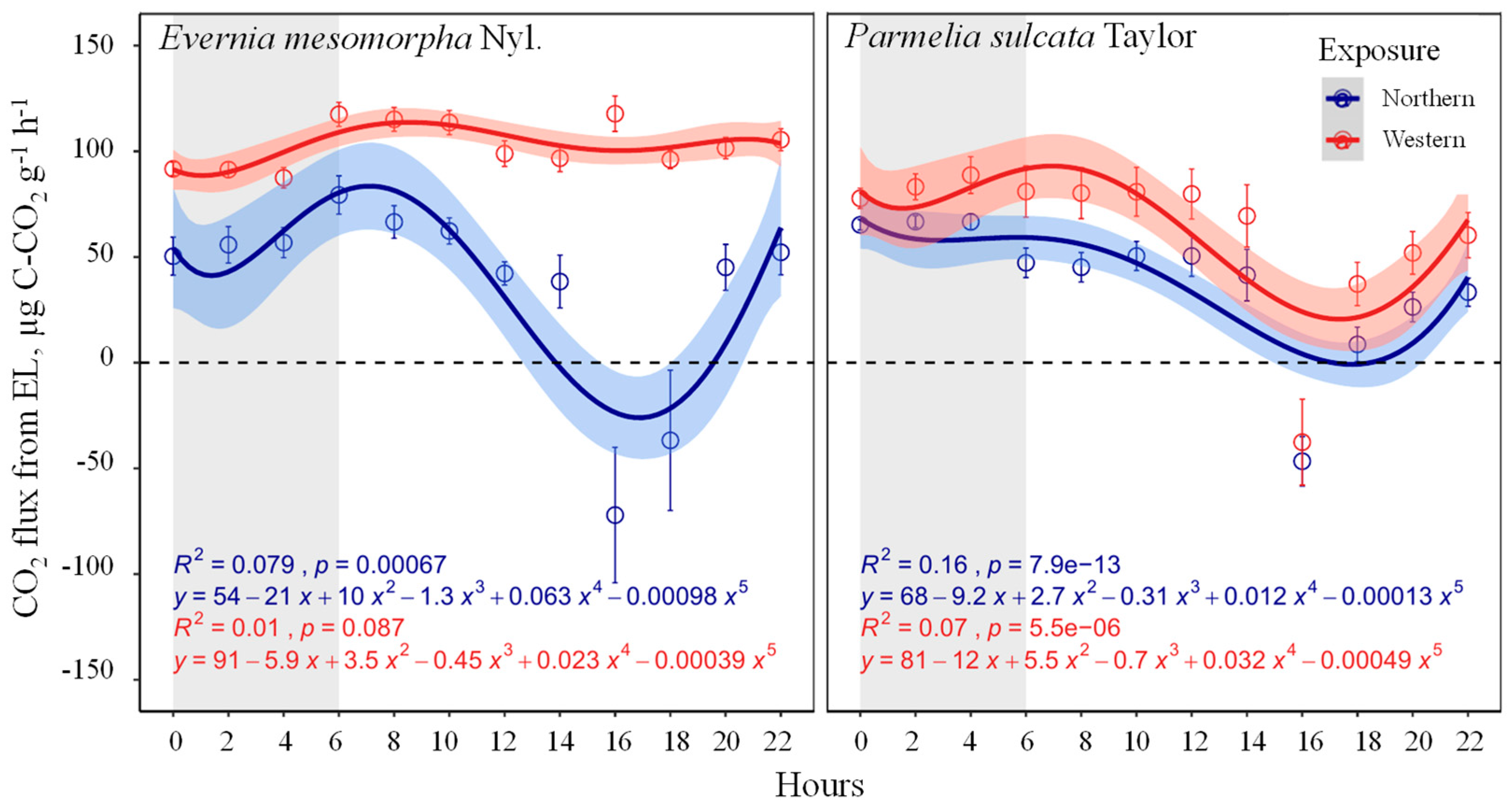
3.2. Diel Dynamics of CO2 Fluxes by the ELs
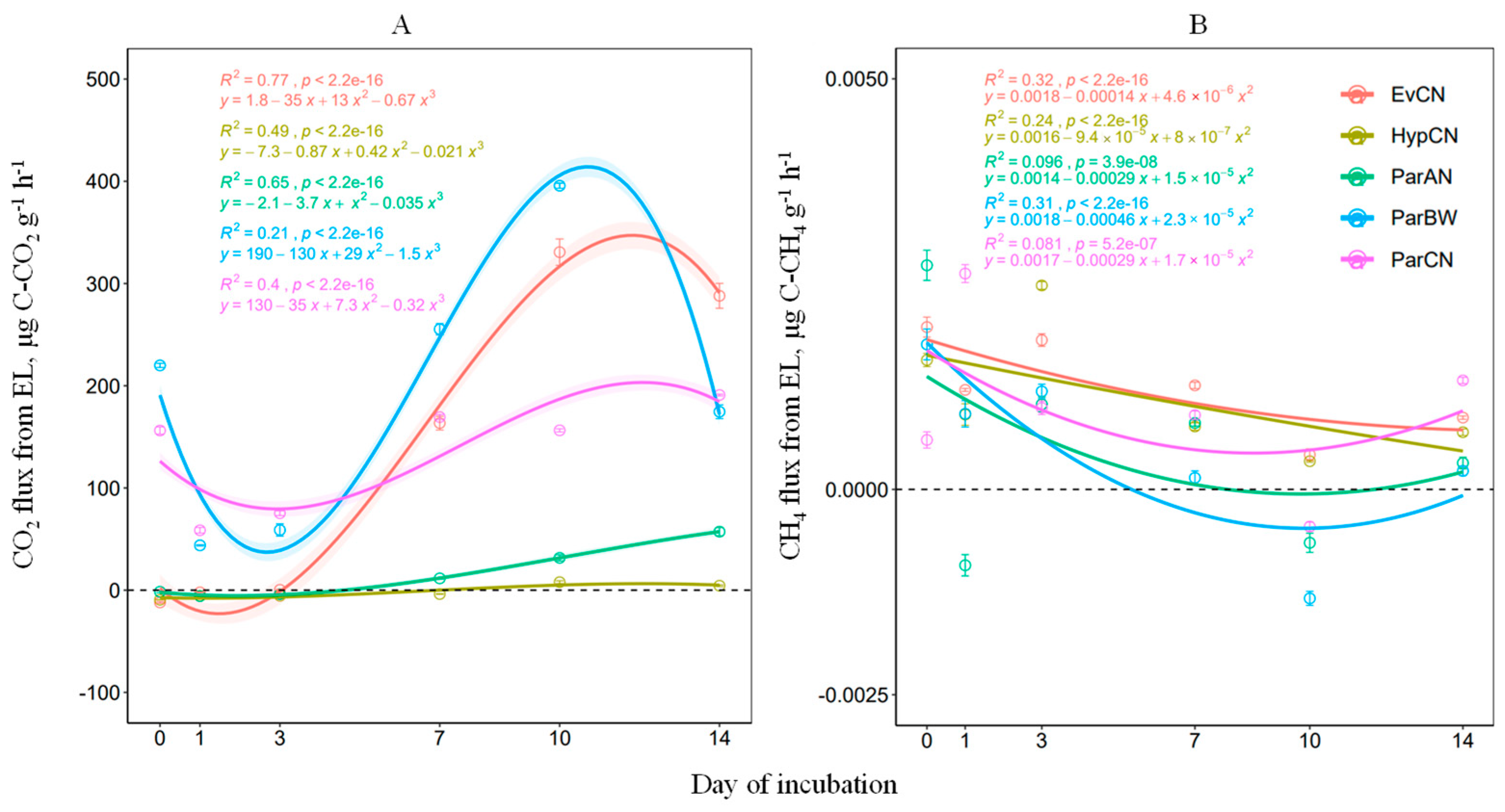
3.3. Assessment of CH4 and CO2 Fluxes by the ELs
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EL Species | Phorophyte | Geographical Location | GHG Fluxes in Author Units | GHG Fluxes in Our Units, μg C-CO2 or μg C-CH4 per g of EL Dry Weight h−1 | Source |
|---|---|---|---|---|---|
| CO2 | |||||
| Parmelia sulcata Taylor, Hypogymnia physodes (L.) Nyl., Evernia mesomorpha Nyl. | Betula pubescens Ehrh. | Krasnoyarsk, Russia | from +0.1 to +510.8; from −1.0 to −23.4 | This study | |
| Lobaria pulmonaria | Taiga zone of the European northeast of Russia | from +0.2 † to +4.9; from −0.1 ‡ to −5.5 mg CO2 g−1 of the dry weight of EL h−1 | from +54.5 to +1336.4; from −27.3 to −1500 | [80,82] | |
| Lobaria pulmonaria | Aspen forest mixed with spruce and fir | Syktyvkar, Russia, 61.57 N, 50.55 E | from −1.7 to −5 µmol CO2 m−2 s−1 | [79,88] | |
| Parmelia sulcata | Castanea sativa Miller | Abbadia San Salvatore, Tuscany, Italy, 42.90 N, 11.65 E | from +0.36 to +1.91; from −4.5 to −8.7 mg CO2 g−1 h−1 * | from +98.2 to +520.9; from −1227.3 to −2372.7 | [84] |
| Peltigera canina, Stereocaulon paschale | Spruce–lichen forest | Southern Ontario, Canada, 44 N, 80 E | from +1.25 to +2.75 mg CO2 g dry wt−1 h−1 | from +341 to +477 | [83] |
| Lobaria pulmonaria, Evernia prunastri, Lobaria scrobiculata, Peltigera canina | Deciduous sub-med. forest; temperate forest | Portugal (Continental and Madeira Island) | from +0.25 to +0.36; from −4.50 to −4.80 nmol CO2 g dry wt−1 s−1 | from +39.6 to +57.02; from −712.8 to −760.3 | [85] |
| Ramalina maciforrnis | Negev desert, Israel | from −0.2 to +10 mg CO2 g−1 h−1 ** | from −54.5 to +2727.3 | [89] | |
| Ramalina menziesii | Central California, USA | −15 mg CO2 g−1 dry wt d−1 * | −170.5 | [90] | |
| Sticta filix, Pseudocyphellaria faveolata | Nothofagus menziesii, Ixerba brexioides, Dacrydium cupressinum | Aniwaniwa river valley, New Zealand, 38.75 S, 177.15 E | from −12.41 to −14.97 nmol CO2 g−1 s−1 | from −536.1 to −646.7 | [91] |
| Erioderma pedicellatum | Picea abies | Rendalen, Norway, 61.87 N, 10.85 E | from +1.14 to +1.49; from −2.5 to −6.1 µmol CO2 m−2 s−1 | [92] | |
| Evernia mesomorpha, Bryoria nadvornikiana | Picea mariana | Northern Clay Belt, Canada, 50 N, 77–78 W | from −0.628 to −0.704 **,***; from +0.359 to +0.456 *** | [50] | |
| Lobaria crenulata, Pseudocyphellaria aurata, P. intricaia, Sticta sublimbata, S. weigelii | Premontane tropical rainforest | Republic of Panama, 8.75 N, 82.25 W | From −2.90 to −12.96 **; from 2.72 to 7.66 mg C (g C)−1 d−1 | [93] | |
| CH4 | |||||
| Parmelia sulcata Taylor, Hypogymnia physodes (L.) Nyl., Evernia mesomorpha Nyl. | Betula pubescens Ehrh. | Krasnoyarsk, Russia | from +0.00006 to +0.0061; from −0.00004 to −0.003 | This study | |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmajian, V. Lichens are more important than you think. Bioscience 1995, 45, 124. Available online: https://scholar.google.com/scholar?cluster=17933703524925869523&hl=ru&as_sdt=0,5 (accessed on 7 October 2023).
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.W.; Davey, S.; Peace, A.J.; Ferris, R.; Harding, K. Lichens and bryophyte communities of planted and semi-natural forests in Britain: The influence of site type, stand structure and deadwood. Biol. Conserv. 2002, 107, 165–180. [Google Scholar] [CrossRef]
- Sedia, E.G.; Ehrenfeld, J.G. Lichens and mosses promote alternate stable plant communities in the New Jersey pinelands. Oikos 2003, 100, 447–458. [Google Scholar] [CrossRef]
- Friedel, A.; Oheimb, G.; von Dengler, J.; Härdtle, W. Species diversity and species composition of epiphytic bryophytes and lichens—A comparison of managed and unmanaged beech forests in NE Germany. Feddes Repert. 2006, 177, 172–185. [Google Scholar] [CrossRef]
- Cleavitta, N.L.; Dibbleb, A.C.; Werierc, D.A. Influence of tree composition upon epiphytic macrolichens and bryophytes in old forests of Acadia National Park, Maine. Bryologist 2009, 112, 467–487. [Google Scholar] [CrossRef]
- Hilmo, O.; Holien, H.; Hytteborn, H.; Ely-Aalstrup, H.R. Of epiphytic lichens in differently aged Picea abies plantations situated in the oceanic region of Central Norway. Lichenologist. 2009, 41, 97–108. [Google Scholar] [CrossRef]
- Jüriado, I.; Liira, J.; Paal, J. Diversity of epiphytic lichens in boreo-nemoral forests on the North-Estonian limestone escarpment: The effect of tree level factors and local environmental conditions. Lichenologist 2009, 41, 81–96. [Google Scholar] [CrossRef]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Ellis, C.J.; Asplund, J.; Benes Peri, R.; Branquinho, C.; Di Nuzzo, L.; Hurtado, P.; Martínez, I.; Matos, P.; Nascimbene, J.; Pinho, P.; et al. Functional traits in lichen ecology: A review of challenge and opportunity. Microorganisms 2021, 9, 766. [Google Scholar] [CrossRef]
- Dubay, S.A.; Hayward, G.D.; Martinez del Rio, C. Nutritional value and diet preference of arboreal lichens and hypogeous fungi for small mammals in the Rocky Mountains. Can. J. Zool. 2008, 86, 851–862. [Google Scholar] [CrossRef]
- Martinez, J.J.I.; Raz, R.; Mgocheki, N.; Álvarez, R. Epiphytic lichen is associated with species richness of gall-inducing aphids but not with niche differentiation among them. Arthropod-Plant Interact. 2014, 8, 17–24. [Google Scholar] [CrossRef]
- Lakatos, M. Lichens and bryophytes: Habitats and species. In Plant Desiccation Tolerance. Ecological Studies (Analysis and Synthesis); Lüttge, U., Beck, E., Bartels, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 215. [Google Scholar] [CrossRef]
- Conti, M.E.; Cecchetti, G. Biological monitoring: Lichens as bioindicators of air pollution assessment—A review. Environ. Pollut. 2001, 114, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Insarov, G. A method for detecting large-scale environmental change with lichens. In Monitoring with Lichens—Monitoring Lichens; Springer: Dordrecht, The Netherlands, 2002; pp. 399–403. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Daba, G.M. Lichens—Masters of extraordinary symbiosis with potent pharmaceuticals. Egypt Pharmaceut. J. 2020, 19, 197–201. [Google Scholar] [CrossRef]
- Shah, A.A.; Badshah, L.; Muhammad, M.; Basit, A.; Ullah, I.; Mohamed, H.I.; Khan, A. Secondary metabolites of lichens and their application. In Fungal Secondary Metabolites; Elsevier: Amsterdam, The Netherlands, 2024; pp. 91–115. [Google Scholar] [CrossRef]
- Kovaleva, N.M. Epiphytic lichen phytomass distribution on the Scotch pine (Pinus sylvestris L.) stems and branches. Vestnik KrasGAU. 2012, 3, 90–95. (In Russian) [Google Scholar]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Keppler, F.; Boros, M.; Frankenberg, C.; Lelieveld, J.; McLeod, A.; Pirttilä, A.M.; Röckmann, T.; Schnitzler, J. Methane formation in aerobic environments. Environ. Chem. 2009, 6, 459–465. [Google Scholar] [CrossRef]
- Kip, N.; Winden, J.; Pan, Y.; Bodrossy, L.; Reichart, G.J.; Smolders, A.J.P.; Jetten, S.M.M.; Sinninghe Damsté, J.S.; Op den Camp, H.J.M. Global prevalence of methane oxidation by symbiotic bacteria in peat-moss ecosystems. Nat. Geosci. 2010, 3, 617–621. [Google Scholar] [CrossRef]
- Johansson, O.; Palmqvist, K.; Olofsson, J. Nitrogen deposition drives lichen community changes through differential species responses. Glob. Chang. Biol. 2012, 18, 2626–2635. [Google Scholar] [CrossRef]
- Lenhart, K.; Bunge, M.; Ratering, S.; Neu, T.R.; Schuttmann, I.; Greule, M.; Kammann, C.; Schnell, S.; Müller, C.; Zorn, H.; et al. Evidence for methane production by saprotrophic fungi. Nat. Commun. 2012, 3, 1046. [Google Scholar] [CrossRef]
- Lenhart, K.; Weber, B.; Elbert, W.; Steinkamp, J.; Clough, T.; Crutzen, P.; Pöschl, U.; Keppler, F. Nitrous oxide and methane emissions from cryptogamic covers. Glob. Chang. Biol. 2015, 21, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, K.; Klintzsch, T.; Langer, G.; Nehrke, G.; Bunge, M.; Schnell, S.; Keppler, F. Evidence for methane production by marine algae (Emiliana huxleyi) and its implication for the methane paradox in oxic waters. Biogeosciences 2016, 13, 3163–3174. [Google Scholar] [CrossRef]
- Masyagina, O.V.; Menyailo, O.V. The impact of permafrost on carbon dioxide and methane fluxes in Siberia: A meta-analysis. Environ. Res. 2020, 182, 109096. [Google Scholar] [CrossRef] [PubMed]
- Beckett, R.P.; Kranner, I.; Minibayeva, F.V. Stress physiology and the symbiosis. In Lichen Biology, 2nd ed.; Nash, T.H., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 134–151. [Google Scholar] [CrossRef]
- Beckett, R.P.; Minibayeva, F.; Solhaug, K.A.; Roach, T. Photoprotection in lichens: Adaptations of photobionts to highlight. Lichenologist 2021, 53, 21–33. [Google Scholar] [CrossRef]
- Zelikova, T.J.; Housman, D.C.; Grote, E.E.; Neher, D.A.; Belnap, J. Warming and increased precipitation frequency on the Colorado Plateau: Implications for biological soil crusts and soil processes. Plant Soil 2012, 355, 265–282. [Google Scholar] [CrossRef]
- Goga, M.; Elečko, J.; Marcinčinová, M.; Ručová, D.; Bačkorová, M.; Bačkor, M. Lichen metabolites: An overview of some secondary metabolites and their biological potential. In Co-Evolution of Secondary Metabolites; Merillon, J.-M., Ramawal, K.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 175–209. [Google Scholar] [CrossRef]
- Lücking, R.; Leavitt, S.D.; Hawksworth, D.L. Species in lichen-forming fungi: Balancing between conceptual and practical considerations, and between phenotype and phylogenomics. Fungal Divers 2021, 109, 99–154. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Zarabska-Bożjewicz, D. Hypogymnia physodes–A lichen with interesting medicinal potential and ecological properties. J. Herb. Med. 2019, 17, 100287. [Google Scholar] [CrossRef]
- Bižić, M.; Klintzsch, T.; Ionescu, D.; Hindiyeh, M.Y.; Günthel, M.; Muro-Pastor, A.M.; Eckert, W.; Urich, T.; Keppler, F.; Grossart, H.-P. Aquatic and terrestrial cyanobacteria produce methane. Sci. Adv. 2020, 6, eaax5343. [Google Scholar] [CrossRef]
- Samylina, O.S.; Rusanov, I.I.; Tarnovetskii, I.Y.; Yakushev, E.V.; Grinko, A.A.; Zakharova, E.E.; Merkel, A.Y.; Kanapatskiy, T.A.; Semiletov, I.P.; Pimenov, N.V. On the possibility of aerobic methane production by pelagic microbial communities of the Laptev Sea. Microbiology 2021, 90, 145–157. [Google Scholar] [CrossRef]
- Shver, T.A.; Gerasimova, A.S. Climate of Krasnoyarsk; Gidrometeoizdat: Krasnoyarsk, Russia, 1982; p. 230. (In Russian) [Google Scholar]
- Bezrukih, V.A.; Elin, O.Y.; Dudnik, N.I. The natural and social background of the Yenisei Siberia agricultural areas economic growth. Vestn. Tomsk. Gos. Univ. 2009, 14, 407–411. (In Russian) [Google Scholar]
- Torzhkov, I.O.; Kushnir, E.A.; Konstantinov, A.V.; Koroleva, T.S.; Efimov, S.V.; Shkolnik, I.M. Assessment of future climate change impacts on forestry in Russia. Russ. Meteorol. Hydrol. 2019, 44, 180–186. [Google Scholar] [CrossRef]
- Kryuchkova, O.E. Epiphytic lichen flora of the city in connection with the acidity of tree bark and air pollution (using the example of Krasnoyarsk). In Abstract of the Dissertation for the Scientific Degree of Candidate of Biological Sciences; GOUVPO “Krasnoyarsk State University”: Krasnoyarsk, Russia, 2006. (In Russian) [Google Scholar]
- Hauck, M.; Helms, G.; Friedl, T. Photobiont selectivity in the epiphytic lichens Hypogymnia physodes and Lecanora conizaeoides. Lichenologist 2007, 39, 195–204. [Google Scholar] [CrossRef]
- Piercey-Normore, M.D. Vegetatively reproducing fungi in three genera of the Parmeliaceae share divergent algal partners. Bryologist 2009, 112, 773–785. Available online: https://www.jstor.org/stable/25614897 (accessed on 25 September 2023). [CrossRef]
- Rocha, B.; Matos, P.; Giordani, P.; Piret, L.; Branquinho, C.; Casanelles-Abella, J.; Aleixo, C.; Deguines, N.; Hallikma, T.; Laanisto, L.; et al. Modelling the response of urban lichens to broad-scale changes in air pollution and climate. Environ. Pollut. 2022, 315, 120330. [Google Scholar] [CrossRef] [PubMed]
- Lebedeva, N.V.; Krivolutsky, D.A.; Puzachenko, Y.G.; Dyakonov, K.N.; Aleshchenko, G.M.; Smurov, A.V.; Maksimov, V.N.; Tikunov, V.S.; Ogureeva, G.N.; Kotova, T.V. Geography and Monitoring of Biodiversity; Publishing House of the Scientific and Educational Methodological Center: Moscow, Russia, 2002; 432p. (In Russian) [Google Scholar]
- Trass, H. Lichen sensitivity to the air pollution and index of poleotolerance (IP). Folia Cryptog. Estonica. 1973, 3, 19–22. Available online: https://ojs.utlib.ee/index.php/FCE/issue/view/959/48 (accessed on 19 December 2023).
- Fadel, D.; Sid, A.S.; Zga, N.; Latrèche, F.; Ali, A.O. Cartography of Air Pollution in an Industrial City in North-Eastern Algeria by Using Two Indexes: Poleotolerance Index and Atmospheric Purity Index. J. Life Sci. 2014, 8, 95–100. [Google Scholar]
- Rocha, B.; Pinho, P.; Vieira, J.; Branquinho, C.; Matos, P. Testing the poleotolerance lichen response trait as an indicator of anthropic disturbance in an urban environment. Diversity 2019, 11, 55. [Google Scholar] [CrossRef]
- Fazan, L.; Gwiazdowicz, D.J.; Fragnière, Y.; Fałtynowicz, W.; Ghosn, D.; Remoundou, I.; Rusińska, A.; Urbański, P.; Pasta, S.; Garfì, G.; et al. Factors influencing the diversity and distribution of epiphytic lichens and bryophytes on the relict tree Zelkova abelicea (Lam.) Boiss. (Ulmaceae). Lichenologist 2022, 54, 195–212. [Google Scholar] [CrossRef]
- Letardi, P.; Ramirez Barat, B.; Cano, E. Analysis of the influence of the electrochemical cell setup for corrosion measurements on metallic cultural heritage. In Proceedings of the European Corrosion Congress-EUROCORR, Prague, Czech Republic, 3–7 September 2017; Available online: https://www.researchgate.net/profile/Paola-Letardi/publication/319979704_Analysis_of_the_influence_of_the_electrochemical_cell_setup_for_corrosion_measurements_on_metallic_cultural_heritage/links/59c4c3a1a6fdccc719137b89/Analysis-of-the-influence-of-the-electrochemical-cell-setup-for-corrosion-measurements-on-metallic-cultural-heritage.pdf (accessed on 19 December 2023).
- Palmqvist, K. Carbon economy in lichens. New Phytol. 2000, 148, 11–36. [Google Scholar] [CrossRef]
- Lange, O.L.; Green, T.A.; Heber, U. Hydration-dependent photosynthetic production of lichens: What do laboratory studies tell us about field performance? J. Exp. Bot. 2001, 52, 2033–2042. [Google Scholar] [CrossRef]
- Boudreault, C.; Coxson, D.; Bergeron, Y.; Stevenson, S.; Bouchard, M. Do forests treated by partial cutting provide growth conditions similar to old-growth forests for epiphytic lichens? Biol. Conserv. 2013, 159, 458–467. [Google Scholar] [CrossRef]
- Wiessner, W. Photoassimilation of Organic Compounds. In Photosynthesis II. En-Cyclopedia of Plant Physiology; Gibbs, M., Latzko, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1979; Volume 6. [Google Scholar] [CrossRef]
- Duan, Z.; Homma, A.; Kobayashi, M.; Nagata, N.; Kaneko, Y.; Fujiki, Y.; Nishida, I. Photoassimilation, assimilate translocation and plasmodesmal biogenesis in the source leaves of Arabidopsis thaliana grown under an increased atmospheric CO2 concentration. Plant Cell Physiol. 2014, 55, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.P. Practical Statistics for Data Scientists: 50+ Essential Concepts Using R and Python; Bruce, P., Bruce, A., Gedeck, P., Eds.; O’Reilly Media Inc.: Boston, MA, USA, 2021; pp. xiii+368. ISBN 9781492072942. [Google Scholar] [CrossRef]
- Dubrovsky, G.I. Materials on the lichen flora of the state reserve “Stolby” of the Krasnoyarsk Territory. Botanical materials of the department of spore plants. Mosc. Leningrad. 1953, 9, 31–39. (In Russian) [Google Scholar]
- Perova, S.V. Lichens in the vicinity of Krasnoyarsk. In Collection of Scientific Student Works; Krasnoyarsk Pedagogical Institute: Krasnoyarsk, Russia, 1961; Volume 1, pp. 45–52. (In Russian) [Google Scholar]
- Otnyukova, T.N. The find of Ramalina sinensis (Ramalinaceae, Lichenes) in Altai province. Bot. Zhurnal. 1998, 83, 132–136. (In Russian) [Google Scholar]
- Otnyukova, T.N. Some interesting and rare species of mosses and lichens of the Yenisei Siberia. Conservation of biological diversity of the Yenisei Siberia. In Proceedings of the First Interregional Scientific and Practical Conference on the Conservation of Biodiversity of the Yenisei Siberia. Part 2, Krasnoyarsk, Russia, 28–30 November 2000; pp. 43–44. (In Russian). [Google Scholar]
- Otnyukova, T. Epiphytic lichen growth abnormalities and element concentrations as early indicators of forest decline. Environ. Pollut. 2007, 146, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.J.; Coppins, B.J. Reproductive strategy and the compositional dynamics of crustose lichen communities on aspen (Populus tremula L.) in Scotland. Lichenologist 2007, 39, 377–391. [Google Scholar] [CrossRef]
- Mežaka, A.; Brumelis, G.; Piterans, A. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. Biodiv. Conserv. 2012, 21, 3221–3241. [Google Scholar] [CrossRef]
- Shriver, R.K.; Cutler, K.; Doak, D.F. Comparative demography of an epiphytic lichen: Support for general life history patterns and solutions to common problems in demographic parameter estimation. Oecologia 2012, 170, 137–146. [Google Scholar] [CrossRef]
- Kuusinen, M. Epiphytic lichen flora and diversity on Populus tremula in old-growth and managed forests of southern and middle boreal Finland. Ann. Bot. Fenn. 1994, 31, 245–260. Available online: https://www.jstor.org/stable/43922219 (accessed on 19 December 2023).
- Kuusinen, M.; Siitonen, J. Epiphytic lichen diversity in old-growth and managed Picea abies stands in southern Finland. J. Veg. Sci. 1998, 9, 283–292. [Google Scholar] [CrossRef]
- Cameron, R.P. Habitat associations of epiphytic lichens in managed and unmanaged forest stands in Nova Scotia. Northeast Natur. 2002, 9, 27–46. [Google Scholar] [CrossRef]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of tree age and microhabitat quality on the occurrence of rhytidomeose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Lie, M.H.; Arup, U.; Grytnes, J.-A.; Ohlson, M. The importance of host tree age, size and growth rate as determinants of epiphytic lichen diversity in boreal spruce forests. Biodiv. Conserv. 2009, 18, 3579–3596. [Google Scholar] [CrossRef]
- Calviño-Cancela, M.; López de Silanes, M.E.; Rubido-Bará, M.; Uribarri, J. The potential role of tree plantations in providing habitat for lichen epiphytes. For. Ecol. Manag. 2013, 291, 386–395. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; García-Franco, J. The relationship between tree size and epiphyte species richness: Testing four different hypotheses. J. Biogeogr. 2006, 33, 323–333. [Google Scholar] [CrossRef]
- Mežaka, A.; Brūmelis, G.; Piterāns, A. The distribution of epiphytic bryophyte and lichen species in relation to phorophyte characters in Latvian natural old-growth broad-leaved forests. Folia Cryptog. Estonica 2008, 44, 89–99. [Google Scholar]
- Fritz, Ö. Vertical distribution of epiphytic bryophytes and lichens emphasizes the importance of old beeches in conservation. Biodiv. Conserv. 2009, 18, 289–304. [Google Scholar] [CrossRef]
- Istomina, N.B.; Likhacheva, O.V. Lichen biota of manor parks with federal protection status in the Pskov region. Bull. Russ. State Univ. Named I Kant 2010, 7, 122–129. (In Russian) [Google Scholar]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, S.B.; Woo, D.; Bae, G.N. Influence of wind direction and speed on the transport of particle-bound PAHs in a roadway environment. Atm. Pollut. Res. 2015, 6, 1024–1034. [Google Scholar] [CrossRef]
- Cortes, D.R.; Basu, I.; Sweet, C.W.; Hites, R.A. Temporal trends in and influence of wind on PAH concentrations measured near the Great Lakes. Environ. Sci. Technol. 2000, 34, 356–360. [Google Scholar] [CrossRef]
- Huang, Y.D.; Hou, R.W.; Liu, Z.Y.; Song, Y.; Cui, P.Y.; Kim, C.N. Effects of wind direction on the airflow and pollutant dispersion inside a long street canyon. Aerosol Air Qual. Res. 2019, 19, 1152–1171. [Google Scholar] [CrossRef]
- Tsurykau, A.H.; Khramchankova, V.M. Influence of the bark acidity on frequency of lichens (on an example of Gomel Town). Nov. Sist. Nizs. Rast. 2009, 43, 261–275. Available online: https://www.researchgate.net/publication/346499881_Influence_of_the_bark_acidity_on_frequency_of_lichens_on_an_example_of_Gomel_Town (accessed on 19 December 2023). [CrossRef]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Ramawat, K., Merillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar] [CrossRef]
- Rankovic, B.; Kosanic, M. Lichens as a potential source of bioactive secondary metabolites. In Lichen Secondary Metabolites; Rankovic, B., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Shelyakin, M.A.; Zakhozhiy, I.G.; Golovko, T.K. Changes of total respiration and respiratory pathways ratio in lichens adaptation to UV-b radiation. Proc. RAS Ufa Sci. Cent. 2018, 3, 100–104. (In Russian) [Google Scholar] [CrossRef]
- Golovko, T.K.; Shelyakin, M.A.; Pystina, T.N. Ecological and biological, and functional traits of lichens in Taiga zone of European Northeast of Russia. Theor. Appl. Ecol. 2020, 1, 6. Available online: http://envjournal.ru/archives/2020_1.pdf (accessed on 19 December 2023). [CrossRef]
- Khabibrakhmanova, V.R.; Rassabina, A.Y.; Khayrullina, A.F.; Minibayeva, F.V. Physico-chemical characteristics and antioxidant properties of melanins extracted from Leptogonium furfuraceum (Harm.). Khimiya Rastit. Syr’ya 2022, 4, 117–127. (In Russian) [Google Scholar] [CrossRef]
- Golovko, T.K.; Dalke, I.V.; Dymova, O.V.; Zakhozhiy, I.G.; Malyshev, R.V.; Silina, E.V.; Tabalenkova, G.N.; Shelyakin, M.A. First results of the ecological and physiological study of lichen biota of the boreal zone of the European north-east of Russia. In Current State and Prospects for the Development of a Network of Specially Protected Natural Territories of the European North and the Urals; IB Komi Scientific Center Ural Branch RAS: Syktyvkar, Russia, 2015; pp. 279–286. (In Russian) [Google Scholar]
- Crittenden, P.D.; Kershaw, K.A. A procedure for the simultaneous measurement of net CO2-exchange and nitrogenase activity in lichens. New Phytol. 1978, 80, 393–401. [Google Scholar] [CrossRef]
- Tretiach, M.; Bertuzzi, S.; Candotto Carniel, F.; Virgilio, D. Seasonal acclimation in the epiphytic lichen Parmelia sulcata is influenced by change in photobiont population density. Oecologia 2013, 173, 649–663. [Google Scholar] [CrossRef]
- Máguas, C.; Griffiths, H.; Broadmeadow, M.S.J. Gas exchange and carbon isotope discrimination in lichens: Evidence for interactions between CO2-concentrating mechanisms and diffusion limitation. Planta 1995, 196, 95–102. [Google Scholar] [CrossRef]
- Segers, R. Methane production and methane consumption: A review of processes underlying wetland methane fluxes. Biogeochemistry 1998, 41, 23–51. [Google Scholar] [CrossRef]
- Liu, D.Y.; Ding, W.X.; Jia, Z.J.; Cai, Z.C. Relation between methanogenic archaea and methane production potential in selected natural wetland ecosystems across China. Biogeosciences 2011, 8, 329–338. [Google Scholar] [CrossRef]
- Shelyakin, M.A.; Zakhozhiy, I.G.; Dalke, I.V.; Dymova, O.V.; Malyshev, R.V.; Golovko, T.K. Photosynthetic and respiratory capacity of foliose lichen Lobaria pulmonaria throughout the annual cycle. Russ. J. Plant Physiol. 2021, 68, 1048–1058. [Google Scholar] [CrossRef]
- Lange, O.L.; Tenhunen, J.D. Moisture content and CO2 exchange of lichens. II. Depression of net photosynthesis in Ramalina maciformis at high water content is caused by increased thallus carbon dioxide diffusion resistance. Oecologia 1981, 51, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Matthes-Sears, U.; Nash III, T.H.; Larson, D.W. The ecology of Ramalina menziesii. III. In situ diurnal field measurements at two sites on a coast–inland gradient. Can. J. Bot. 1986, 64, 988–996. [Google Scholar] [CrossRef]
- Green, T.A.; Meyer, A.; Buedel, B.; Zellner, H.; Lange, O.L. Diel patterns of CO2-exchange for six lichens from a temperate rain forest in New Zealand. Symbiosis 1995, 18, 251–273. Available online: https://dalspace.library.dal.ca/bitstream/handle/10222/77482/VOLUME%2018-NUMBER%203-1995-PAGE%20251.pdf?sequence=1 (accessed on 19 December 2023).
- Nilsson, A.R.; Solhaug, K.A.; Gauslaa, Y. The globally threatened epiphytic cyanolichen Erioderma pedicellatum depends on a rare combination of habitat factors. Lichenologist 2022, 54, 123–136. [Google Scholar] [CrossRef]
- Lange, O.L.; Büdel, B.; Meyer, A.; Zellner, H.; Zotz, G. Lichen carbon gain under tropical conditions: Water relations and CO2 exchange of Lobariaceae species of a lower montane rainforest in Panama. Lichenologist 2004, 36, 329–342. [Google Scholar] [CrossRef]




| Plot | Tree | Tree Height, m | DBH, cm | Canopy Cover, % | Age, Years |
|---|---|---|---|---|---|
| A | A1 | 25.3 | 67.7 | 54 | 111 |
| A2 | 28.0 | 47.8 | 58 | 106 | |
| A3 | 25.5 | 42.9 | 75 | 94 | |
| A4 | 28.5 | 42.5 | 68 | 100 | |
| B | B5 | 25.9 | 37.3 | 64 | 118 |
| B6 | 26.0 | 47.7 | 70 | 117 | |
| B7 | 32.9 | 47.1 | 86 | 115 | |
| B8 | 27.9 | 58.6 | 72 | 101 | |
| C | C9 | 29.3 | 35.4 | 65 | 95 |
| C10 | 33.8 | 38.2 | 70 | 102 | |
| C11 | 32.0 | 36.8 | 49 | 98 | |
| C12 | 24.2 | 41.9 | 60 | 115 |
| Plot | Projecting Cover (Mean ± SE), % | Occurrence, % | Total EL Projecting Cover (as an Average Value of 20 Slots ± SE) at a Height of 1–1.5 m, % | ||||
|---|---|---|---|---|---|---|---|
| P | H | Ev | P | H | Ev | ||
| A1 | 13.5 ± 3.8 | 2.5 ± 1.2 | 0.5 ± 0.3 | 50 | 25 | 10 | 33.5 ± 4.1 |
| A2 | 5.2 ± 1.6 | 3.0 ± 1.6 | 3.0 ± 1.1 | 45 | 20 | 40 | 28.9 ± 3.8 |
| A3 | 6.5 ± 1.3 | 0.5 ± 0.5 | 1.0 ± 0.5 | 65 | 5 | 20 | 48.3 ± 3.5 |
| A4 | 11.8 ± 3.0 | 1.5 ± 0.6 | 8.5 ± 2.3 | 60 | 25 | 75 | 40.1 ± 5.5 |
| B5 | 6.2 ± 1.8 | 1.5 ± 0.6 | 1.3 ± 1.0 | 50 | 25 | 10 | 38.5 ± 5.7 |
| B6 | 5.5 ± 1.5 | 0 ± 0 | 1.4 ± 0.5 | 45 | 0 | 30 | 36.2 ± 5.5 |
| B7 | 2.9 ± 1.2 | 4.0 ± 1.7 | 1.0 ± 0.4 | 30 | 25 | 25 | 24.1 ± 5.3 |
| B8 | 8.2 ± 1.9 | 1.5 ± 1.1 | 0.3 ± 0.3 | 65 | 10 | 5 | 29.4 ± 4.7 |
| C9 | 5.8 ± 1.8 | 2.7 ± 0.8 | 3.9 ± 1.0 | 45 | 45 | 55 | 22.4 ± 5.2 |
| C10 | 1.9 ± 0.4 | 3.0 ± 0.6 | 1.4 ± 0.4 | 65 | 80 | 60 | 27.8 ± 6.1 |
| C11 | 6.5 ± 1.7 | 1.4 ± 0.5 | 2.7 ± 0.6 | 90 | 55 | 85 | 15.8 ± 2.3 |
| C12 | 7.3 ± 1.9 | 3.3 ± 0.6 | 2.7 ± 0.9 | 60 | 70 | 45 | 26.6 ± 3.2 |
| Factors and Their Interactions | CO2 Fluxes | |
|---|---|---|
| F | p | |
| EL species | 10.1 | <0.001 |
| Photoperiod | 20.3 | <0.0001 |
| Exposure on the tree stem | 100.9 | <0.0001 |
| EL species × Photoperiod | 13.4 | <0.0001 |
| EL species × Exposure on the tree stem | 20.0 | <0.0001 |
| Photoperiod × Exposure on the tree stem | 10.5 | <0.001 |
| EL species × Photoperiod × Exposure on the tree stem | 3.1 | 0.76 |
| Factors and Their Interactions | CH4 | CO2 | ||
|---|---|---|---|---|
| F | p | F | p | |
| EL species | 0.698 | 0.610 | 13.77 | <0.0001 |
| Time | 5.158 | <0.01 | 21.06 | <0.0001 |
| EL species × Time | 0.903 | 0.585 | 2.25 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matvienko, A.I.; Evgrafova, S.Y.; Kovaleva, N.M.; Sideleva, E.V.; Sitnikova, M.V.; Menyailo, O.V.; Masyagina, O.V. Greenhouse Gas Fluxes from the Epiphytic Lichens: Incubation Experiments. Forests 2024, 15, 107. https://doi.org/10.3390/f15010107
Matvienko AI, Evgrafova SY, Kovaleva NM, Sideleva EV, Sitnikova MV, Menyailo OV, Masyagina OV. Greenhouse Gas Fluxes from the Epiphytic Lichens: Incubation Experiments. Forests. 2024; 15(1):107. https://doi.org/10.3390/f15010107
Chicago/Turabian StyleMatvienko, Anastasia I., Svetlana Y. Evgrafova, Natalia M. Kovaleva, Elizaveta V. Sideleva, Maria V. Sitnikova, Oleg V. Menyailo, and Oxana V. Masyagina. 2024. "Greenhouse Gas Fluxes from the Epiphytic Lichens: Incubation Experiments" Forests 15, no. 1: 107. https://doi.org/10.3390/f15010107