
Soil Microbial Community and Soil Abiotic Factors Are Linked to Microorganisms’ C:N:P Stoichiometry in Larix Plantations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Sampling and Laboratory Analysis
2.3. The Characteristics of Microbial Communities Analysis
2.4. Statistical Analyses
3. Results
3.1. Variation in Environmental Parameters with Plantation Age
3.2. The Characteristics of Soil and Microorganisms
3.3. The Stoichiometric Ratios of Elements in Soil and Microbial Biomass
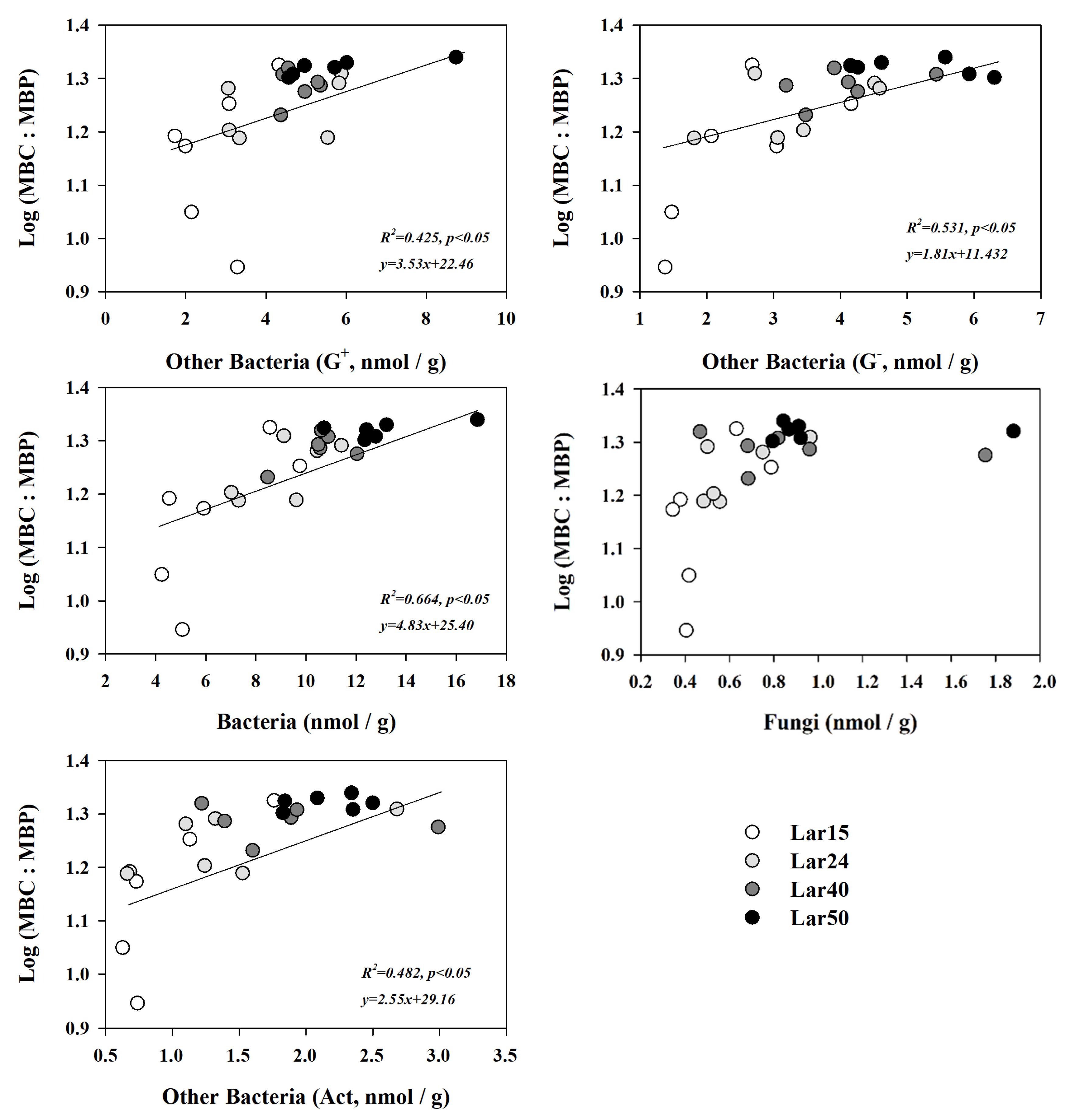
3.4. Combined Effects of Soil Properties and Microbial Communities on Microbial Stoichiometric Ratios
4. Discussion
4.1. Shifts in Basic Environmental Variables under Different Plantation Ages
4.2. The Influence of Forest Age on the Soil Elemental Stoichiometric Ratios and Microbial Characteristics
4.3. Strong Linkages between Microbial C:N:P Ratios, Soil Properties, and Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peñuelas, J.; Sardans, J. Elementary factors. Nature 2009, 460, 803–804. [Google Scholar] [CrossRef]
- Vicca, S.; Stocker, B.D.; Reed, S.; Wieder, W.R.; Bahn, M.; Fay, P.A.; Janssens, I.A.; Lambers, H.; Peñuelas, J.; Piao, S. Using research networks to create the comprehensive datasets needed to assess nutrient availability as a key determinant of terrestrial carbon cycling. Environ. Res. Lett. 2018, 13, 125006. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NY, USA, 2002. [Google Scholar]
- Heyburn, J.; McKenzie, P.; Crawley, M.J.; Fornara, D.A. Effects of grassland management on plant C:N:P stoichiometry: Implications for soil element cycling and storage. Ecosphere 2017, 8, e01963. [Google Scholar] [CrossRef]
- Fanin, N.; Fromin, N.; Buatois, B.; Hättenschwiler, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Magruder, M.; Chhin, S.; Palik, J.B.B. Thinning increases climatic resilience of red pine. Can. J. For. Res. 2013, 43, 878–889. [Google Scholar] [CrossRef]
- Hooper, D.U.; Bignell, D.E.; Brown, V.K.; Brussaard, L.; Dangerfield, J.M.; Wall, D.H.; Wardle, D.A.; Coleman, D.C.; Giller, K.E.; Lavelle, P.; et al. Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: Patterns, mechanisms, and feedbacks. BioScience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Bao, Y.; Qiuliang, Z.; Liming, W. Comprehensive characteristics of the vertical structure of middle young over cutting forest of Larix gmelinii. Sci. Silvae Sin. 2015, 51, 132–139. [Google Scholar]
- Lunstrum, A.; Chen, L. Soil carbon stocks and accumulation in young mangrove forests. Soil Biol. Biochem. 2014, 75, 223–232. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. Bioenergy 2010, 10, 2052–2077. [Google Scholar] [CrossRef]
- Hember, R.A.; Kurz, W.A.; Metsaranta, J.M.; Black, T.A.; Guy, R.D.; Coops, N.C. Accelerating regrowth of temperate-maritime forests due to environmental change. Glob. Chang. Biol. Bioenergy 2012, 18, 2026–2040. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Wang, S.; He, T.; Liu, L. Fresh carbon and nitrogen inputs alter organic carbon mineralization and microbial community in forest deep soil layers. Soil Biol. Biochem. 2014, 72, 145–151. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, X.; Wang, W. Variation of soil carbon pools in Pinus sylvestris, plantations of different ages in north China. Acta Ecol. Sin. 2018, 38, 248–254. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Chen, X.-L.; Wang, D.; Chen, X.; Wang, J.; Diao, J.-J.; Zhang, J.-Y.; Guan, Q.-W. Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation. Appl. Soil Ecol. 2015, 92, 35–44. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, Y.; An, S.S. How C:N:P stoichiometry in soils and plants responds to succession in Robinia pseudoacacia forests on the Loess Plateau, China. For. Ecol. Manag. 2020, 475, 118394. [Google Scholar] [CrossRef]
- Dang, P.; Gao, Y.; Liu, J.; Yu, S.; Zhao, Z. Effects of thinning intensity on understory vegetation and soil microbial communities of a mature Chinese pine plantation in the Loess Plateau. Sci. Total Environ. 2018, 630, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps inthe forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Chen, X.; Han, Y.H.; Chen, X.; Wang, J.; Chen, B.; Wang, D.; Guan, Q. Soil labile organic carbon and carbon-cycle enzyme activities under different thinning intensities in Chinese fir plantations. Appl. Soil Ecol. 2016, 107, 162–169. [Google Scholar] [CrossRef]
- Ma, J.Y.; Han, H.R.; Cheng, X.Q. Soil temperatures and active carbon components as key drivers of C stock dynamics between two different stand ages of Larix principis-rupprechtii plantation. Peerj 2020, 8, e8384. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B.; Kou, D.; Liu, L.; Fang, K.; Zhang, B.B.; et al. Linking microbial C:N:P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar] [CrossRef]
- Cheng, C.; Wang, Y.; Fu, X.; Xu, M.; Dai, X.; Wang, H. Thinning effect on understory community and photosynthetic characteristics in a subtropical Pinus massoniana plantation. Can. J. For. Res. 2017, 47, 1104–1115. [Google Scholar] [CrossRef]
- Li, H.L.; Crabbe, M.J.C.; Xu, F.L.; Wang, W.L.; Niu, R.L.; Gao, X.; Zhang, P.; Chen, H.K. Seasonal variations in carbon, nitrogen and phosphorus concentrations and C:N:P stoichiometry in the leaves of differently aged Larix principis-rupprechtii Mayr. Plantations. Forests 2017, 8, 373. [Google Scholar] [CrossRef]
- Zeng, Q.; Lal, R.; Chen, Y.; An, S. Leaf and root ecological stoichiometry of Caragana korshinskii on the Loess Plateau of China in relation to plantation age. PLoS ONE 2017, 12, e0168890. [Google Scholar] [CrossRef]
- Liang, X.; Liu, S.; Wang, H.; Wang, J. Variation of carbon and nitrogen stoichiometry along a chronosequence of natural temperate forest in northeastern China. J. Plant Ecol. 2017, 11, 339–350. [Google Scholar] [CrossRef]
- Sun, X.L.; Zhao, J.; You, Y.M.; Sun, O.J.X. Soil microbial responses to forest floor litter manipulation and nitrogen addition in a mixed-wood forest of northern China. Sci. Rep. 2015, 6, 19536. [Google Scholar] [CrossRef] [PubMed]
- FAO. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports No. 106; The Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Schumacher, B.A. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; US Environmental Protection Agency, Office of Research and Development, Ecological Risk Assessment Support Center: Washington, DC, USA, 2002.
- Zeng, Q.; Li, X.; Dong, Y.; An, S.; Darboux, F. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 2016, 147, 481–488. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Wu, J.S.; Lin, Q.M.; Huang, Q.Y.; Xiao, H.A. Soil Microbial Biomass–Methods and Applications; China Meteorological Press: Beijing, China, 2006. [Google Scholar]
- Hedley, M.J.; Stewart, J.W.B. Method to measure microbial phosphate in soils. Soil Biol. Biochem. 1982, 14, 377–385. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H.; Reich, P.B. Global-scale latitudinal patterns of plant fine root nitrogen and phosphorus. Nat. Commun. 2011, 2, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Mu, X.; Yuan, Z.; Deng, Q.; Chen, Y.; Yuan, L.Y.; Ryan, L.T.; Kallenbach, R.L. Soil nutrients and water affect the age-related fine root biomass but not production in two plantation forests on the Loess Plateau, China. J. Arid Environ. 2016, 135, 173–180. [Google Scholar] [CrossRef]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Aber, J.D.; Nadelhoffer, K.J.; Steudler, P.; Melillo, J.M. Nitrogen saturation in northern forest ecosystems. BioScience 1989, 39, 378–386. [Google Scholar] [CrossRef]
- Tian, L.M.; Zhao, L.; Wu, X.D.; Fang, H.B.; Zhao, Y.H.; Hu, G.J.; Yue, G.Y.; Sheng, Y.; Wu, J.C.; Chen, J.; et al. Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland. Sci. Total Environ. 2018, 622–623, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Kang, J.; Yuan, Z.; Xu, Z.; Han, G. Responses of nutrient resorption to warming and nitrogen fertilization in contrasting wet and dry years in a desert grassland. Plant Soil 2018, 432, 65–73. [Google Scholar] [CrossRef]
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Hu, L.; Cai, Q. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Chen, L.L.; Deng, Q.; Yuan, Z.Y.; Mu, X.M.; Kallenbach, R.L. Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of China. Ecol. Eng. 2018, 120, 14–22. [Google Scholar] [CrossRef]
- Yang, Y.H.; Luo, Y.Q. Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Bai, Z.; Lv, C. Effects of vegetation on runoff and soil erosion on reclaimed land in an opencast coal-mine dump in a loess area. Catena 2015, 128, 44–53. [Google Scholar] [CrossRef]
- Rice, S.K.; Westerman, B.; Federici, R. Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine-oak ecosystem. Plant Ecol. 2004, 174, 97–107. [Google Scholar] [CrossRef]
- Maassen, S.; Wirth, S. Soil microbiological monitoring of a pine forest after partial thinning for stand regeneration with beech seedlings. Soil Sci. Plant Nutr. 2004, 50, 815–819. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Lado-Monserrat, L.; Lidón, A.; Bautista, I. Litterfall, litter decomposition and associated nutrient fluxes in Pinus halepensis: Influence of tree removal intensity in a Mediterranean forest. Eur. J. For. Res. 2015, 5, 833–844. [Google Scholar] [CrossRef]
- Thibodeau, L.; Raymond, P.; Camiré, C.; Munson, A.D. Impact of precommercial thinning in balsam fir stands on soil nitrogen dynamics, microbial biomass, decomposition, and foliar nutrition. Can. J. For. Res. 2000, 30, 229–238. [Google Scholar] [CrossRef]
- Bauhus, J.; Paré, D.; Coté, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Lemma, B.; Nilsson, I.; Kleja, D.B.; Olsson, M.; Knicker, H. Decomposition and substrate quality of leaf litters and fine roots from three exotic plantations and a native forest in the southwestern highlands of Ethiopia. Soil Biol. Biochem. 2007, 39, 2317–2328. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J.; Doughty, R. Changes of soil microbial and enzyme activities are linked to soil C, N and P stoichiometry in afforested ecosystems. For. Ecol. Manag. 2018, 427, 289–295. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Z.; Peng, D.; Zhu, H.; Yang, G.; Ha, V.N.; Zhong, Z. Response of soil microbial community dynamics to Robinia pseudoacacia L. afforestation in the loess plateau: A chronosequence approach. Plant Soil 2018, 423, 327–338. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Huang, C. Spatial variability of soil total nitrogen and soil total phosphorus under different land uses in a small watershed on the Loess Plateau, China. Geoderma 2009, 150, 141–149. [Google Scholar] [CrossRef]
- Lazzaro, L.; Mazza, G.; d’Errico, G.; Fabiani, A.; Giuliani, C.; Inghilesi, A.F.; Lagomarsino, A.; Landi, S.; Lastrucci, L.; Pastorelli, R.; et al. How ecosystems change following invasion by Robinia pseudoacacia: Insights from soil chemical properties and soil microbial, nematode, microarthropod and plant communities. Sci. Total Environ. 2018, 622–623, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Hammerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Pang, X.Y.; Hu, B.; Bao, W.K.; Tian, G.L. Does thinning- induced gap size result in altered soil microbial community in pine plantation in eastern Tibetan Plateau? Ecol. Evol. 2017, 7, 2986–2993. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, W.; Xu, M.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Radtke, A.; Ambraß, S.; Zerbe, S.; Tonon, G.; Fontana, V.; Ammer, C. Traditional coppice forest management drives the invasion of Ailanthus altissima and Robinia pseudoacacia into deciduous forests. For. Ecol. Manag. 2013, 291, 308–317. [Google Scholar] [CrossRef]
- Fichtner, A.; von Oheimb, G.; Härdtle, W.; Wilken, C.; Gutknecht, J.L.M. Effects of anthropogenic disturbances on soil microbial communities in oak forests persist for more than 100 years. Soil Biol. Biochem. 2014, 70, 79–87. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Exotic plant species alter the microbial community structure and function in the soil. Ecology 2002, 83, 3152–3166. [Google Scholar] [CrossRef]
- Bossio, D.A.; Fleck, J.A.; Scow, K.M.; Fujii, R. Alteration of soil microbial communities and water quality in restored wetlands. Soil Biol. Biochem. 2006, 38, 1223–1233. [Google Scholar] [CrossRef]
- Gavazov, K.; Ingrisch, J.; Hasibeder, R.; Mills, R.T.; Buttler, A.; Gleixner, G.; Pumpanen, J.; Bahn, M. Winter ecology of a subalpine grassland: Effects of snow removal on soil respiration, microbial structure and function. Sci. Total Environ. 2017, 590–591, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.T.; Zak, D.R.; White, D.C.; Peacock, A. Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci. Soc. Am. J. 2001, 65, 359–367. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Y.; Ren, T.; Tian, Z.; Wang, G.; He, X.; Tian, C. Short-term effect of tillage and crop rotation on microbial community structure and enzyme activities of a clay loam soil. Biol. Fertil. Soils 2014, 50, 1077–1085. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Frostergård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Litter Characteristics | Root Characteristics | |||||
|---|---|---|---|---|---|---|---|
| Biomass (t ha−1) | Element Content (g kg−1) | Element Content (g kg−1) | |||||
| C | N | P | C | N | P | ||
| Lar15 | 50.68 ± 6.78 a | 70.71 ± 1.48 ab | 6.57 ± 0.65 ab | 0.59 ± 0.04 a | 52.07 ± 2.11 ab | 3.05 ± 0.24 a | 0.33 ± 0.02 a |
| Lar24 | 58.74 ± 11.26 b | 66.92 ± 1.14 a | 5.88 ± 0.34 a | 0.51 ± 0.02 b | 48.91 ± 1.04 a | 3.48 ± 0.19 a | 0.42 ± 0.01 b |
| Lar40 | 56.93 ± 8.10 b | 68.34 ± 1.31 ab | 6.21 ± 1.03 ab | 0.62 ± 0.04 a | 57.46 ± 3.72 b | 4.11 ± 0.43 b | 0.51 ± 0.03 c |
| Lar50 | 66.21 ± 13.79 c | 72.48 ± 1.64 b | 6.92 ± 0.71 b | 0.66 ± 0.01 a | 63.77 ± 2.34 c | 4.27 ± 0.56 b | 0.56 ± 0.02 c |
| Sampling Time | Thinning Treatments | STC (g kg−1) | STN (g kg−1) | STP (g kg−1) | MBC (mg kg−1) | MBN (mg kg−1) | MBP (mg kg−1) | NO3−-N (mg kg−1) | NH4+-N (mg kg−1) | SAP (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| August 2016 | Lar15 | 16.41 ± 0.42 Aa | 1.42 ± 0.26 Aa | 0.28 ± 0.02 Aa | 371.37 ± 16.46 Aa | 92.49 ± 3.74 Aa | 20.64 ± 2.40 Aa | 6.77 ± 1.32 Aa | 3.00 ± 0.47 Aa | 2.74 ± 0.17 Ac |
| Lar24 | 33.04 ± 1.51 Ab | 2.84 ± 0.25 Ab | 0.39 ± 0.01 Ab | 460.62 ± 17.63 Ab | 106.93 ± 5.48 Ab | 23.41 ± 1.14 Aa | 9.37 ± 0.65 Ab | 4.31 ± 0.03 Ab | 2.62 ± 0.21 Ab | |
| Lar40 | 45.65 ± 0.89 Ac | 3.66 ± 0.08 Ac | 0.49 ± 0.00 Ac | 626.00 ± 6.55 Ac | 143.61 ± 10.07 Ac | 32.45 ± 0.64 Ab | 12.43 ± 4.13 Ac | 5.49 ± 0.50 Ac | 2.25 ± 0.57 Aab | |
| Lar50 | 52.71 ± 2.85 Ac | 4.35 ± 0.11 Ad | 0.57 ± 0.02 Ac | 783.37 ± 9.36 Ad | 175.53 ± 4.22 Ad | 37.24 ± 0.93 Ac | 11.89 ± 6.19 Ac | 7.30 ± 0.15 Ad | 1.84 ± 0.76 Aa | |
| August 2017 | Lar15 | 12.23 ± 1.12 Ba | 1.13 ± 0.10 Ba | 0.19 ± 0.02 Ba | 226.04 ± 67.86 Ba | 64.49 ± 11.17 Ba | 19.31 ± 2.77 Aa | 3.20 ± 1.14 Ba | 2.08 ± 0.27 Ba | 5.48 ± 0.50 Bb |
| Lar24 | 26.24 ± 1.36 Bb | 2.10 ± 0.18 Bb | 0.28 ± 0.03 Bb | 355.28 ± 12.85 Bb | 92.26 ± 10.43 Ab | 22.76 ± 1.08 Aa | 6.46 ± 3.72 Bb | 3.33 ± 0.28 Bb | 5.19 ± 0.21 Bb | |
| Lar40 | 37.76 ± 2.40 Bc | 2.77 ± 0.09 Bb | 0.39 ± 0.01 Bc | 620.67 ± 42.46 Ac | 142.27 ± 7.20 Ac | 32.07 ± 1.76 Ab | 6.36 ± 2.21 Bb | 4.78 ± 0.37 Bc | 3.87 ± 0.11 Ba | |
| Lar50 | 51.09 ± 3.98 Ad | 3.79 ± 0.09 Bc | 0.48 ± 0.02 Bd | 756.70 ± 21.82 Ac | 170.86 ± 11.15 Ad | 36.31 ± 1.02 Ab | 8.50 ± 3.13 Bc | 6.32 ± 0.34 Bd | 3.35 ± 0.06 Ba |
| Treatment Methods | Soil Stoichiometry | Microbial Biomass | Microbial Biomass Stoichiometry | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| STC:STN | STC:STP | STN:STP | Microbial Biomass C (mg kg−1) | Microbial Biomass N (mg kg−1) | Microbial Biomass P (mg kg−1) | MBC:MBN | MBC:MBP | MBN:MBP | ||
| 2016 | Lar15 | 10.60 ± 0.51 a | 60.98 ± 0.61 a | 5.77 ± 0.34 a | 371.37 ± 16.46 a | 92.49 ± 3.74 a | 20.64 ± 2.40 a | 4.01 ± 0.02 a | 18.21 ± 2.81 a | 4.53 ± 0.67 a |
| Lar24 | 11.71 ± 1.51 b | 84.44 ± 5.68 b | 7.25 ± 0.48 b | 460.62 ± 17.63 b | 106.93 ± 5.48 b | 23.41 ± 1.14 a | 4.32 ± 0.29 b | 19.69 ± 0.65 b | 4.57 ± 0.19 a | |
| Lar40 | 12.47 ± 0.50 c | 93.87 ± 1.61 c | 7.53 ± 0.22 c | 626.00 ± 6.55 c | 143.61 ± 10.07 c | 32.45 ± 0.64 b | 4.37 ± 0.32 b | 19.29 ± 0.39 b | 4.43 ± 0.39 a | |
| Lar50 | 12.12 ± 0.85 c | 92.43 ± 2.26 c | 7.64 ± 0.36 c | 783.37 ± 9.36 d | 175.53 ± 4.22 d | 37.24 ± 0.93 c | 4.47 ± 0.16 c | 21.05 ± 0.77 c | 4.71 ± 0.03 a | |
| 2017 | Lar15 | 11.84 ± 0.25 a | 69.13 ± 6.74 a | 5.85 ± 0.69 a | 226.04 ± 67.86 a | 64.49 ± 11.17 a | 19.31 ± 2.77 a | 3.51 ± 1.01 a | 11.65 ± 3.06 a | 3.33 ± 0.12 a |
| Lar24 | 12.53 ± 0.61 b | 93.40 ± 3.79 b | 7.46 ± 0.31 b | 355.28 ± 12.85 b | 92.26 ± 10.43 b | 22.76 ± 1.08 a | 3.87 ± 0.29 a | 15.62 ± 0.30 b | 4.05 ± 0.26 b | |
| Lar40 | 13.61 ± 0.68 c | 97.06 ± 6.18 b | 7.13 ± 0.36 b | 620.67 ± 42.46 c | 142.27 ± 7.20 c | 32.07 ± 1.76 b | 4.36 ± 0.14 b | 19.42 ± 2.07 c | 4.44 ± 0.06 bc | |
| Lar50 | 13.48 ± 0.98 c | 107.00 ± 9.02 c | 7.94 ± 0.16 c | 756.70 ± 21.82 d | 170.86 ± 11.15 d | 36.31 ± 1.02 c | 4.44 ± 0.36 b | 20.85 ± 0.71 c | 4.70 ± 0.24 c | |
| Predicted Variables | R2 | Sig. | Contribution of the Individual Predictor (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | Actinobacteria | Gram-Positive Bacteria | Gram-Negative Bacteria | Soil Temperature | SWC | STP | NH4+-N | |||
| Microbial C:N | 0.27 | <0.05 | 29.1 | 23.8 | 22.4 | 10.4 | 14.3 | |||
| Microbial C:P | 0.53 | <0.01 | 26.9 | 33.7 | 23.5 | 15.9 | ||||
| Microbial N:P | 0.20 | <0.05 | 34.6 | 13.9 | 14.9 | 17.3 | 19.3 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, M.; Cheng, X.; Liu, L.; Peng, X.; Shang, T.; Han, H. Soil Microbial Community and Soil Abiotic Factors Are Linked to Microorganisms’ C:N:P Stoichiometry in Larix Plantations. Forests 2023, 14, 1914. https://doi.org/10.3390/f14091914
Cai M, Cheng X, Liu L, Peng X, Shang T, Han H. Soil Microbial Community and Soil Abiotic Factors Are Linked to Microorganisms’ C:N:P Stoichiometry in Larix Plantations. Forests. 2023; 14(9):1914. https://doi.org/10.3390/f14091914
Chicago/Turabian StyleCai, Mengke, Xiaoqin Cheng, Li Liu, Xinhao Peng, Tianxiong Shang, and Hairong Han. 2023. "Soil Microbial Community and Soil Abiotic Factors Are Linked to Microorganisms’ C:N:P Stoichiometry in Larix Plantations" Forests 14, no. 9: 1914. https://doi.org/10.3390/f14091914