
How Forest Management with Clear-Cutting Affects the Regeneration, Diversity and Structure of a Seasonally Dry Tropical Forest in Brazil
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
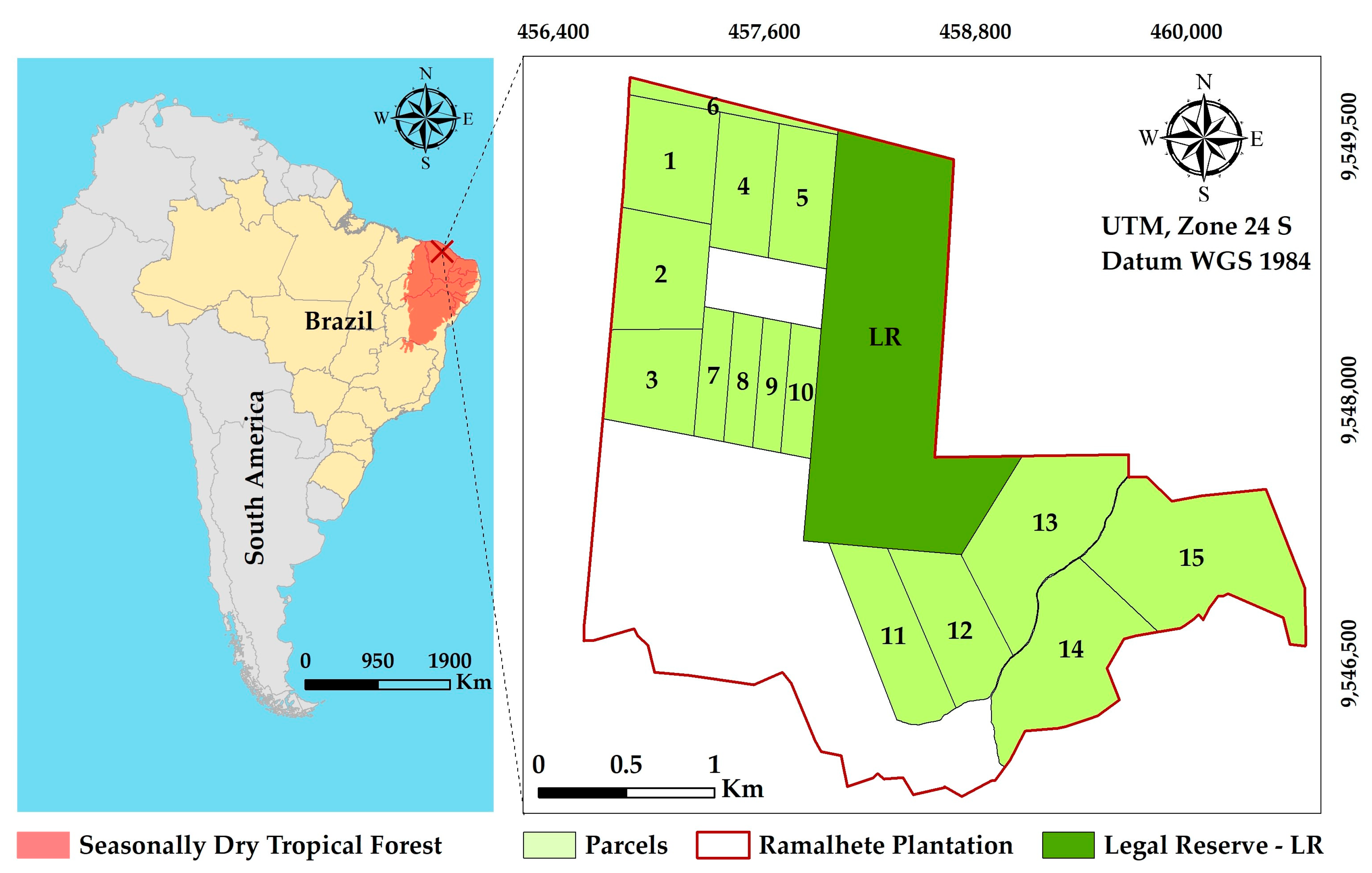
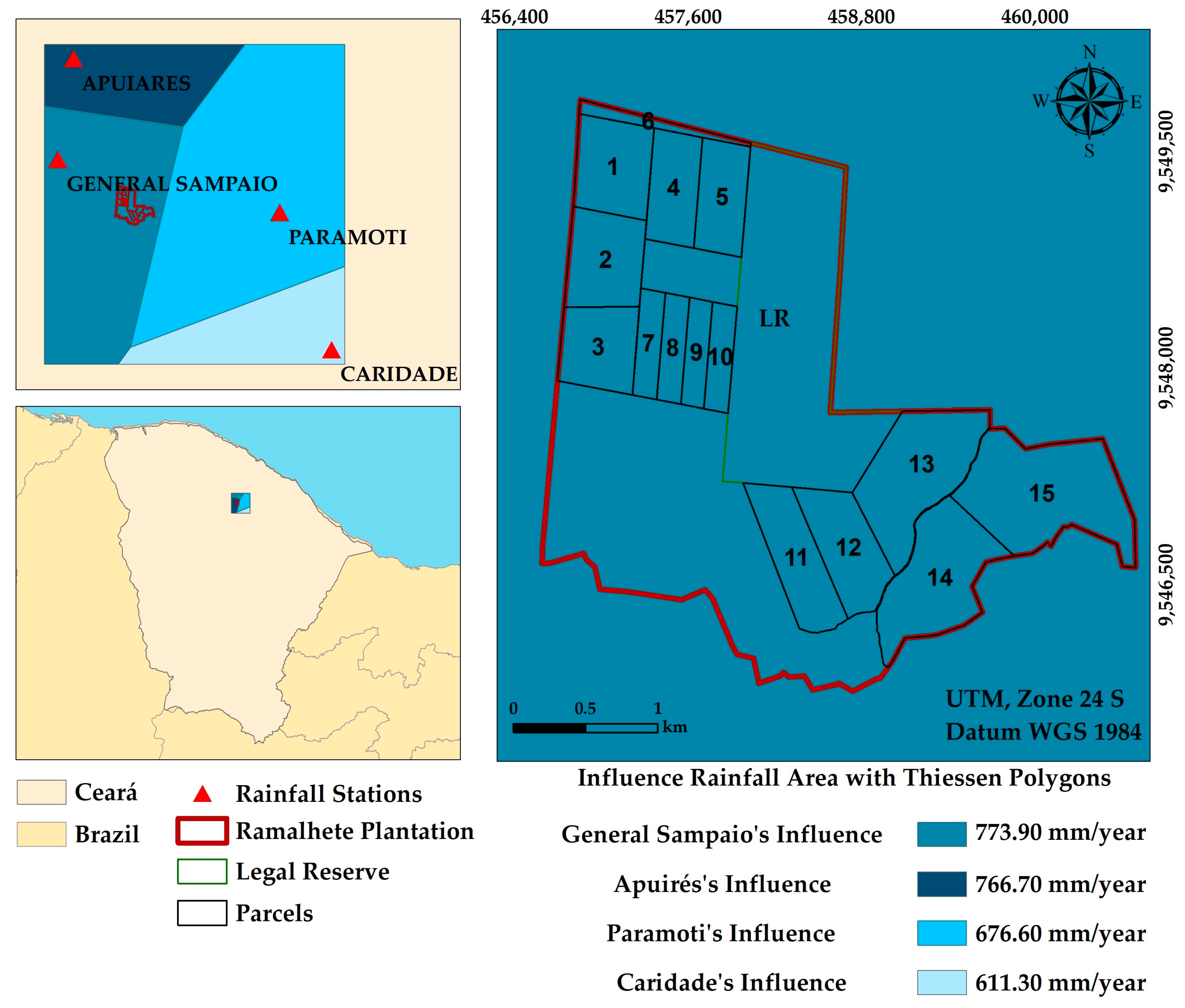
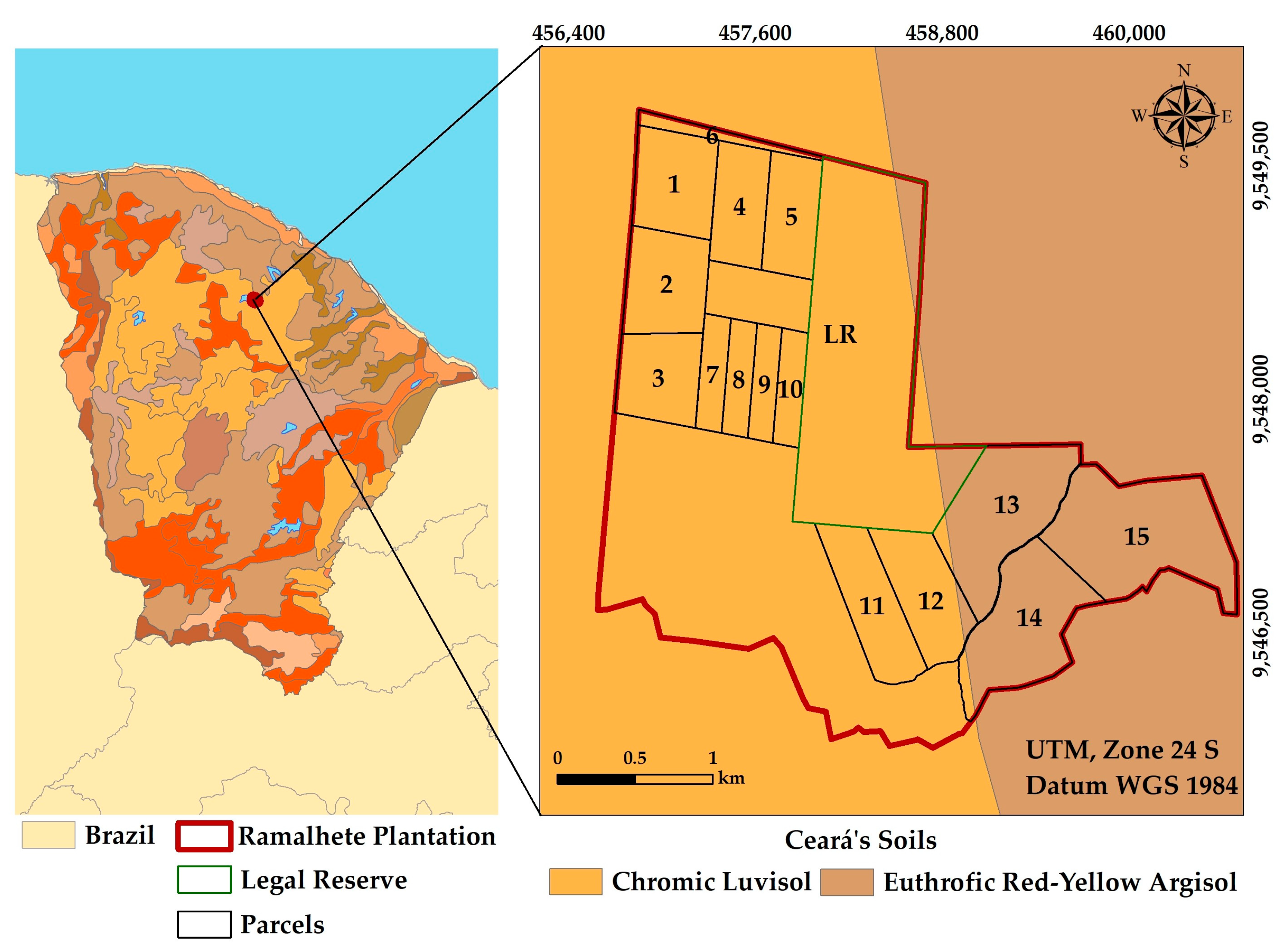
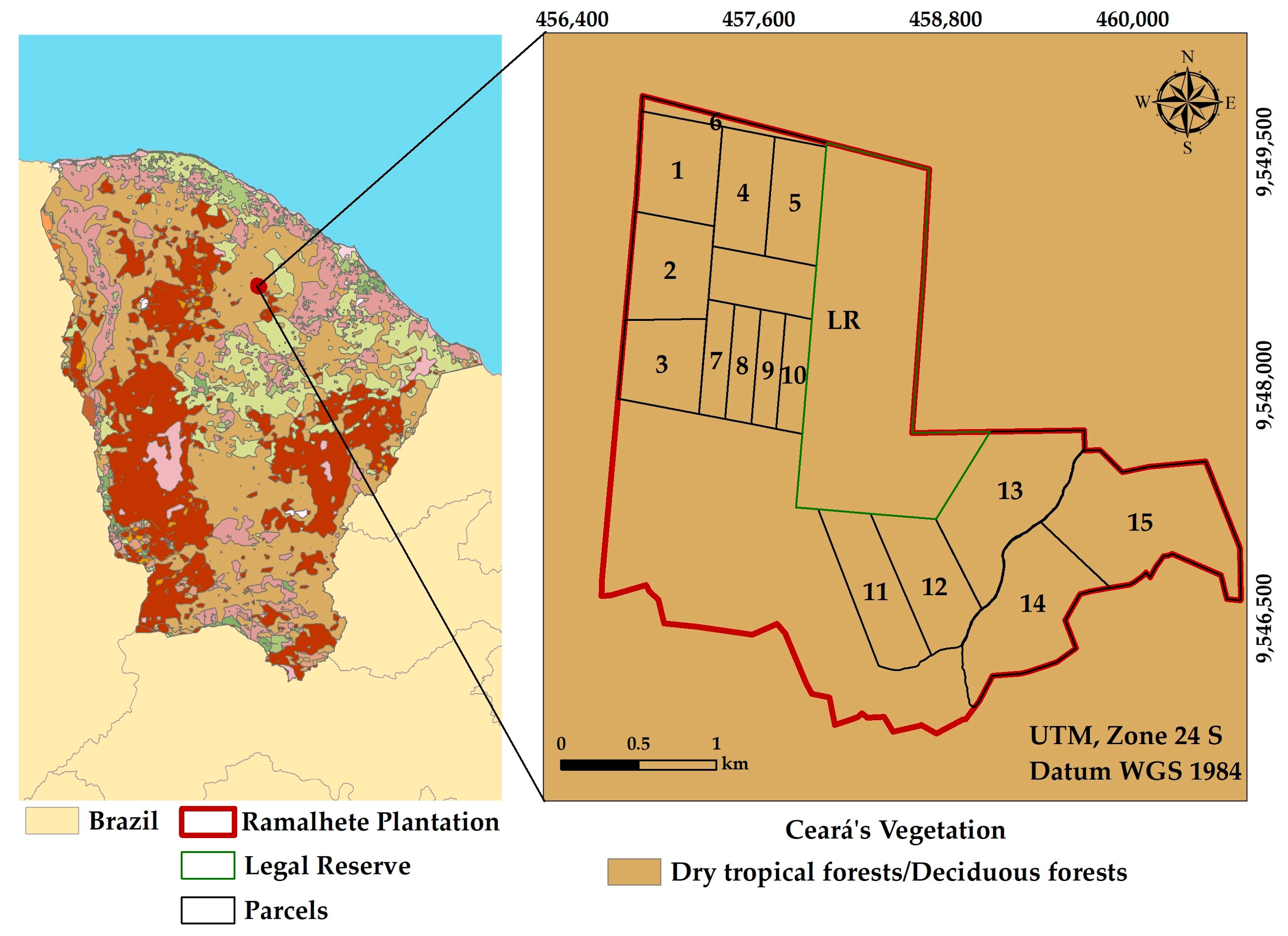
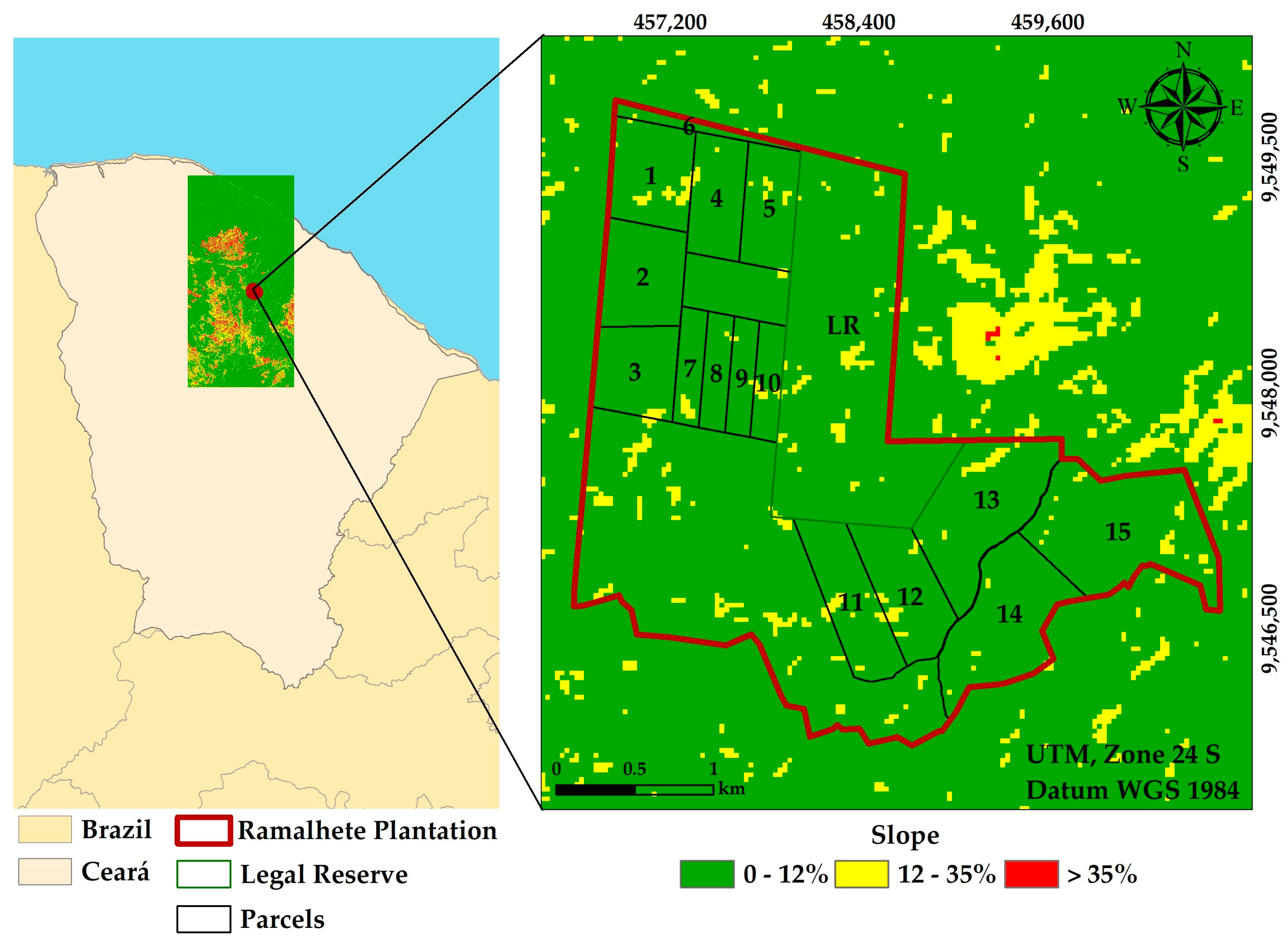
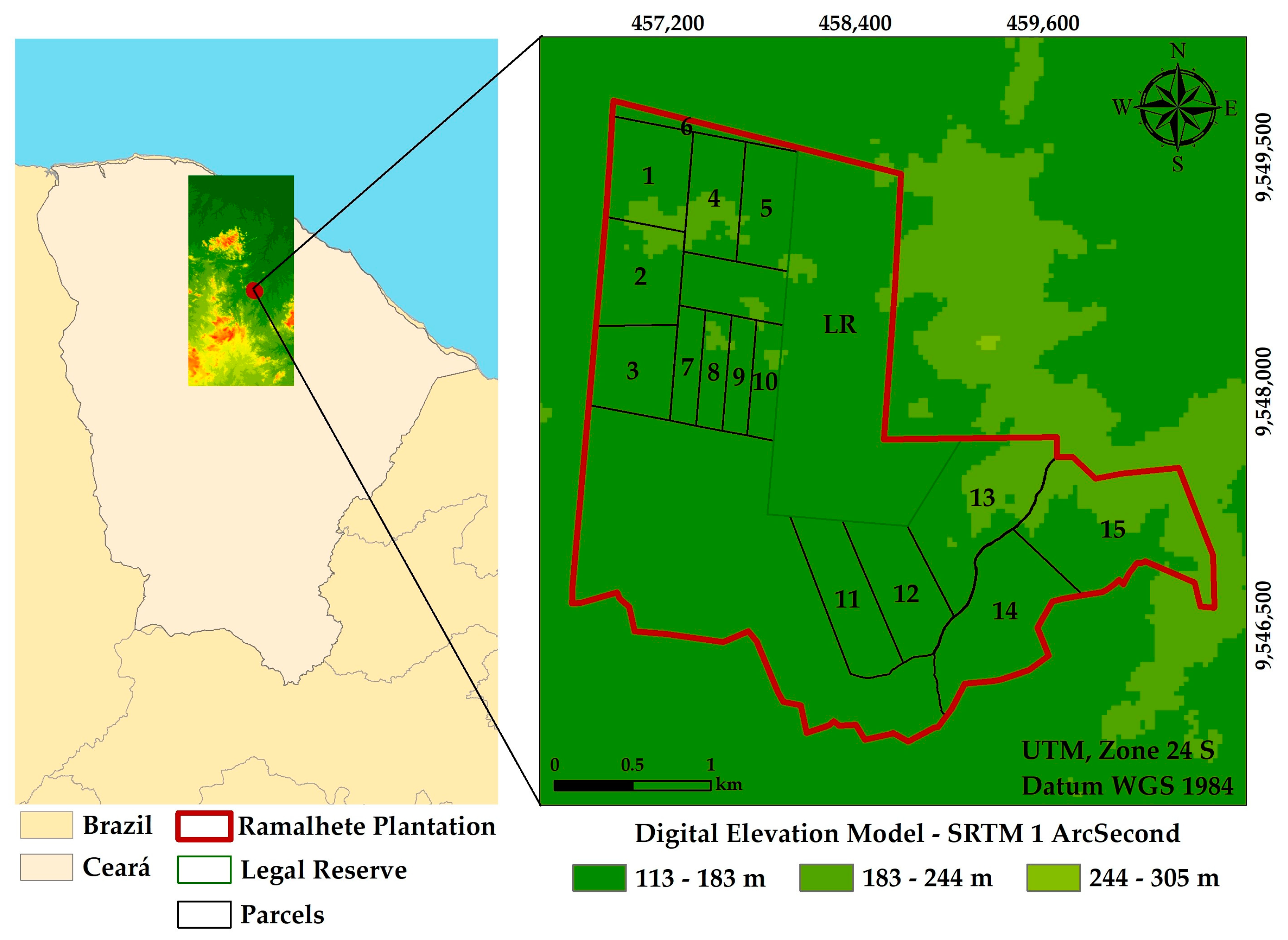
2.1. Study Area
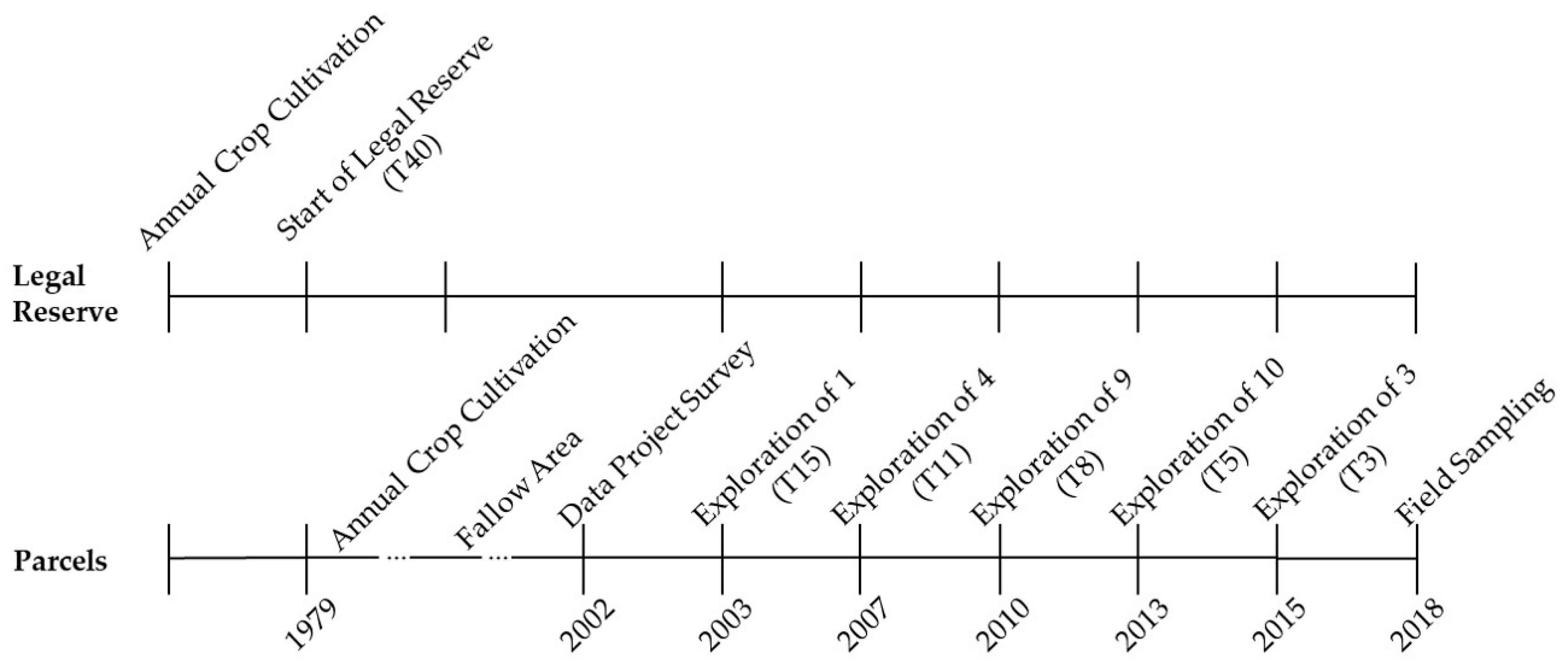
2.2. Sampling the Vegetation
2.3. Analysis
2.3.1. Species Diversity between Treatments
2.3.2. Differences between Treatments
3. Results
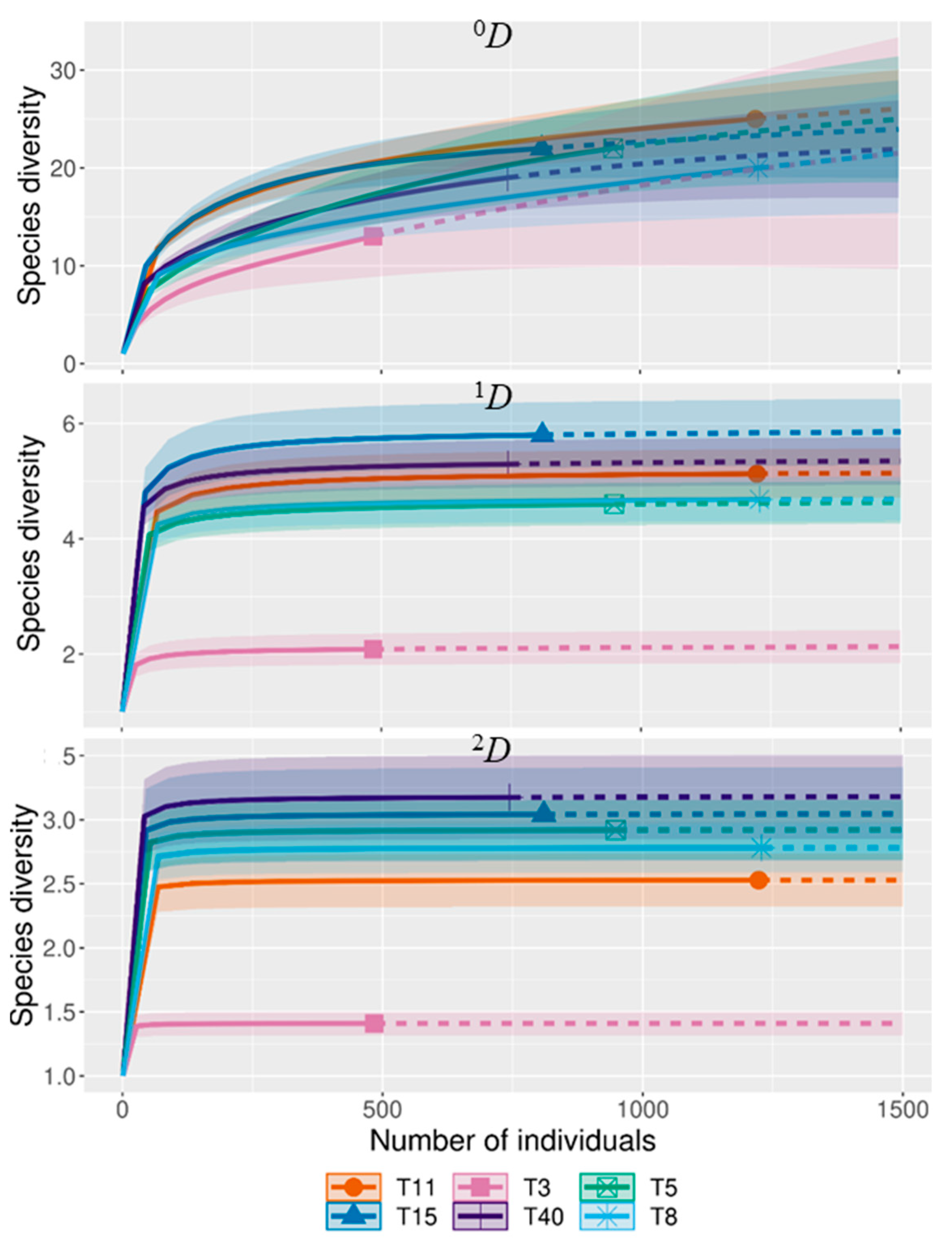
3.1. Species Composition and Diversity
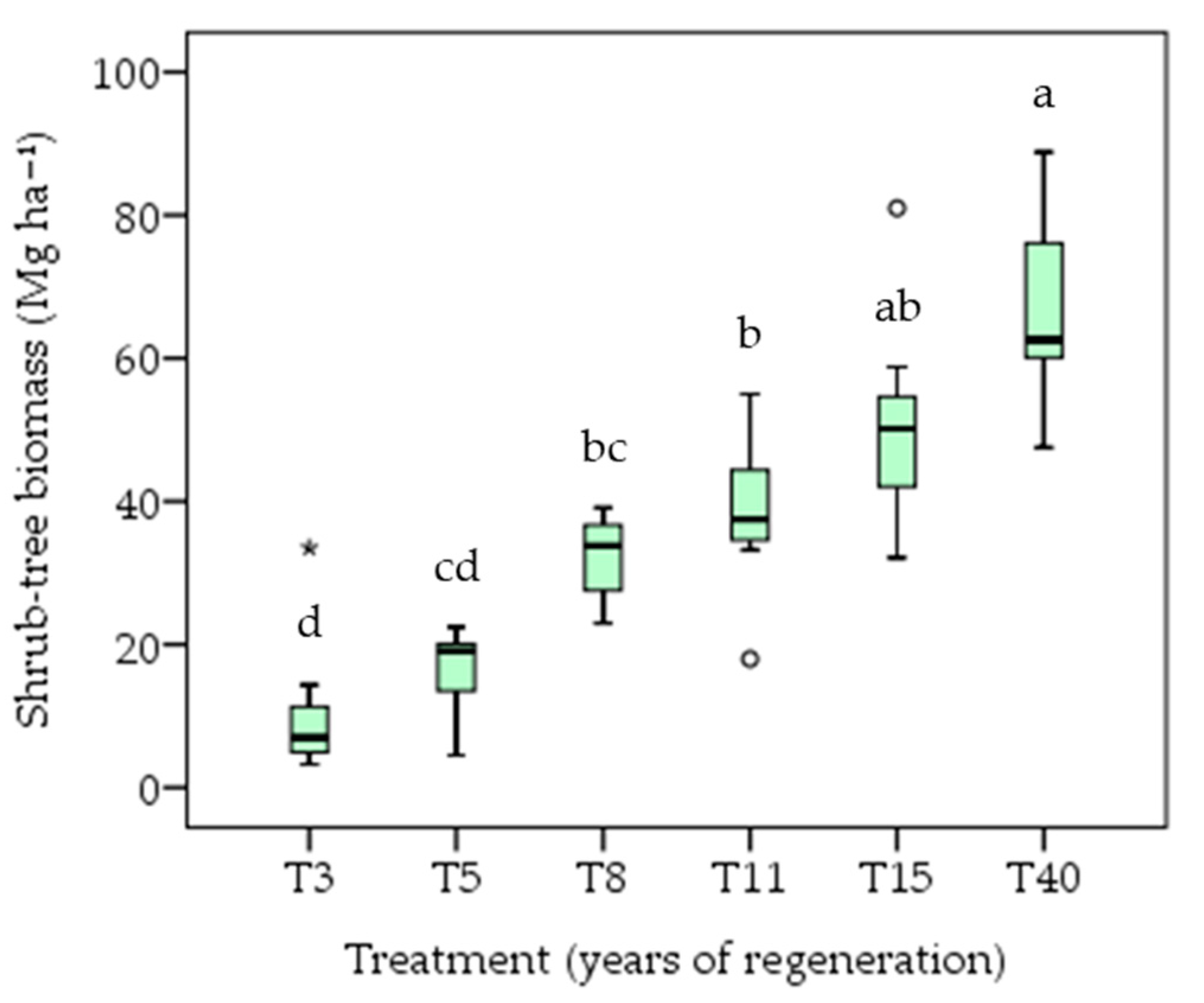
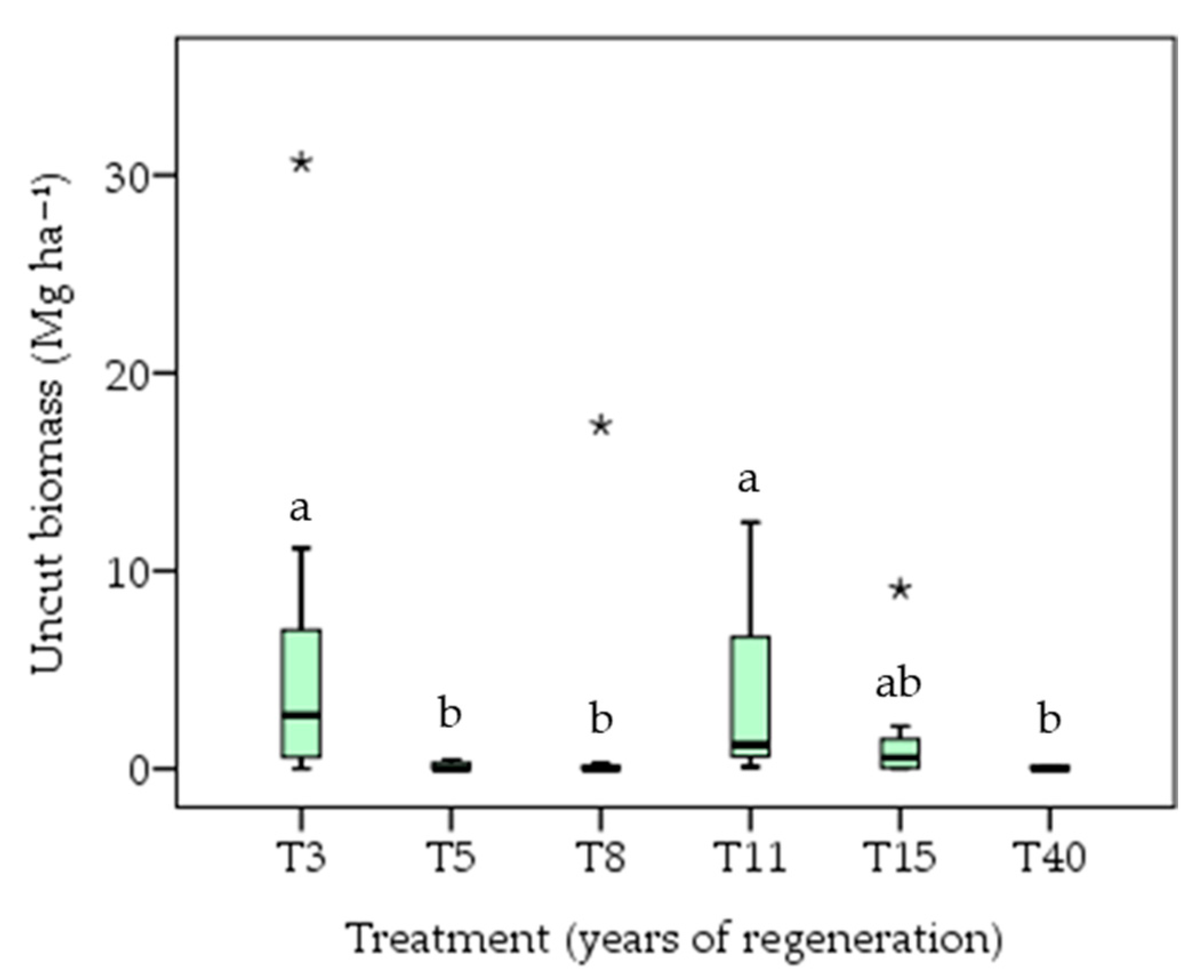
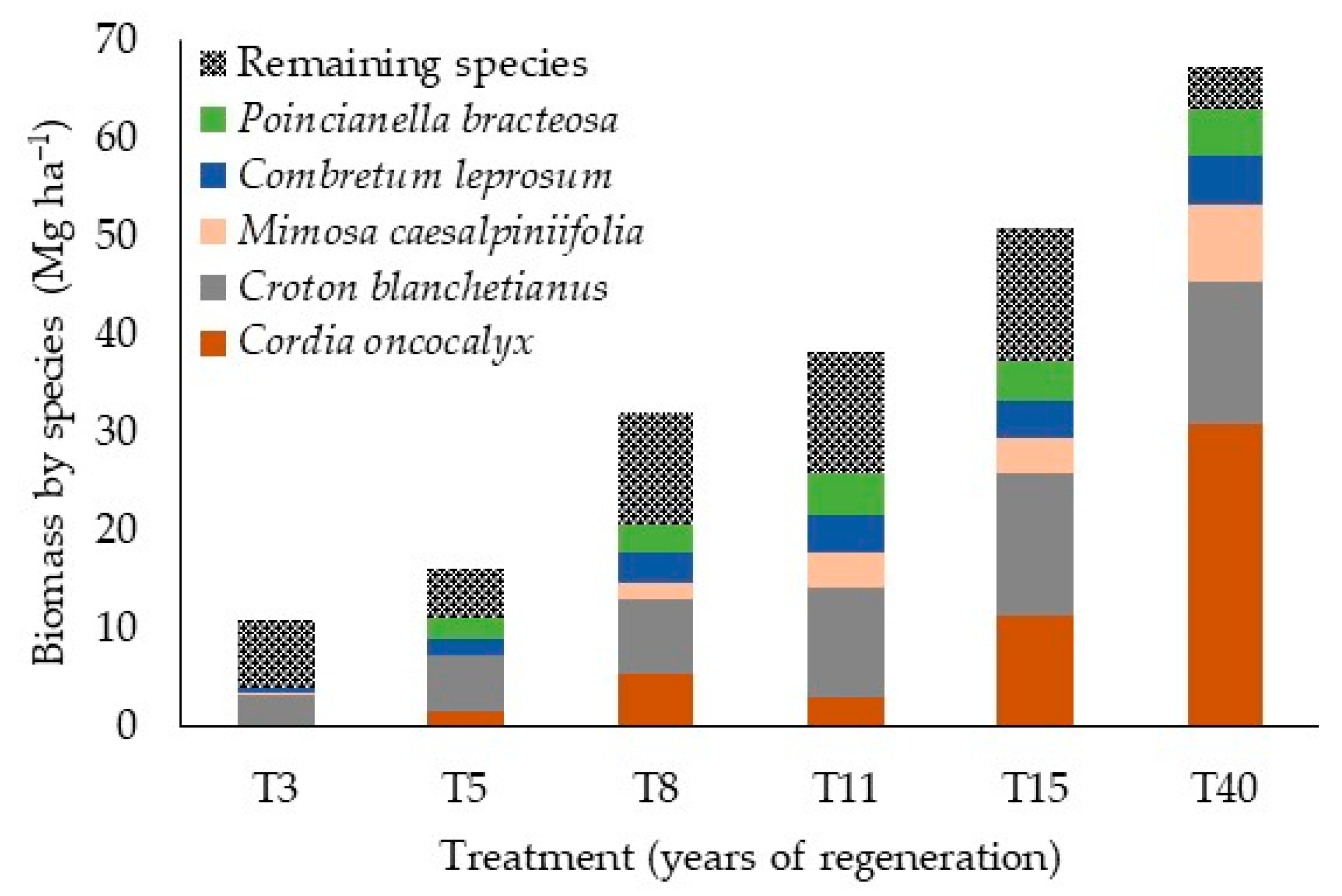
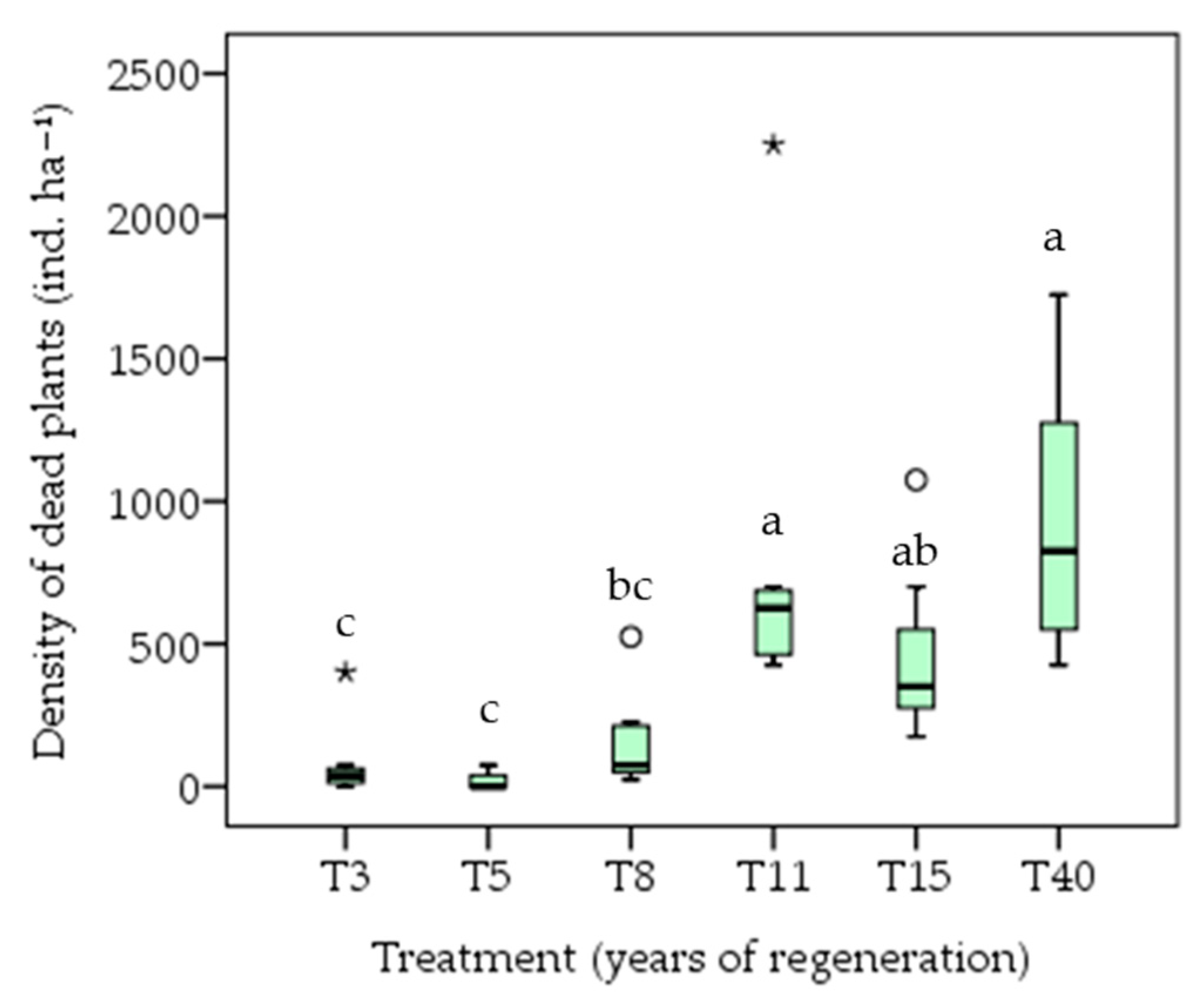
3.2. Biomass Accumulation
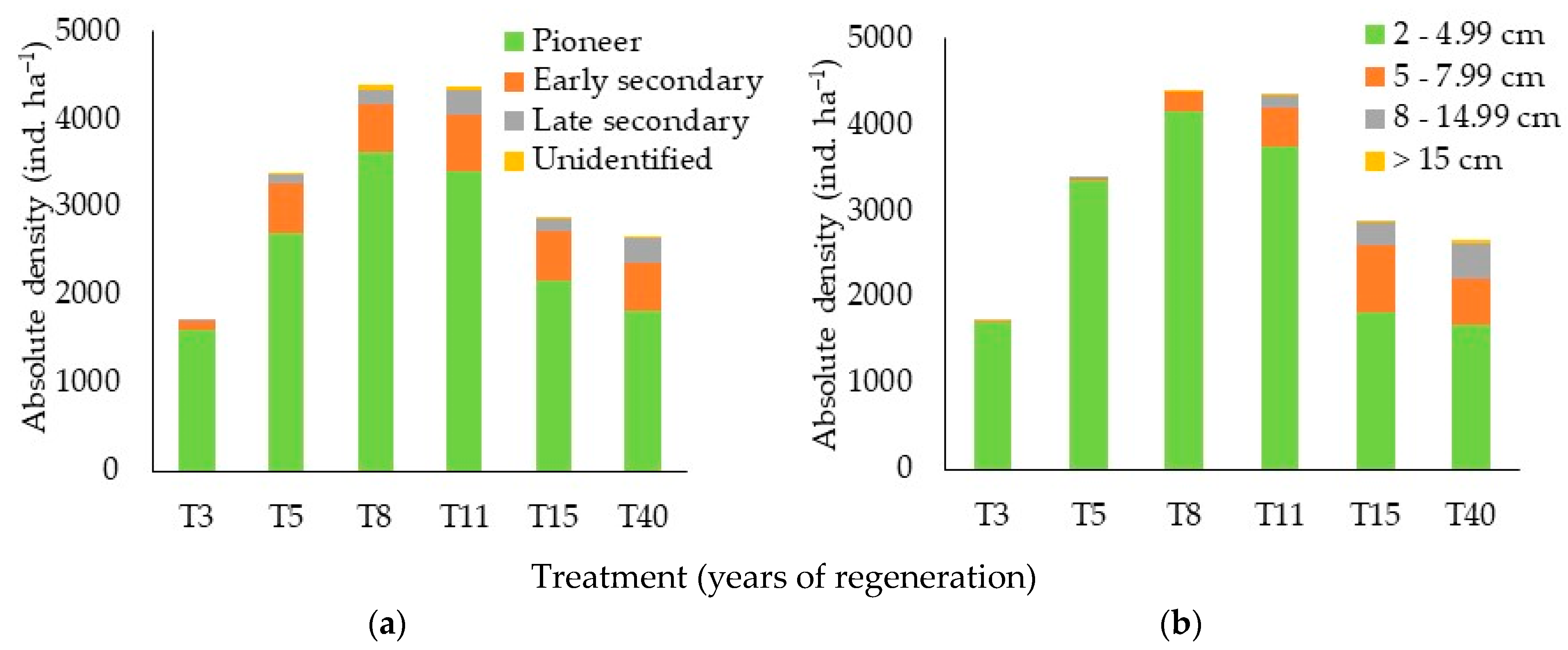
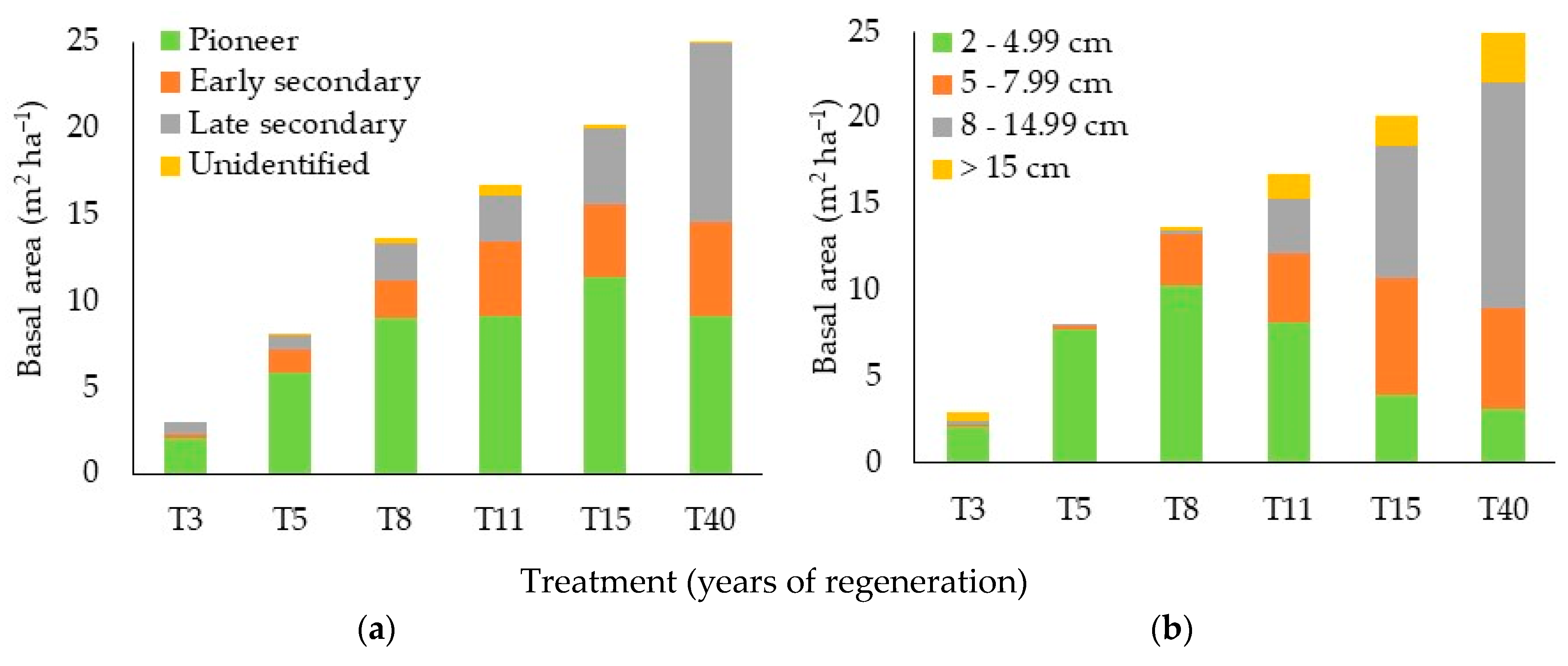
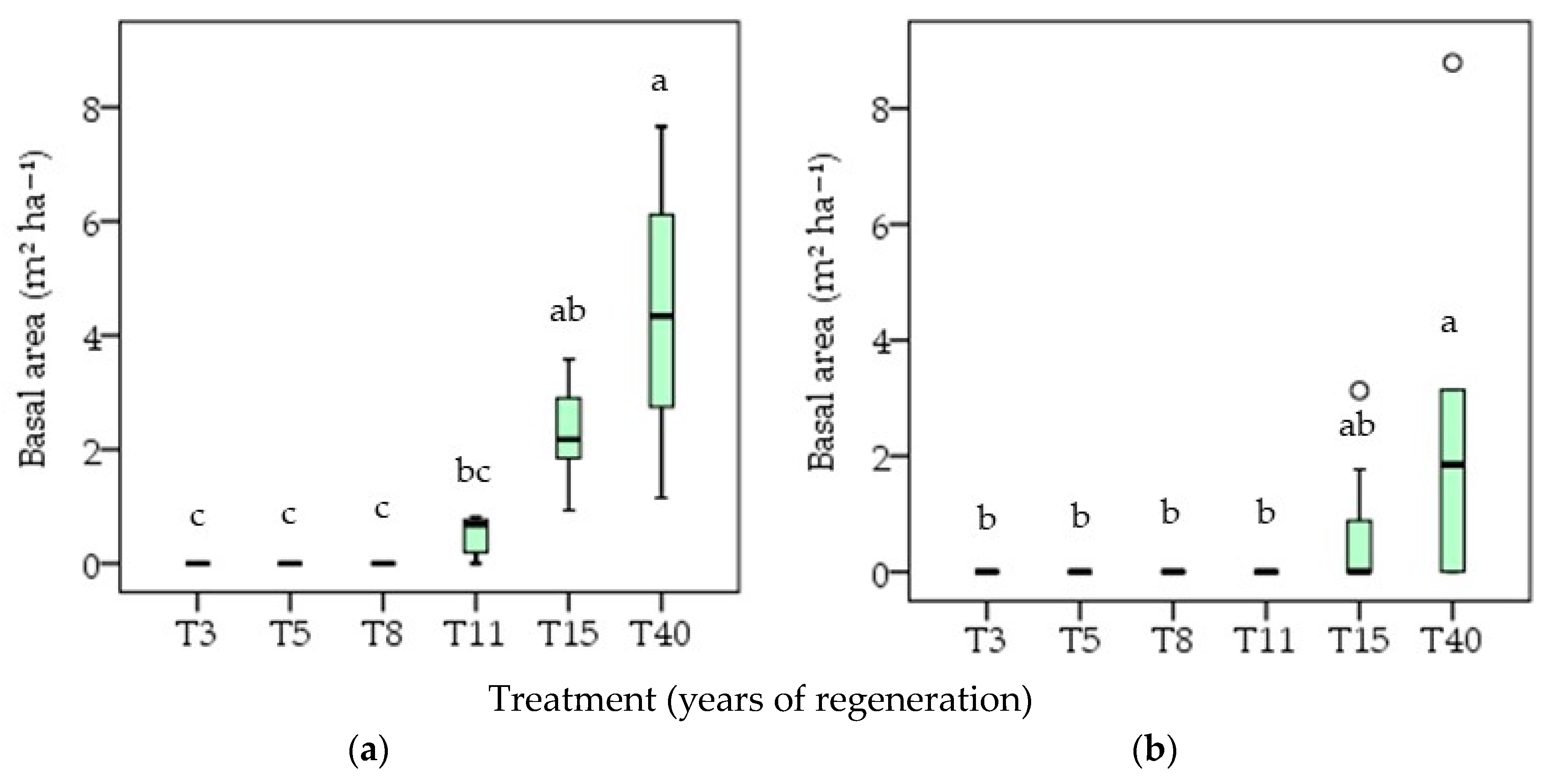
3.3. Community Structure
4. Discussion
4.1. Species Composition and Diversity
4.2. Biomass Accumulation
4.3. Community Structure
4.3.1. Density and Basal Area by Ecological Group and Diameter Class
4.3.2. Basal Area of Individuals with the Potential for Greater Added Value
4.3.3. Value Index by Species and Ecological Group
4.4. Effects of Management and Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Trees, Forests and Land Use in Drylands: The First Global Assessment—Full Report. FAO Forestry Paper No. 184. Rome. 2019. Available online: https://www.fs.usda.gov/rm/pubs_journals/2019/rmrs_2019_patriarca_c001.pdf (accessed on 20 June 2023).
- Silva, J.M.C.; Leal, I.R.; Tabarelli, M. Caatinga: The Largest Tropical Dry Forest Region in South America; Springer International Publishing: Cham, Switzerland, 2017; 482p. [Google Scholar] [CrossRef]
- Moro, M.F.; Nic Lughadha, E.; de Araújo, F.S.; Martins, F.R. A Phytogeographical Metaanalysis of the Semiarid Caatinga Domain in Brazil. Bot. Rev. 2016, 82, 91–148. [Google Scholar] [CrossRef]
- Souza, D.G.; Sfair, J.C.; de Paula, A.S.; Barros, M.F.; Rito, K.F.; Tabarelli, M. Multiple drivers of aboveground biomass in a human-modified landscape of the Caatinga dry forest. For. Ecol. Manag. 2018, 435, 57–65. [Google Scholar] [CrossRef]
- Barros, M.F.; Ribeiro, E.M.; Vanderlei, R.S.; de Paula, A.S.; Silva, A.B.; Wirth, R.; Cianciaruso, M.V.; Tabarelli, M. Resprouting drives successional pathways and the resilience of Caatinga dry forest in human-modified landscapes. For. Ecol. Manag. 2020, 482, 118881. [Google Scholar] [CrossRef]
- Arnan, X.; Leal, I.R.; Tabarelli, M.; Andrade, J.F.; Barros, M.F.; Câmara, T.; Jamelli, D.; Knoechelmann, C.M.; Menezes, T.G.; Menezes, A.G.; et al. A framework for deriving measures of chronic anthropogenic disturbance: Surrogate, direct, single and multi-metric indices in Brazilian Caatinga. Ecol. Indic. 2018, 94, 274–282. [Google Scholar] [CrossRef]
- APNE—Associação Plantas do Nordeste: Planos de Manejo Florestal Sustentado na Caatinga. Available online: http://www.cnip.org.br/planos_manejo.html (accessed on 20 June 2023).
- SEMACE—Superintendência Estadual do Meio Ambiente do Estado do Ceará. Relação de Planos de Manejo. Available online: https://www.semace.ce.gov.br/wp-content/uploads/sites/46/2019/11/PLANILHA-PMF-ATUALIZADA.pdf (accessed on 20 June 2023).
- Riegelhaupt, E.; Pareyn, F.G.C.; Gariglio, M.A. O manejo florestal como ferramenta para o uso sustentável e conservação da caatinga. In Uso Sustentável e Conservação dos Recursos Florestais da Caatinga; Gariglio, M.A., Sampaio, E.V.S.B., Cestaro, L.A., Kageyama, P.Y., Eds.; Serviço Florestal Brasileiro: Brasília, Brasil, 2010; pp. 349–367. ISBN 978-85-63269-04-1. [Google Scholar]
- Sist, P.; Mazzei, L.; Blanc, L.; Rutishauser, E. Large trees as key elements of carbon storage and dynamics after selective logging in the Eastern Amazon. For. Ecol. Manag. 2014, 318, 103–109. [Google Scholar] [CrossRef]
- Martínez-Meléndez, N.; Ramírez-Marcial, N.; Martínez-Zurimendi, P.; Cach-Pérez, M.J.; García-Franco, J.G. How Timber Harvest Affects the Structure and Diversity of a Montane Forest in Southern Mexico. Forests 2021, 12, 895. [Google Scholar] [CrossRef]
- Lopes, J.F.B.; Lopes, F.B.; Andrade, E.M.; Aquino, D.N. Composição florística com indutor da agregação de valor à biomassa florestal em vegetação caatinga. In Proceedings of the V Simpósio Brasileiro de Recursos Naturais do Semiárido-SBRNS, Fortaleza, Ceará, Brasil, 27 October 2021; Available online: https://drive.google.com/file/d/1Y2HHyi38nKBxVdbBR8WCw2kmNgqmWLNJ/view (accessed on 21 June 2023).
- Schröder, J.M.; Rodríguez, L.P.; Günter, S. Research trends: Tropical dry forests: The neglected research agenda? For. Policy Econ. 2020, 122, 102333. [Google Scholar] [CrossRef]
- Alcoforado-Filho, F.G.; Sampaio, E.V.d.S.B.; Rodal, M.J.N. Florística e fitossociologia de um remanescente de vegetação caducifólia espinhosa arbórea em Caruaru, Pernambuco. Acta Bot. Bras. 2003, 17, 287–303. [Google Scholar] [CrossRef]
- Sampaio, E.V.; Silva, G.C. Biomass equations for Brazilian semiarid caatinga plants. Acta Bot. Bras. 2005, 19, 935–943. [Google Scholar] [CrossRef]
- Araújo Filho, J.A. Manejo Pastoril Sustentável da Caatinga; Projeto Dom Helder Câmara: Recife, PE, Brasil, 2013; 200p, ISBN 978-85-64154-04-9. [Google Scholar]
- Andrade, E.M.; Aquino, D.N.; Chaves, L.C.G.; Lopes, F.B. Water as Capital and Its Uses in the Caatinga, In Caatinga: The Largest Tropical Dry Forest Region in South America, 1st ed.; Silva, J.M.C., Leal, I.R., Tabarelli, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 281–302. [Google Scholar] [CrossRef]
- Júnior, L.R.P.; de Andrade, E.M.; Palácio, H.A.d.Q.; Raymer, P.C.L.; Filho, J.C.R.; Pereira, F.J.S. Carbon stocks in a tropical dry forest in Brazil. Rev. Ciên. Agron. 2016, 47, 32–40. [Google Scholar] [CrossRef]
- Sabino, F.G.d.S.; Cunha, M.D.C.L.; Santana, G.M. Estrutura da Vegetação em Dois Fragmentos de Caatinga Antropizada na Paraíba. Flor. Amb. 2016, 23, 487–497. [Google Scholar] [CrossRef]
- de Souza, M.R.; Ferreira, M.B.; de Sousa, G.G.; Alves, A.R.; de Holanda, A.C. Caracterização florística e fitossociológica do componente lenhoso de um fragmento florestal de caatinga em serra do mel, rio grande do norte, brasil. Nativa 2020, 8, 329–335. [Google Scholar] [CrossRef]
- Santos, D.L.; Lopes, F.B.; Aquino, D.N. Análise da densidade de indivíduos em fragmentos da vegetação arbórea-arbustiva no bioma caatinga. In Proceedings of the V Simpósio Brasileiro de Recursos Naturais do Semiárido-SBRNS, Fortaleza, Ceará, Brasil, 27 October 2021; Available online: https://drive.google.com/file/d/16ybbmslYhDrT_cahyuOUTUMDcK5aq0sL/view (accessed on 23 June 2023).
- Maia, A.R.S.; Lopes, F.B.; de Andrade, E.M. Influence of Climatic Seasonality on a Survey of Land Use and Cover in the Semi-arid Region. J. Agric. Sci. 2018, 10, 311–325. [Google Scholar] [CrossRef]
- Costa, R.C.; Araújo, F.S. Physiognomy and structure of a caatinga with Cordia oncocalyx (Boraginaceae), a new type of community in Andrade-Lima’s classification of caatingas. Rodriguésia 2012, 63, 269–276. [Google Scholar] [CrossRef]
- Souza, E.B.; Nepomuceno, F.Á.A.; Santos, F.D.S.; Araújo, F.F.; Nepomuceno, I.V.; Paula, A.S.; Amorim, V.O.; Branco, M.S.D.; Rabelo, S.T.; Pinto, D.M.M.; et al. Flora and physiognomy of Caatinga vegetation over crystalline bedrock in the northern Caatinga domain, Brazil. Rodriguésia 2022, 73, e01252021. [Google Scholar] [CrossRef]
- Lopes, J.F.B.; De Andrade, E.M.; Pereira, E.C.B.; Campos, D.A.; Aquino, D.D.N. Cut cycles and soil carbon potential stocks in a managed forest in the caatinga domain in brazil. Rev. Caatin. 2020, 33, 735–745. [Google Scholar] [CrossRef]
- Campos, D.A.; Andrade, E.M.; Castanho, A.D.A.; Feitosa, R.C.; Palácio, H.Q.A. Biomass Dynamics in a Fragment of Brazilian Tropical Forest (Caatinga) over Consecutive Dry Years. Appl. Sci. 2020, 10, 7813. [Google Scholar] [CrossRef]
- Andrade, L.A.; Pereira, I.M.; Leite, U.T.; Barbosa, M.R.V. Análise da cobertura de duas fitofisionomias de caatinga, com diferentes históricos de uso, no município de São João do Cariri, Estado da Paraíba. Rev. Cerne 2005, 11, 253–262. Available online: https://cerne.ufla.br/site/index.php/CERNE/article/view/442/381 (accessed on 23 June 2023).
- Lopes, C.G.R.; Ferraz, E.M.N.; de Castro, C.C.; de Lima, E.N.; dos Santos, J.M.F.F.; dos Santos, D.M.; Araújo, E.d.L. Forest succession and distance from preserved patches in the Brazilian semiarid region. For. Ecol. Manag. 2012, 271, 115–123. [Google Scholar] [CrossRef]
- Cabral, G.A.L.; Sampaio, E.V.S.B.; Almeida-Cortez, J.S. Estrutura espacial e biomassa da parte aérea em diferentes estágios sucessionais de Caatinga, em Santa Terezinha, Paraíba. Rev. Bras. Geog. Fís. 2013, 6, 566–574. [Google Scholar] [CrossRef]
- Siyum, Z.G. Tropical dry forest dynamics in the context of climate change: Syntheses of drivers, gaps, and management perspectives. Ecol. Proc. 2020, 9, 1–16. [Google Scholar] [CrossRef]
- Aquino, D.d.N.; Neto, O.C.d.R.; Moreira, M.A.; Teixeira, A.d.S.; de Andrade, E.M. Use of remote sensing to identify areas at risk of degradation in the semi-arid region. Rev. Cienc. Agron. 2018, 49, 420–429. [Google Scholar] [CrossRef]
- Duarte, M.R.N.; Pereira, T.M.; Lima, P.F.; Pereira, E.C.B.; Lopes, F.B.; Rezende, C.F. Limnological dynamics in artificial reservoir and intermittent river in the semiarid region depending on land use and occupation. Rev. Cien. Agron. 2021, 51, e20207181. [Google Scholar] [CrossRef]
- Chaves, L.C.G.; Lopes, F.B.; Maia, A.R.S.; Meireles, A.C.M.; De Andrade, E.M. Water quality and anthropogenic impact in the watersheds of service reservoirs in the Brazilian semi-arid region. Rev. Cien. Agron. 2019, 50, 223–233. [Google Scholar] [CrossRef]
- Pereira, E.C.B.; Lopes, F.B.; Almeida, A.M.M.; Andrade, E.M.; Lopes, J.F.B. Sedimentos e nutrientes aportados a um reservatório de pequeno porte no Semiárido tropical. Rev. Cien. Agron. 2021, 52, e20196717. [Google Scholar] [CrossRef]
- Edwards, D.P.; Socolar, J.B.; Mills, S.C.; Burivalova, Z.; Koh, L.P.; Wilcove, D.S. Conservation of Tropical Forests in the Anthropocene. Cur. Biol. 2019, 29, R1008–R1020. [Google Scholar] [CrossRef]
- Carlson, K.M.; Heilmayr, R.; Gibbs, H.K.; Noojipady, P.; Burns, D.N.; Morton, D.C.; Walker, N.F.; Paoli, G.D.; Kremen, C. Effect of oil palm sustainability certification on deforestation and fire in Indonesia. Proc. Natl. Acad. Sci. USA 2018, 115, 121–126. [Google Scholar] [CrossRef]
- WWF—World Wildlife Fund. Forests the Size of France Grow Again in the Last 20 Years—New Study. Available online: https://www.wwf.org.uk/press-release/wwf-forest-regeneration-study (accessed on 16 August 2023).
- Nanni, A.S.; Sloan, S.; Aide, T.M.; Graesser, J.; Edwards, D.; Grau, H.R. The neotropical reforestation hotspots: A biophysical and socioeconomic typology of contemporary forest expansion. Glob. Environ. Chang. 2019, 54, 148–159. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.C.; Pickens, A.; Hernandez-Serna, A.; Tyukavina, A.; Turubanova, S.; Zalles, V.; Li, X.; Khan, A.; Stolle, F.; et al. The Global 2000–2020 Land Cover and Land Use Change Dataset Derived from the Landsat Archive: First Results. Front. Remo. Sens. 2022, 3, 856903. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Chazdon, R.L. Beyond hectares: Four principles to guide reforestation in the context of tropical forest and landscape restoration. Restor. Ecol. 2017, 25, 491–496. [Google Scholar] [CrossRef]
- Ribeiro, E.M.S.; Arroyo-Rodríguez, V.; Santos, B.A.; Tabarelli, M.; Leal, I.R. Chronic anthropogenic disturbance drives the biological impoverishment of the Brazilian Caatinga vegetation. J. Appl. Ecol. 2015, 52, 611–620. [Google Scholar] [CrossRef]
- Ribeiro, E.M.S.; Santos, B.A.; Arroyo-Rodríguez, V.; Tabarelli, M.; Souza, G.; Leal, I.R. Phylogenetic impoverishment of plant communities following chronic human disturbances in the Brazilian Caatinga. Ecology 2016, 97, 1583–1592. [Google Scholar] [CrossRef]
- Montfort, F.; Nourtier, M.; Grinand, C.; Maneau, S.; Mercier, C.; Roelens, J.; Blanc, L. Regeneration capacities of woody species biodiversity and soil properties in Miombo woodland after slash-and-burn agriculture in Mozambique. For. Ecol. Manag. 2021, 488, 119039. [Google Scholar] [CrossRef]
- Tallei, E.; Rivera, L.; Schaaf, A.; Vivanco, C.; Politi, N. Post-logging changes in a neotropical dry forest composition and structure modify the ecosystem functioning. For. Ecol. Manag. 2023, 537, 120944. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Melo, F.P.L.; Martínez-Ramos, M.; Bongers, F.; Chazdon, R.L.; Meave, J.A.; Norden, N.; Santos, B.A.; Leal, I.R.; Tabarelli, M. Multiple successional pathways in human-modified tropical landscapes: New insights from forest succession, forest fragmentation and landscape ecology research. Biol. Rev. 2015, 92, 326–340. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L.; Guariguata, M.R. Natural regeneration as a tool for large-scale forest restoration in the tropics: Prospects and challenges. Biotropica 2016, 48, 716–730. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Lu, X.; Huang, J. The impacts of selective logging and clear-cutting on woody plant diversity after 40 years of natural recovery in a tropical montane rain forest, south China. Sci. Total Environ. 2017, 579, 1683–1691. [Google Scholar] [CrossRef]
- FUNCEME. Fundação Cearense de Meteorologia e Recursos Hídricos. Índices de Aridez do Estado do Ceará. Available online: http://www.funceme.br/?page_id=5826 (accessed on 11 August 2023).
- Andrade, E.M.; Sena, M.G.T.; Silva, A.G.R.; Pereira, F.J.S.; Lopes, F.B. Uncertainties of the rainfall regime in a tropical semi-arid region: The case of the State of Ceará. Rev. Agro@mbiente Online 2016, 10, 88–95. [Google Scholar] [CrossRef]
- VERDE VIDA. Reformulação de Plano de Manejo Agrossilvipastoril de Ramalhete; Verde Vida Engenharia Ambiental: General Sampaio, CE, Brasil, 2010; 121p. [Google Scholar]
- Santos, H.G.; Carvalho Junior, W.; Dart, R.O.; Aglio, M.L.D.; Sousa, J.S.; Pares, J.G.; Fontana, A.; Martins, A.L.S.; Oliveira, A.P. O Novo Mapa de Solos do Brasil: Legenda Atualizada, 1st ed.; Embrapa Solos: Rio de Janeiro, Brasil, 2011; 67p, ISSN 1517-2627. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/123772/1/DOC-130-O-novo-mapa-de-solos-do-Brasil.pdf (accessed on 16 August 2023).
- EMBRAPA. Embrapa Monitoramento por Satélite. Mapa de Cobertura Vegetal do Brasil. Available online: http://geoinfo.cnpm.embrapa.br/ (accessed on 10 August 2023).
- NASA JPL. NASA Shuttle Radar Topography Mission Global 1 Arc Second [Data Set]. NASA EOSDIS Land Processes Distributed Active Archive Center. 2013. Available online: https://lpdaac.usgs.gov/products/srtmgl1v003/ (accessed on 10 August 2023).
- Lopes, J.F.B. Dinâmica da Biomassa Aérea e Remanescente em área de Plano de Manejo na Floresta Tropical Seca, Caatinga. Mestrado; Universidade Federal do Ceará: Fortaleza, CE, Brasil, 2018; Available online: https://repositorio.ufc.br/handle/riufc/37030 (accessed on 10 August 2023).
- Jesus, K.N.; Albuquerque, E.R.G.M.; Pareyn, F.G.C.; Menezes, R.S.C.; Sampaio, E.V.S.B.; Costa, T.L.; Althoff, T.D. Guia Metodológico Para o Desenvolvimento de Atividades de Campo e Laboratório do Projeto CNPQ; Rede—C—NE: Recife, PE, Brasil, 2016; 35p. [Google Scholar]
- Rodal, M.J.N.F.; Sampaio, E.V.S.B.; Figueiredo, M.A. Manual Sobre Métodos de Estudos Florísticos e Fitossociológicos—Ecossistema Caatinga, 1st ed.; Sociedade Botânica do Brasil: Brasília, Brasil, 2013; 24p, ISBN 978-85-60428-03-8. Available online: https://www.botanica.org.br/wp-content/uploads/man_sob_met_est_flo_fit.pdf (accessed on 26 June 2023).
- Curtis, J.T.; Mcintosh, R.P. The interrelations of certain analytic and synthetic phytosociological characters. Ecology 1950, 31, 434–455. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monog. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Meth. Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Banda, T.; Mwangulango, N.; Meyer, B.; Schwartz, M.W.; Mbago, F.; Sungula, M.; Caro, T. The woodland vegetation of the Katavi-Rukwa ecosystem in western Tanzania. For. Ecol. Manag. 2008, 255, 3382–3395. [Google Scholar] [CrossRef]
- Sahu, S.C.; Suresh, H.S.; Ravindranath, N.H. Forest Structure, Composition and Above Ground Biomass of Tree Community in Tropical Dry Forests of Eastern Ghats, India. Not. Sci. Biol. 2016, 8, 125–133. [Google Scholar] [CrossRef]
- Rito, K.F.; Arroyo-Rodríguez, V.; Queiroz, R.T.; Leal, I.R.; Tabarelli, M. Precipitation mediates the effect of human disturbance on the Brazilian Caatinga vegetation. J. Ecol. 2017, 105, 828–838. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F.; Aide, T.M.; Zambrano, A.M.A.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.S.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass resilience of Neotropical secondary forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef]
- Feitosa, R.C. Estoque de Carbono em Floresta Tropical Sazonalmente Seca no Nordeste do Brasil: Uma Comparação Entre dois usos do solo. Doutorado; Universidade Federal do Ceará: Fortaleza, CE, Brasil, 2017; Available online: https://repositorio.ufc.br/handle/riufc/28686 (accessed on 3 July 2023).
- Sampaio, E.V.S.B. Caracterização do bioma caatinga—características e potencialidades. In Uso Sustentável e Conservação dos Recursos Florestais da Caatinga; Gariglio, M.A., Sampaio, E.V.S.B., Cestaro, L.A., Kageyama, P.Y., Eds.; Serviço Florestal Brasileiro: Brasília, Brasil, 2010; pp. 29–42. ISBN 978-85-63269-04-1. [Google Scholar]
- Sampaio, E.V.S.B.; Araújo, E.L.; Salcedo, I.H.; Tiessen, H. Regeneração da vegetação de caatinga após corte e queima, em Serra Talhada, PE. Pesq. Agrop. Bras. 1998, 33, 621–632. Available online: https://seer.sct.embrapa.br/index.php/pab/article/view/4890 (accessed on 26 June 2023).
- Kiill, L.H.P. (Coord.). Plantas da Caatinga Ameaçadas de Extinção: Estudos Preliminares e Manejo; Embrapa Semi-Árido: Petrolina, PE, Brasil, 2008; 87p, Available online: https://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/160098 (accessed on 27 June 2023).
- Kammesheidt, L. Forest recovery by root suckers and above-ground sprouts after slash-and-burn agriculture, fire and logging in Paraguay and Venezuela. J. Trop. Ecol. 1999, 15, 143–157. [Google Scholar] [CrossRef]
- Stoner, K.E.; Sánchez-Azofeifa, G.A. Ecology and regeneration of tropical dry forests in the Americas: Implications for management. For. Ecol. Manag. 2009, 258, 903–906. [Google Scholar] [CrossRef]
- Moro, M.F.; Araújo, F.S.; Rodal, M.J.N.; Martins, F.R. Síntese dos estudos florísticos e fitossociológicos realizados no semiárido brasileiro. In Fitossociologia No Brasil, 1st ed.; Eisenlohr, P.V., Felfili, J.M., Melo, M.M.R.F., Andrade, L.A., Meira Neto, J.A.A., Eds.; Editora da Universidade Federal de Viçosa: Viçosa, MG, Brasil, 2015; Volume 2, pp. 412–451. [Google Scholar]
- Yan, P.; Xu, L.; He, N. Variation in the calorific values of different plants organs in China. PLoS ONE 2018, 13, e0199762. [Google Scholar] [CrossRef]
- Zeng, W.S.; Tang, S.Z.; Xiao, Q.H. Calorific values and ash contents of different parts of Masson pine trees in southern China. J. For. Res. 2014, 25, 779–786. [Google Scholar] [CrossRef]
- Pereira Filho, J.M.; Bakke, O.A. Produção de forragem de espécies herbáceas da caatinga. In Uso Sustentável e Conservação dos Recursos Florestais da Caatinga; Gariglio, M.A., Sampaio, E.V.S.B., Cestaro, L.A., Kageyama, P.Y., Eds.; Serviço Florestal Brasileiro: Brasília, Brasil, 2010; pp. 145–159. ISBN 978-85-63269-04-1. [Google Scholar]
- Brito, L.B.M.; Araújo, F.S. Banco de sementes de Cordia oncocalyx Allemão em uma área de Caatinga sobre planossolo. Rev. Caat. 2009, 22, 206–212. Available online: https://periodicos.ufersa.edu.br/caatinga/article/view/321/631 (accessed on 27 June 2023).
- Andrade-Lima, D. The caatinga dominium. Rev. Bras. Bot. 1981, 4, 149–153. [Google Scholar]
- Muniz, C.R.; Correia, D.; Soares, A.A. Method of opening the fruits, biometry and anatomical description of embryonic development in pau-branco (Cordia oncocalyx Allemão) seeds. Ciên. Flor. 2022, 32, 979–995. [Google Scholar] [CrossRef]
- Aguiar, M.I.; Fialho, J.S.; Campanha, M.M.; Oliveira, T.S. Florística e estrutura vegetal em áreas de Caatinga sob diferentes sistemas de manejo. Pesq. Flor. Bra. 2019, 39, 1–11. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Accumulated Number and Percentage of Species of Each Treatment | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T3 | % | T5 | % | T8 | % | T11 | % | T15 | % | T40 | % | Total | |
| Species | 13 | 38 | 22 | 65 | 20 | 59 | 25 | 74 | 23 | 68 | 19 | 56 | 34 |
| Ecological Group | |||||||||||||
| Pioneer | 7 | 54 | 8 | 36 | 7 | 35 | 7 | 28 | 8 | 35 | 6 | 32 | 10 |
| Early secondary | 4 | 31 | 7 | 32 | 6 | 30 | 8 | 32 | 8 | 35 | 7 | 37 | 9 |
| Late secondary | 2 | 15 | 5 | 23 | 4 | 20 | 6 | 24 | 4 | 17 | 5 | 26 | 8 |
| Undetermined | 0 | 0 | 2 | 9 | 3 | 15 | 4 | 16 | 3 | 13 | 1 | 5 | 7 |
| Total | 13 | 100 | 22 | 100 | 20 | 100 | 25 | 100 | 23 | 100 | 19 | 100 | 34 |
| Species | Biomass of Each Treatment (Mg ha−1) | |||||
|---|---|---|---|---|---|---|
| T3 | T5 | T8 | T11 | T15 | T40 | |
| Poincianella bracteosa * | 0.05 ± 0.03 b | 2.29 ± 0.58 a | 3.04 ± 0.86 a | 4.50 ± 1.88 a | 4.10 ± 1.21 a | 4.66 ± 3.19 a |
| Croton blanchetianus * | 3.28 ± 0.83 d | 5.74 ± 0.80 cd | 7.69 ± 1.07 bcd | 11.38 ± 2.16 abc | 14.76 ± 1.66 a | 14.60 ± 4.29 ab |
| Combretum leprosum * | 0.35 ± 0.08 a | 1.54 ± 0.33 a | 3.01 ± 1.37 a | 3.83 ± 1.64 a | 3.69 ± 1.25 a | 4.91 ± 2.50 a |
| Cordia oncocalyx * | 0.00 b | 1.57 ± 0.45 a | 5.37 ± 2.06 a | 2.96 ± 1.17 a | 11.25 ± 4.64 a | 30.88 ± 11.28 a |
| Mimosa caesalpiniifolia * | 0.27 ± 0.21 b | 0.04 ± 0.03 b | 1.63 ± 0.80 a | 3.35 ± 1.17 a | 3.53 ± 1.50 a | 7.94 ± 3.30 a |
| Remaining species 1 | 6.93 ± 4.21 | 4.98 ± 1.12 | 11.32 ± 2.09 | 12.31 ± 3.48 | 13.62 ± 2.63 | 4.33 ± 2.43 |
| Variable | Density and Basal Area of Trees of Each Treatment | |||||
|---|---|---|---|---|---|---|
| T3 | T5 | T8 | T11 | T15 | T40 | |
| Density (individuals ha−1) * | 1729 ± 265 c | 3386 ± 392 ab | 4386 ± 216 a | 4368 ± 516 a | 2893 ± 242 abc | 2657 ± 441 bc |
| Ecological Group | ||||||
| Pioneer * | 1604 ± 281 b | 2704 ± 307 ab | 3625 ± 122 a | 3407 ± 537 a | 2168 ± 293 ab | 1821 ± 417 b |
| Early secondary ** | 107 ± 15 b | 564 ± 177 a | 546 ± 156 a | 650 ± 131 a | 557 ± 147 a | 543 ± 99 a |
| Late secondary ** | 18 ± 7 c | 100 ± 26 bc | 164 ± 59 ab | 275 ± 31 a | 139 ± 22 ab | 289 ± 135 ab |
| Undetermined | 0 | 18 ± 9 | 50 ± 29 | 36 ± 24 | 29 ± 16 | 4 ± 4 |
| Diameter class | ||||||
| 2–4.99 cm ** | 1693 ± 270 c | 3357 ± 386 ab | 4161 ± 209 a | 3750 ± 537 a | 1814 ± 232 bc | 1689 ± 407 c |
| 5–7.99 cm ** | 14 ± 9 d | 25 ± 13 cd | 214 ± 45 bc | 457 ± 121 ab | 779 ± 68 a | 525 ± 80 ab |
| 8–14.99 cm ** | 14 ± 5 bc | 4 ± 4 c | 7 ± 7 c | 139 ± 57 ab | 279 ± 44 a | 414 ± 68 a |
| >15 cm ** | 7 ± 5 abc | 0 c | 4 ± 4 bc | 21 ± 10 ab | 21 ± 9 ab | 29 ± 10 a |
| Basal área (m2 ha−1) * | 3.0 ± 0.6 e | 8.0 ± 1.1 de | 13.7 ± 1.1 cd | 16.7 ± 1.7 bc | 20.2 ± 1.8 ab | 25.0 ± 1.6 a |
| Ecological Group | ||||||
| Pioneer ** | 2.1 ± 0.5 c | 5.8 ± 0.8 bc | 9.0 ± 0.3 ab | 9.2 ± 1.1 ab | 11.4 ± 0.7 a | 9.2 ± 2.5 ab |
| Early secondary ** | 0.2 ± 0.0 c | 1.3 ± 0.3 bc | 2.2 ± 0.7 b | 4.2 ± 1.3 ab | 4.1 ± 0.9 ab | 5.4 ± 0.9 a |
| Late secondary ** | 0.7 ± 0.4 c | 0.8 ± 0.2 bc | 2.1 ± 0.8 ab | 2.7 ± 0.7 ab | 4.4 ± 1.5 ab | 10.3 ± 3.8 a |
| Undetermined | 0.0 | 0.0 | 0.4 ± 0.3 | 0.7 ± 0.5 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| Diameter class | ||||||
| 2–4.99 cm ** | 2.2 ± 0.5 d | 7.8 ± 1.1 ab | 10.3 ± 0.9 a | 8.2 ± 1.3 ab | 4.0 ± 0.3 bcd | 3.2 ± 0.7 cd |
| 5–7.99 cm ** | 0.1 ± 0.1 b | 0.2 ± 0.1 b | 2.9 ± 0.8 a | 4.0 ± 0.9 a | 6.8 ± 0.8 a | 5.8 ± 1.1 a |
| 8–14.99 cm ** | 0.1 ± 0.1 c | 0.1 ± 0.1 c | 0.2 ± 0.2 c | 3.2 ± 1.1 a | 7.7 ± 1.9 a | 13.1 ± 2.3 a |
| >15 cm ** | 0.6 ± 0.4 bc | 0.0 c | 0.2 ± 0.2 bc | 1.4 ± 0.6 ab | 1.7 ± 0.9 ab | 2.9 ± 1.2 a |
| Treatment | EG Pioneer | IVI | EG Early Secondary | IVI | EG Late Secondary | IVI | Undetermined | IVI |
|---|---|---|---|---|---|---|---|---|
| T3 | Croton blanchetianus | 54.07 | Combretum leprosum | 9.77 | Amburana cearensis | 9.86 | ||
| Mimosa arenosa | 8.52 | Mimosa caesalpiniifolia | 2.08 | Myracrodruon urundeuva | 1.34 | |||
| Poincianella bracteosa | 4.98 | Manihot glaziovii | 1.01 | |||||
| Jatropha mollissima | 4.36 | Bauhinia cheliantha | 0.95 | |||||
| Croton anisodontus | 1.17 | |||||||
| Piptadenia stipulacea | 0.96 | |||||||
| Mimosa tenuiflora | 0.95 | |||||||
| Total | 75.01 | 13.80 | 11.20 | 0.00 | ||||
| T5 | Croton blanchetianus | 34.55 | Combretum leprosum | 9.51 | Cordia oncocalyx | 6.52 | Unknown 1 | 1.75 |
| Mimosa arenosa | 15.83 | Cordia thichotoma | 6.90 | Libidibia ferrea | 1.23 | Chloroleucon dumosum | 0.67 | |
| Poincianella bracteosa | 10.88 | Bauhinia cheliantha | 2.60 | Pisonia tomentosa | 0.88 | |||
| Mimosa tenuiflora | 0.61 | Anadenanthera colubrina | 1.29 | Handroanthus impetiginosus | 0.83 | |||
| Croton anisodontus | 0.60 | Ziziphus joazeiro | 1.25 | Pseudobombax marginatum | 0.57 | |||
| Cochlospermum vitifolium | 0.59 | Mimosa caesalpiniifolia | 1.22 | |||||
| Piptadenia stipulacea | 0.57 | Aspidosperma pyrifolium | 0.60 | |||||
| Piptadenia viridiflora | 0.56 | |||||||
| Total | 64.17 | 23.37 | 10.03 | 2.43 | ||||
| T8 | Croton blanchetianus | 32.92 | Combretum leprosum | 7.82 | Cordia oncocalyx | 9.39 | Unknown 3 | 2.26 |
| Mimosa arenosa | 13.39 | Mimosa caesalpiniifolia | 7.70 | Amburana cearensis | 0.65 | Acacia glomerosa | 0.58 | |
| Poincianella bracteosa | 9.35 | Bauhinia cheliantha | 1.86 | Libidibia ferrea | 0.64 | Unknown 2 | 0.58 | |
| Mimosa tenuiflora | 6.48 | Capparis cynophallophora | 0.58 | Pisonia tomentosa | 0.58 | |||
| Piptadenia stipulacea | 2.56 | Cordia thichotoma | 0.57 | |||||
| Piptadenia viridiflora | 0.89 | Anadenanthera colubrina | 0.57 | |||||
| Croton anisodontus | 0.64 | |||||||
| Total | 66.23 | 19.09 | 11.27 | 3.41 | ||||
| T11 | Croton blanchetianus | 34.77 | Mimosa caesalpiniifolia | 7.15 | Cordia oncocalyx | 5.88 | Ximenia americana | 2.21 |
| Poincianella bracteosa | 7.98 | Combretum leprosum | 6.89 | Amburana cearensis | 2.94 | Chloroleucon dumosum | 0.43 | |
| Mimosa arenosa | 4.52 | Piptadenia stipulacea | 4.46 | Myracrodruon urundeuva | 2.88 | Crataeva tapia | 0.41 | |
| Croton anisodontus | 2.56 | Bauhinia cheliantha | 3.40 | Commiphora leptophloeos | 2.31 | Unknown 2 | 0.41 | |
| Mimosa tenuiflora | 1.93 | Cordia thichotoma | 2.60 | Libidibia ferrea | 1.80 | |||
| Aspidosperma pyrifolium | 0.87 | Anadenanthera colubrina | 1.19 | Pisonia tomentosa | 0.60 | |||
| Jatropha mollissima | 0.41 | Manihot glaziovii | 0.98 | |||||
| Ziziphus joazeiro | 0.41 | |||||||
| Total | 53.04 | 27.08 | 16.42 | 3.46 | ||||
| T15 | Croton blanchetianus | 32.16 | Combretum leprosum | 7.95 | Cordia oncocalyx | 9.98 | Unknown 2 | 1.06 |
| Poincianella bracteosa | 9.08 | Mimosa caesalpiniifolia | 5.97 | Libidibia ferrea | 2.06 | Unknown 1 | 0.65 | |
| Mimosa tenuiflora | 7.57 | Bauhinia cheliantha | 5.25 | Pisonia tomentosa | 1.93 | Acacia glomerosa | 0.08 | |
| Mimosa arenosa | 3.42 | Anadenanthera colubrina | 2.52 | Myracrodruon urundeuva | 1.59 | |||
| Piptadenia stipulacea | 2.09 | Cordia thichotoma | 1.79 | |||||
| Cochlospermum vitifolium | 1.22 | Aspidosperma pyrifolium | 1.35 | |||||
| Croton anisodontus | 0.86 | Manihot glaziovii | 0.49 | |||||
| Croton conduplicatus | 0.46 | Capparis cynophallophora | 0.48 | |||||
| Total | 56.85 | 25.80 | 15.56 | 1.79 | ||||
| T40 | Croton blanchetianus | 30.36 | Mimosa caesalpiniifolia | 9.59 | Cordia oncocalyx | 20.48 | Ximenia americana | 0.78 |
| Poincianella bracteosa | 9.77 | Combretum leprosum | 8.66 | Pisonia tomentosa | 2.15 | |||
| Mimosa arenosa | 3.87 | Cordia thichotoma | 2.78 | Myracrodruon urundeuva | 1.35 | |||
| Mimosa tenuiflora | 3.61 | Aspidosperma pyrifolium | 1.33 | Commiphora leptophloeos | 0.71 | |||
| Cochlospermum vitifolium | 1.00 | Manihot glaziovii | 0.81 | Libidibia ferrea | 0.68 | |||
| Piptadenia stipulacea | 0.68 | Bauhinia cheliantha | 0.72 | |||||
| Anadenanthera colubrina | 0.67 | |||||||
| Total | 49.29 | 24.56 | 25.37 | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, J.F.B.; Lopes, F.B.; Araújo, I.C.d.S.; Pereira, E.C.B.; Brandão, M.L.S.M.; Feitosa, E.d.O.; Luna, N.R.d.S.; Sousa, G.G.d.; Amorim, A.V.; Iwata, B.d.F.; et al. How Forest Management with Clear-Cutting Affects the Regeneration, Diversity and Structure of a Seasonally Dry Tropical Forest in Brazil. Forests 2023, 14, 1870. https://doi.org/10.3390/f14091870
Lopes JFB, Lopes FB, Araújo ICdS, Pereira ECB, Brandão MLSM, Feitosa EdO, Luna NRdS, Sousa GGd, Amorim AV, Iwata BdF, et al. How Forest Management with Clear-Cutting Affects the Regeneration, Diversity and Structure of a Seasonally Dry Tropical Forest in Brazil. Forests. 2023; 14(9):1870. https://doi.org/10.3390/f14091870
Chicago/Turabian StyleLopes, José Frédson Bezerra, Fernando Bezerra Lopes, Isabel Cristina da Silva Araújo, Erich Celestino Braga Pereira, Maria Letícia Stefany Monteiro Brandão, Erialdo de Oliveira Feitosa, Nayara Rochelli de Sousa Luna, Geocleber Gomes de Sousa, Aiala Vieira Amorim, Bruna de Freitas Iwata, and et al. 2023. "How Forest Management with Clear-Cutting Affects the Regeneration, Diversity and Structure of a Seasonally Dry Tropical Forest in Brazil" Forests 14, no. 9: 1870. https://doi.org/10.3390/f14091870