
Characteristics and Relationships between Species Diversity and Productivity of Different Grassland Types in the Burqin Forest Region of the Altai Mountains
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
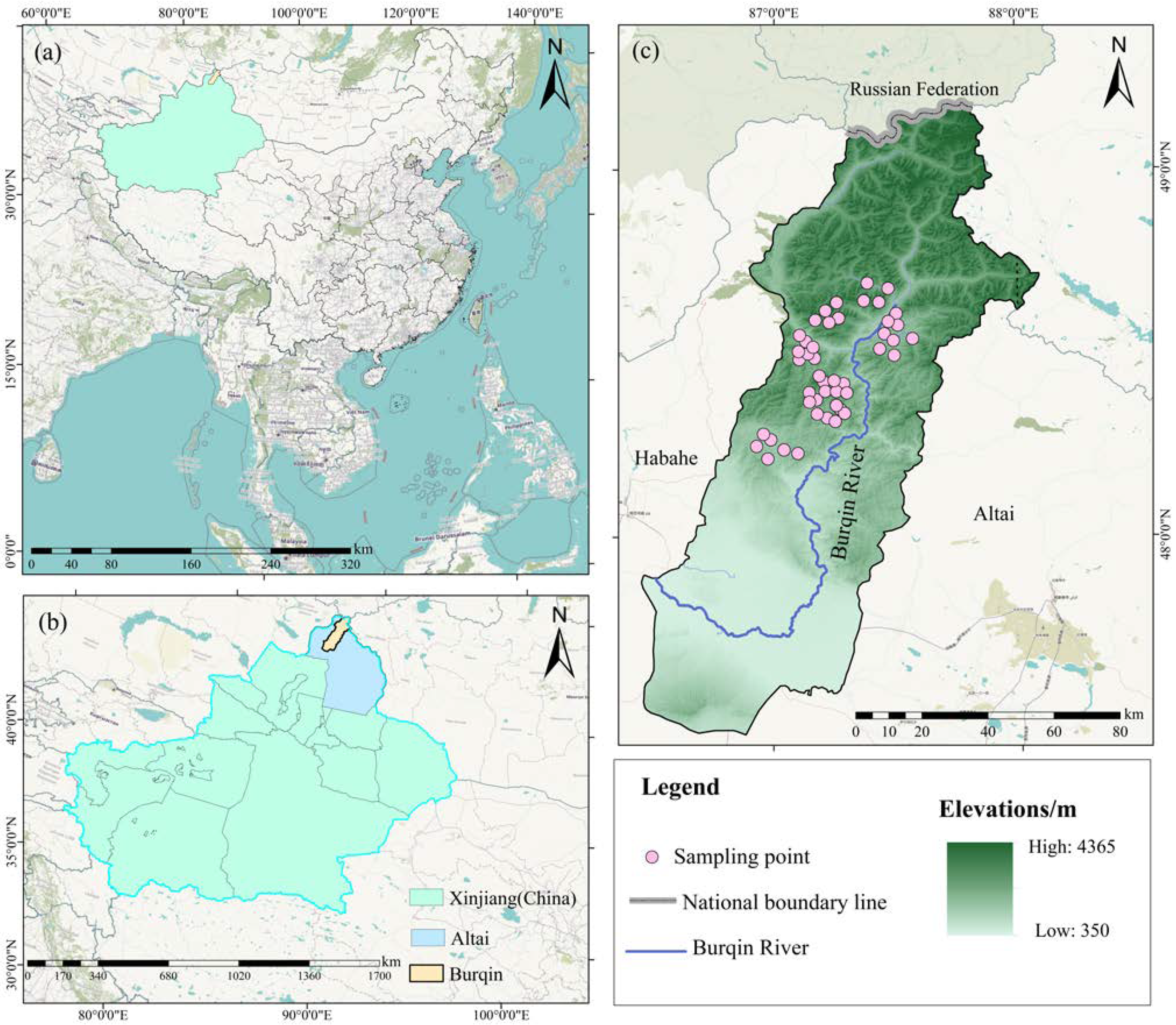
2.1. Study Area
2.2. Design of Experiments
2.3. Calculation of Species Diversity
2.4. Statistical Analysis
3. Results
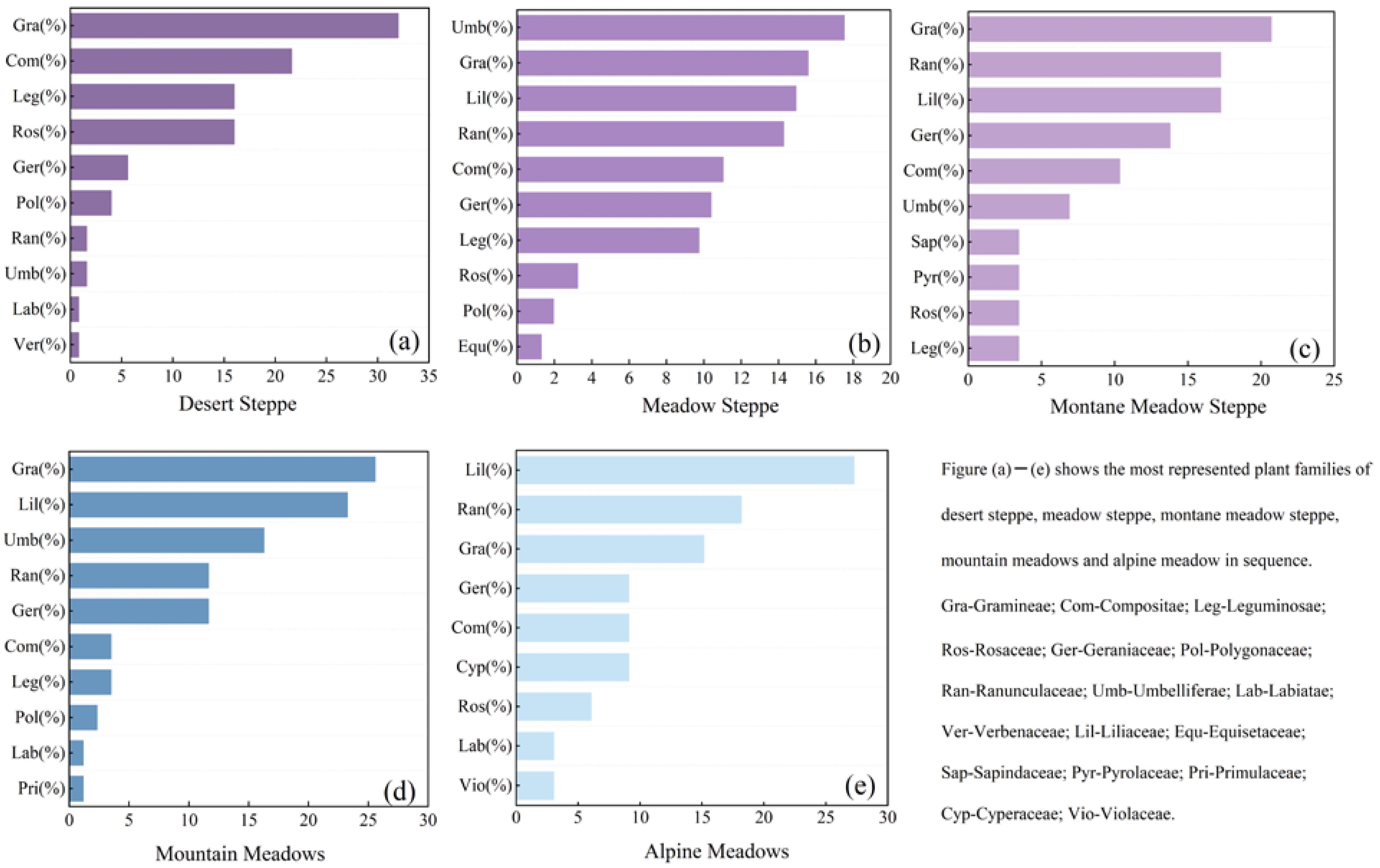
3.1. Plant Family Differences between Grassland Types
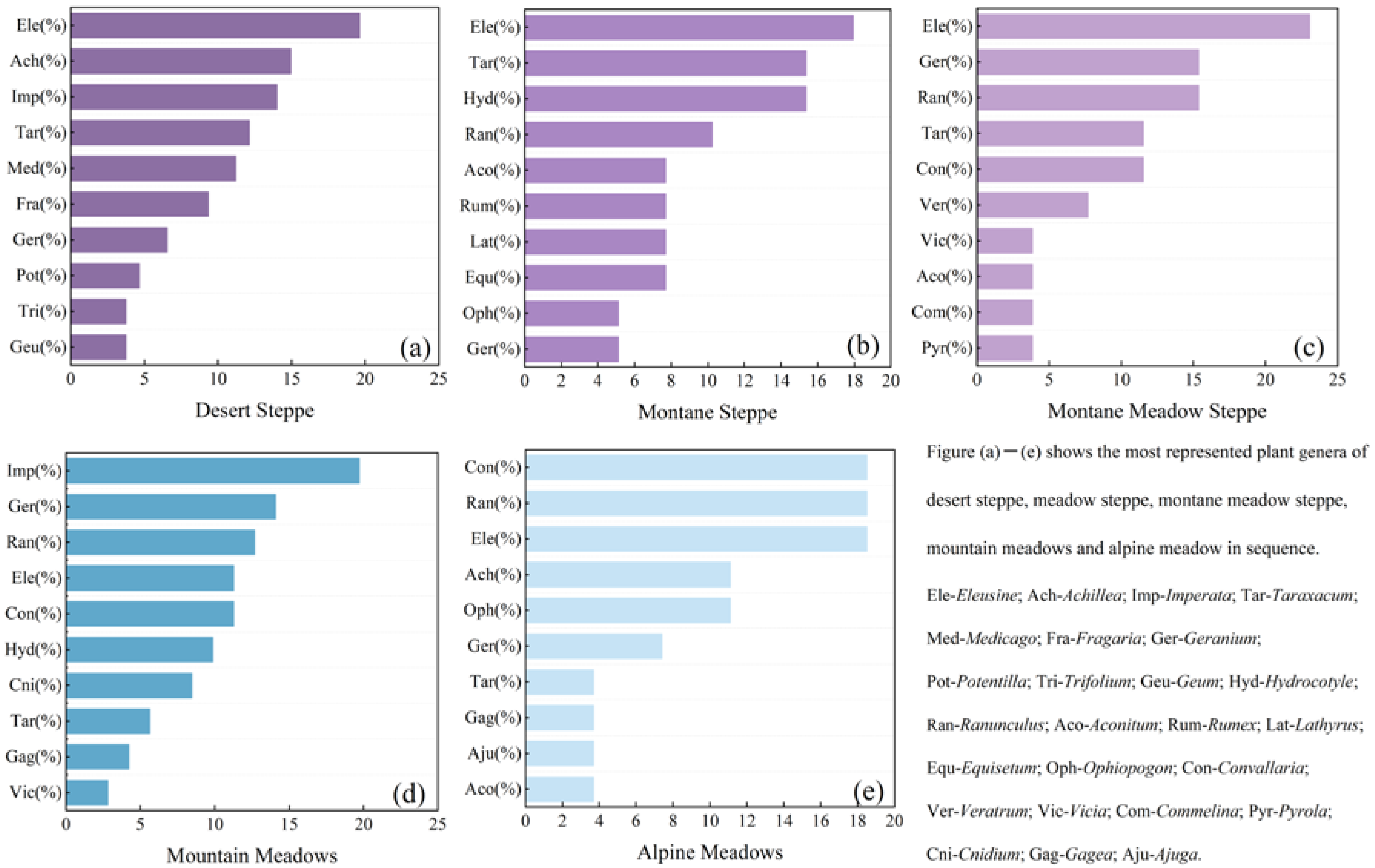
3.2. Characterization of Changes in Plant Family Species in Different Grassland Types
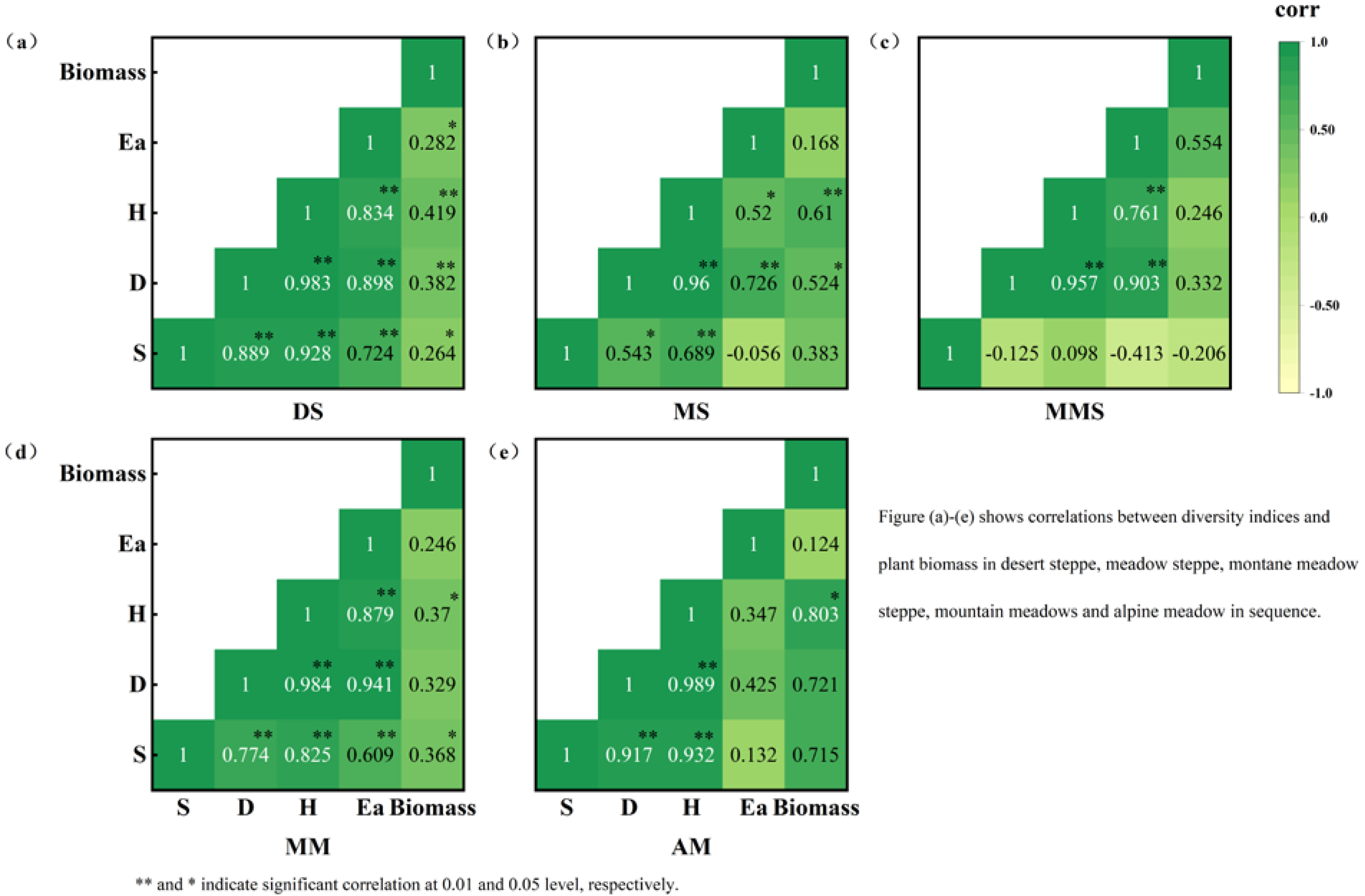
3.3. Similarity between Grassland Types and Correlations between Species Diversity and Above-Ground Biomass
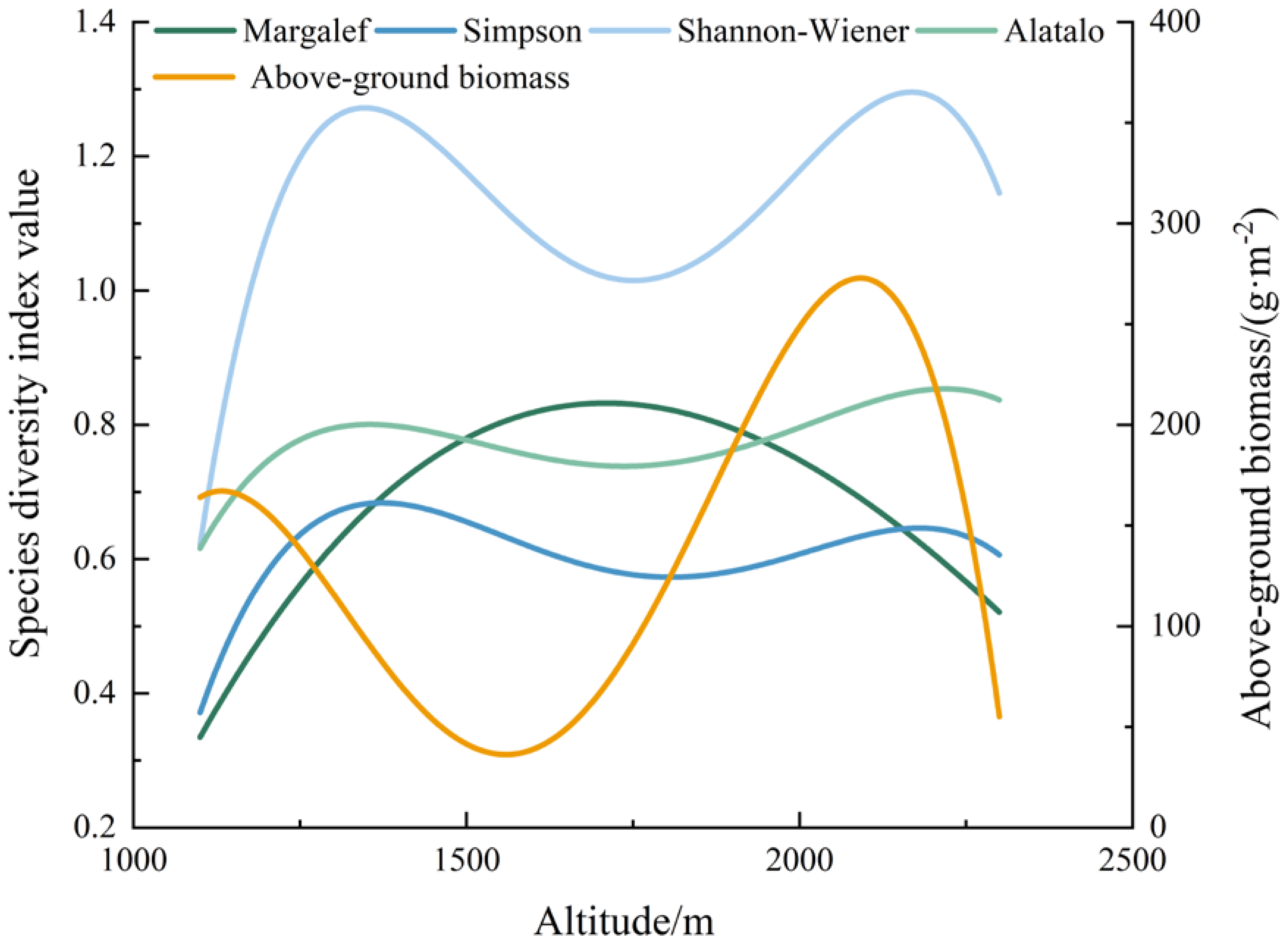
3.4. Vertical Changes in Species Diversity and Above-Ground Biomass
4. Discussion
4.1. Characteristics of Grassland Plant Species in Burqin
4.2. Similarities between Grassland Types and Correlations between Species Diversity and Above-Ground Biomass
4.3. Characteristics of Elevational Changes in Species Diversity and Above-Ground Biomass
5. Conclusions
- (1)
- Desert steppe has Gramineae, Compositae, and Rosaceae as the dominant families, and Eleusine, Achillea, and Imperata as the dominant genera. Montane steppe and mountain meadows have Umbelliferae, Gramineae, and Liliaceae as the dominant families, although these are present in different proportions. Montane steppe has Eleusine, Taraxacum, and Hydrocotyle as the dominant genera. Mountain meadows and montane meadow steppe have Imperata, Geranium, and Ranunculus as the dominant genera. Both montane meadow steppe and alpine meadows are dominated by Gramineae, Ranunculaceae, and Liliaceae, although the dominant genera differ. Convallaria, Ranunculus, and Eleusine are dominant in alpine meadows.
- (2)
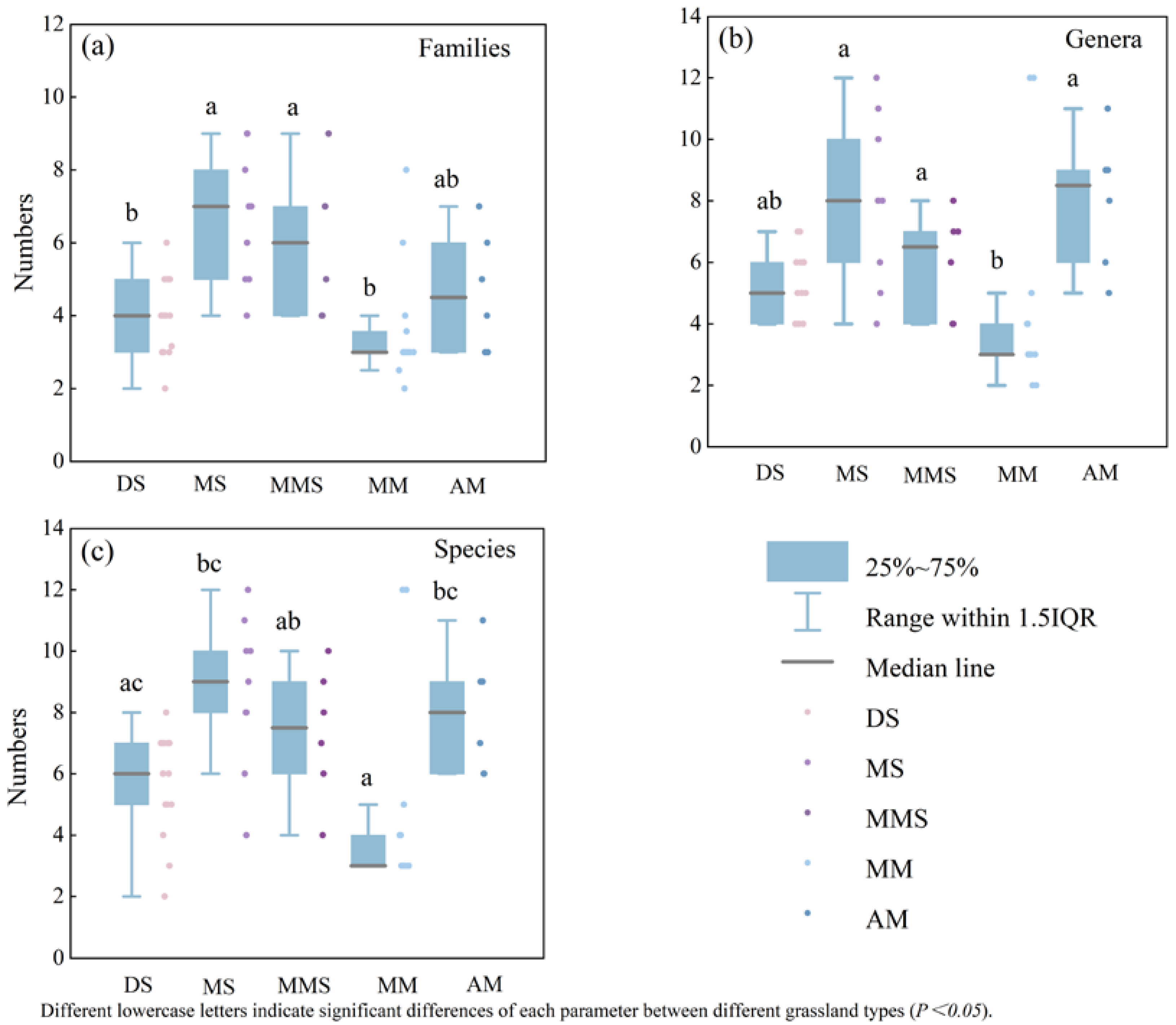
- Variations in the numbers of plants at the family, genus, and species levels are characterized as montane steppe > alpine meadow > montane meadow steppe > desert steppe > mountain meadow.
- (3)
- In DS and MM, the Margalef, Simpson, Alatalo, and Shannon–Wiener indices determine the biomass, and the number of grassland species affects the biomass. The biomass of MS and AM was determined using the Simpson and Shannon–Wiener indices. The number of dominant species determines the biomass of MS and AM. The number of grasslands and the number of species do not have a significant impact on the biomass of MMS, which may be due to human factors such as grazing. The similarity in species composition between different grassland types is low.
- (4)
- The dominant species in the grassland at an altitude of 1200–1400 m is prominent. The number of individuals in the dominant species is large and evenly distributed. The Margalef index peaks at altitudes of 1600–1800 m, but the number of individuals is small, resulting in low biomass and diversity in this range. With the increase in altitude, the Simpson, Alatalo, and Shannon–Wiener indices and biomass peak at an altitude of 2000–2200 m, which is suitable for the survival of most plant species.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ye, X.; Zhou, H.K.; Zhao, X.Q.; Wen, J.; Chen, Z.; Duan, J.C. Review on grassland ecosystem health. Pratacultural Sci. 2011, 28, 549–560. [Google Scholar]
- Gao, Q.H.; Qin, Y.Y.; Liang, M.C.; Gao, X. Interpretation of the Main Conclusions and Suggestions of IPCC AR6 Synthesis Report. Environ. Prot. 2023, 51, 82–84. [Google Scholar]
- Deborah, S.; Valentin, H.K.; Till, K.; Boeddinghaus, R.S.; Hinderling, J.; Ellen, K.; Sven, M.; Sascha, N.; Llja, S.; Susanne, W.; et al. Recovery of Ecosystem functions after experimental disturbance in 73 grasslands differing in land use intensity, plant species richness and community composition. J. Ecol. 2019, 107, 2635–2649. [Google Scholar]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Calderira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 1286, 1123–1127. [Google Scholar] [CrossRef]
- Du, G.Z.; Liu, Z.H. Relationship between species richness and productivity in an alpine meadow plant community. Acta Agrestia Sin. 2003, 27, 125–132. [Google Scholar]
- Yang, L.M.; Zhou, G.S.; Li, J.D. Relationship between productivity and plant species diversity of grassland communities in Songnen plain of northeast China. Acta Agrestia Sin. 2022, 26, 589–593. [Google Scholar]
- Frank, P.; Mikael, P.; Andrea, T.; Florian, A.; Roman, A.; Yves, C.; Fronhofer, E.A.; Pravin, G.; Aurélie, G.; Jason, I.; et al. Biodiversity increases and decreases ecosystem stability. Nature 2018, 10, 17. [Google Scholar]
- Kirkman, K.L.; Mitchell, R.J.; Helton, C.R.; Drew, M.B. Productivity and Species Richness across an Environmental Gradient in a Fire-Dependent Ecosystem. Am. J. Bot. 2001, 11, 2119–2128. [Google Scholar] [CrossRef]
- Yang, X.; Yan, X.H.; Li, M.H.; Tuo, X.X.; Zhang, B.; Wen, Z.M.; Li, W. The relationship between species diversity and aboveground productivity at temporal scale in Yunwushan typical grassland of Ningxia. Acta Agrestia Sin. 2022, 30, 259–268. [Google Scholar]
- Jiang, F.Y.; Wei, X.T.; Kang, B.Y.; Shao, X.Q. Effects of warming on Alpine meadow diversity and primary productivity. Acta Agrestia Sin. 2019, 27, 298–305. [Google Scholar]
- Wu, H.B.; Shui, H.W.; Hu, G.Z.; Wang, X.X.; Gan, Z.Z.B.; Yan, J.; He, S.C.; Xie, W.D.; Gao, Q.Z. Species diversity and biomass distribution patterns of alpine grassland along an elevation gradient in the Northern Tibetan Plateau. Ecol. Environ. Sci. 2019, 28, 1071–1079. [Google Scholar]
- Jia, W.X.; Liu, Y.R.; Zhang, Y.S.; Cao, W.X. Species diversity and biomass of grassland steppe in Qinlian Mountains and their relationships with climate factors. Arid Zone Res. 2015, 32, 1167–1172. [Google Scholar]
- Zhang, Y.; Zhao, J.L.; Xin, X.P.; Wang, M.; Pan, F.J.; Yan, R.R.; Li, L.H. Effects of stocking rate on the interannual patterns of ecosystem biomass and soil nitrogen mineralization in a meadow steppe of northeast China. Plant Soil 2021, 5, 1–23. [Google Scholar] [CrossRef]
- Lu, H.; Cong, J.; Liu, X.; Wang, X.L.; Tang, J.; Li, D.Q.; Zhang, Y.G. Plant diversity patterns a long altitudinal Gradients in alpine meadows in the Three River Headwater Region, China. Acta Agrestia Sin. 2015, 24, 197–204. [Google Scholar]
- Yin, X.K.; Ye, M.; Guo, J.X.; Zhang, K.L.; Zhao, F.F. Study on the relationship between species diversity and productivity of different grassland types in Buerjin forest area of Altai mountains. J. Soil Water Conserv. 2022, 36, 111–115. [Google Scholar]
- Ma, J.J.; Liu, Y.H.; Sheng, J.D.; Li, N.; Wu, H.Q.; Jia, H.T.; Sun, Z.J.; Cheng, J.H. Changes of relationships between dominant species and their relative biomass along elevational gradients in Xinjiang grasslands. Acta Prataculturae Sin. 2021, 30, 25–35. [Google Scholar]
- Li, Q.; He, G.X.; Liu, Z.G.; Guan, W.H.; Qiao, H.H.; Han, T.H.; Sun, B.; Pan, D.R.; Liu, X.N. Responses of vegetation Characteristics and Biodiversity to habitat in Alpine Meadows in Eastern Qi Lian Mountains. Acta Agrestia Sin. 2022, 30, 169–177. [Google Scholar]
- Huang, J.H.; Lu, X.H.; Guo, Z.J. Service functions of the natural forest ecosystem in Burqin Forest Farm, Xinjiang. Arid Zone Res. 2014, 31, 866–873. [Google Scholar]
- Huang, J.H.; Lu, X.H.; Guo, Z.J.; Wang, J.P.; Zang, R.G. Assessment of natural forest ecosystem services in Buerjin County, Xinjiang. J. Beijing For. Univ. 2015, 37, 62–69. [Google Scholar]
- Zhang, F.; Liu, H.; Fang, Y.; Bai, Z.Q.; Ye, G.; Han, Y.L. Stand spatial structure of natural coniferous forest in the Altai Mountains of Xinjiang. J. Anhui Agric. Univ. 2014, 41, 629–635. [Google Scholar]
- Zhang, D.L.; Li, Y.B.; Yang, Y.P.; Lan, B. Synthesized Climate Change in the North Altay Mountains in the Past 2000 Years. Arid Zone Res. 2019, 36, 176–185. [Google Scholar]
- Xu, H.J.; Han, B.P. Main Soil Types and Soil Vertical Distribution in Altay Mountains Area, Xinjiang. Chin. J. Soil Sci. 2008, 29, 465–470. [Google Scholar]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part one). Chin. Biodivers. 1994, 2, 229–239. [Google Scholar]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part two). Chin. Biodivers. 1994, 2, 231–239. [Google Scholar]
- Chen, B.; Ma, R.; Qin, J.H.; Sun, H. The Aboveground Biomass for Ecological Restoration of Highway Construction in the Zoige Wetland, NW Sichuan Province. Hubei Agric. Sci. 2015, 54, 4676–4681. [Google Scholar]
- Zhang, J.L.; Shen, R.; Shi, W.; Liu, X.K.; Ou, X.K. The structure and similarity characteristic of the grassland community in hot-dry valley upper middle and lower of Jinsha River. Ecol. Environ. Sci. 2010, 19, 1272–1277. [Google Scholar]
- Zhao, X.J.; Liang, Z.D.; Shao, L.J.; Zhao, X.F. Analysis and Evaluation on Nonlinear Regression Function of SPSS Software. Stat. Decis. Mak. 2021, 23, 20–22. [Google Scholar]
- Luo, Y.; He, Y.B.; Li, D.Z.; Wang, Y.H.; Yi, T.S.; Wang, H. A Comparison of Classifications of Families of Chinese Vascular Plants among Flora Republicae Popularis Sinicae, Flora of China and the New Classifications. Plant Divers. Resour. 2012, 34, 231–238. [Google Scholar] [CrossRef]
- Wang, M.M.; Liu, X.P.; He, Y.H.; Zhng, T.H.; Wei, J.; Che, L.G.M.; Sun, S.S. How enclosure influences restored plant community changes of different initial types in Horqin Sandy Land. Chin. J. Plant Ecol. 2019, 43, 672–684. [Google Scholar] [CrossRef]
- Liu, L.L.; Sheng, J.D.; Cheng, J.H.; Liu, Y.H.; Li, R.X.; Zhao, D. Relationship between plant species characteristic sand climate factors in different grassland types of Xinjiang. Acta Prataculturae Sin. 2016, 25, 1–12. [Google Scholar]
- He, B.; Wang, G.Y.; Su, H.C.; Shen, Y.P. Response of extreme hydrological events to climate change in the regions of Altay Mountains, Xinjiang. J. Glaciol. Geocryol. 2012, 34, 927–933. [Google Scholar]
- Wu, Y.P.; Chen, K.L.; Zhang, F.; Cao, G.C.; Liu, Z.J.; Su, M.X. Relations between plant species richness and community productivity of typical grasslands in Qinghai Lake Basin. Chin. J. Ecol. 2011, 30, 1449–1453. [Google Scholar]
- Sha, W.; Dong, S.K.; Liu, S.L.; Liu, Q.R.; Shi, J.B.; Li, X.W.; Su, X.K.; Wu, Y. Spatial patterns of plant community biomass and species diversity in Aerjin Mountain Nature Reserve and their influencing factors. Chin. J. Ecol. 2016, 35, 330–337. [Google Scholar]
- He, M.Y.; Wang, Y.X.; Peng, Z.C.; Chang, C.H.; Saman, B.; Liu, Y.J.; Hou, F.J. Spatial pattern of aboveground biomass and species richness of Qilian Mountain grassland. Pratacultural Sci. 2020, 37, 2012–2021. [Google Scholar]
- Sun, X.L.; Kang, S.L.R.; Zhang, Q.; Chang, C.M.; Ma, W.J.; Niu, J.M. Relationship between species diversity, productivity, climatic factors and soil nutrients in the desert steppe. Acta Prataculturae Sin. 2015, 24, 10–19. [Google Scholar]
- Guo, Z.G.; Liang, T.G.; Liu, X.Y.; Zhang, H.J. Species diversity of grassland communities in the Aletai region of the northern Xinjiang province, Acta Bot. Occident. Sin. 2003, 10, 1719–1724. [Google Scholar]
- Yang, L.M.; Han, M.; Li, J.D. Plant diversity change in grassland communities along a grazing disturbance gradient in the northeast China transect. Chin. J. Plant Ecol. 2001, 25, 110–114. [Google Scholar]
- Guo, Q.F. Temporal species richness-biomass relationships along successional gradients. J. Veg. Sci. 2003, 14, 121–128. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Hu, Y.K.; Wang, X.; Gong, Y.M. Vertical differentiation of plant species diversity and biomass in alpine grassland in the middle section of Tianshan Mountains southern slope, Xinjiang of Northwest China. Chin. J. Ecol. 2013, 32, 311–318. [Google Scholar]
- Bai, Y.F.; Li, L.H.; Huang, J.H.; Chen, Z.Z. The influence of plant diversity and functional composition on ecosystem stability of four Stipa communities in the Inner Mongolia Plateau. Acta Bot. Sin. 2001, 43, 280–287. [Google Scholar]
- Tian, Z.P.; Zhuang, L.; Li, J.G. The vertical distribution of vegetation patterns and its relationship with environment factors at the northern slope of Ili River Valley: A bimodal distribution pattern. Acta Ecol. Sin. 2012, 32, 1151–1162. [Google Scholar] [CrossRef]
- Lang, P.; Wang, Y.H.; Xu, H.L.; Zhao, W.Y.; Liu, X.H.; Jinesibieke, M.; Hlehashi, S.; Kulishayila, W. Effects of grazing prohibition years on community characteristics and soil factors in temperate desert grassland. Pratacultural Sci. 2022, 39, 431–442. [Google Scholar]
- Zhang, Y.J.; Zhu, J.T.; Shen, R.N.; Wang, L. Research progress on the effects of grazing on grassland ecosystem. Chin. J. Plant Ecol. 2020, 44, 553–564. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Similarity | DS | MS | MMS | MM | AM |
|---|---|---|---|---|---|
| DS | 1.00 | ||||
| MS | 0.239 | 1.00 | |||
| MMS | 0.093 | 0.382 | 1.00 | ||
| MM | 0.171 | 0.378 | 0.363 | 1.00 | |
| AM | 0.185 | 0.185 | 0.165 | 0.355 | 1.00 |
| Species Diversity Index and Above-Ground Biomass | Fitting Equations | Correlation (R2) | Significance | |
|---|---|---|---|---|
| Altitude | Above-ground biomass | Y = −3.39 × 10−9 x4 + 2.16 × 10−5 x3 − 0.05 x2 + 50.07 x − 18,061.95 | 0.69 | p < 0.05 |
| Margalef | Y = 3.73 × 10−10 x3 − 3.02 × 10−6 x2 + 0.01 x − 4.28 | 0.61 | p < 0.05 | |
| Simpson | Y = −3.43 × 10−12 x4 + 2.45 × 10−8 x3 − 6.45 × 10−5 x2 + 0.07 x − 30.70 | 0.58 | p > 0.05 | |
| Shannon–Wiener | Y = −3.45 × 10−12 x4 + 6.64 × 10−8 x3 − 1.72 × 10−4 x2 + 0.19 x − 78.99 | 0.69 | p < 0.05 | |
| Alatalo | Y= − 2.49 × 10−12 x4 + 1.76 × 10−8 x3 − 4.58 × 10−5 x2 + 0.05 x − 20.83 | 0.71 | p < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, X.; Ye, M.; Xu, X.; He, Q.; Gu, X.; Zeng, G.; Chen, W.; Li, M. Characteristics and Relationships between Species Diversity and Productivity of Different Grassland Types in the Burqin Forest Region of the Altai Mountains. Forests 2023, 14, 1829. https://doi.org/10.3390/f14091829
Pan X, Ye M, Xu X, He Q, Gu X, Zeng G, Chen W, Li M. Characteristics and Relationships between Species Diversity and Productivity of Different Grassland Types in the Burqin Forest Region of the Altai Mountains. Forests. 2023; 14(9):1829. https://doi.org/10.3390/f14091829
Chicago/Turabian StylePan, Xiaoting, Mao Ye, Xingbin Xu, Qingzhi He, Xinchen Gu, Guoyan Zeng, Weilong Chen, and Miaomiao Li. 2023. "Characteristics and Relationships between Species Diversity and Productivity of Different Grassland Types in the Burqin Forest Region of the Altai Mountains" Forests 14, no. 9: 1829. https://doi.org/10.3390/f14091829