
Divergent Decomposition Patterns of Leaf Litter and Fine Roots from an Urban Forest in Mid-Subtropical China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Litter Decomposition and Sample Analyses
2.4. Soil Samples Collection and Analyses
2.5. Data Analysis
3. Results
3.1. The Initial Soil Physicochemical Characteristics and Initial Litter Quality
3.2. Leaf Litter Production and Nutrient Storage
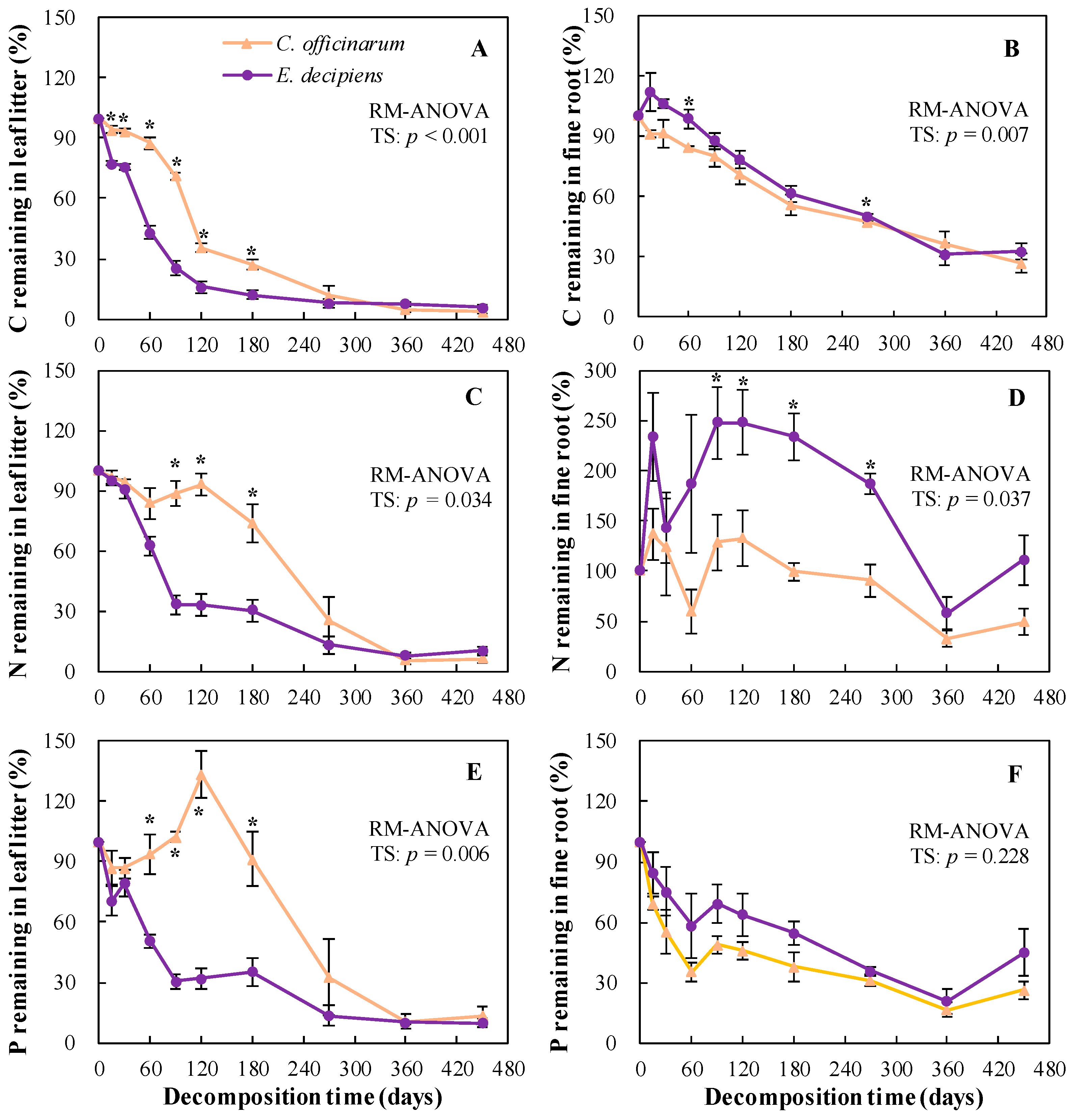
3.3. Litter Decomposition
3.4. Nutrient Mineralization
3.5. The Correlations of Initial Soil Physicochemical Properties and Litter Decomposition
4. Discussion
4.1. Effect of Litter Initial Quality on Litter Decomposition
4.2. Effect of Soil Chemical Properties on Litter Decomposition
4.3. Effect of Soil Microbial Community on Litter Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powers, J.S.; Montgomery, R.A.; Adair, E.C.; Brearley, F.Q.; DeWalt, S.J.; Castanho, C.T.; Chave, J.; Deinert, E.; Ganzhorn, J.U.; Gilbert, M.E.; et al. Decomposition in tropical forests: A pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J. Ecol. 2009, 974, 801–811. [Google Scholar] [CrossRef]
- Cao, J.; He, X.; Chen, Y.; Chen, Y.; Zhang, Y.; Yu, S.; Zhou, L.; Liu, Z.; Zhang, C.; Fu, S. Leaf litter contributes more to soil organic carbon than fine roots in two 10-year-old subtropical plantations. Sci. Total Environ. 2020, 704, 135341. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 9414, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Han, S.; Zhou, Y.; Zhang, J.; Zheng, X.; Dai, G.; Li, M.H. Fine root growth and contribution to soil carbon in a mixed mature Pinus koraiensis forest. Plant Soil 2016, 400, 275–284. [Google Scholar] [CrossRef]
- Tresch, S.; Frey, D.; Le Bayon, R.C.; Zanetta, A.; Rasche, F.; Fliessbach, A.; Moretti, M. Litter decomposition driven by soil fauna, plant diversity and soil management in urban gardens. Sci. Total Environ. 2019, 658, 1614–1629. [Google Scholar] [CrossRef]
- Yang, Z.; Fang, C.; Li, G.; Mu, X. Integrating multiple semantics data to assess the dynamic change of urban green space in Beijing, China. Int. J. Appl. Earth Obs. 2021, 103, 102479. [Google Scholar] [CrossRef]
- Shahtahmassebi, A.R.; Li, C.; Fan, Y.; Wu, Y.; Gan, M.; Wang, K.; Malik, A.; Blackburn, G.A. Remote sensing of urban green spaces: A review. Urban For. Urban Green. 2021, 57, 126946. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Baker, L.A.; Buyarski, C.; Nidzgorski, D.; Finlay, J.C. Decomposition of tree leaf litter on pavement: Implications for urban water quality. Urban Ecosyst. 2014, 17, 369–385. [Google Scholar] [CrossRef]
- Creamer, C.A.; de Menezes, A.B.; Krull, E.S.; Sanderman, J.; Newton-Walters, R.; Farrell, M. Microbial community structure mediates response of soil C decomposition to litter addition and warming. Soil Biol. Biochem. 2015, 80, 175–188. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Ruiz-Peinado, R.; Onrubia, R.; del Río, M. Thinning alters the early-decomposition rate and nutrient immobilization-release pattern of foliar litter in Mediterranean oak-pine mixed stands. For. Ecol. Manage. 2017, 391, 309–320. [Google Scholar] [CrossRef]
- Harmon, M.E.; Silver, W.L.; Fasth, B.; Chen, H.U.A.; Burke, I.C.; Parton, W.J.; Hart, S.C.; Currie, W.S.; Lidet. Long-term patterns of mass loss during the decomposition of leaf and fine root litter: An intersite comparison. Glob. Chang. Biol. 2009, 155, 1320–1338. [Google Scholar] [CrossRef]
- Müller, K.; Marhan, S.; Kandeler, E.; Poll, C. Carbon flow from litter through soil microorganisms: From incorporation rates to mean residence times in bacteria and fungi. Soil Biol. Biochem. 2017, 115, 187–196. [Google Scholar] [CrossRef]
- Seidelmann, K.N.; Scherer-Lorenzen, M.; Niklaus, P.A. Direct vs. microclimate-driven effects of tree species diversity on litter decomposition in young subtropical forest stands. PLoS ONE 2016, 118, e0160569. [Google Scholar] [CrossRef]
- Guo, L.; Deng, M.; Yang, S.; Liu, W.; Wang, X.; Wang, J.; Liu, L. The coordination between leaf and fine root litter decomposition and the difference in their controlling factors. Glob. Ecol. Biogeogr. 2021, 30, 2286–2296. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef]
- Tong, J.; Xiang, W.; Liu, C.; Lei, P.; Tian, D.; Deng, X.; Peng, C. Tree species effects on fine root decomposition and nitrogen release in subtropical forests in southern China. Plant Ecol. Divers. 2012, 53, 323–331. [Google Scholar] [CrossRef]
- Eldhuset, T.D.; Kjønaas, O.J.; Lange, H. Decomposition rates and nutrient dynamics of Picea abies needles, twigs and fine roots after stem-only harvesting in eastern and western Norway. Plant Soil 2017, 418, 357–375. [Google Scholar] [CrossRef]
- Vaidelys, T.; Straigytė, L.; Manton, M. Effects of seasonality, tree species and urban green space on deciduous leaf litter decomposition in Lithuania. Sustainability 2020, 12, 2210. [Google Scholar] [CrossRef]
- Pavao-Zuckerman, M.A.; Coleman, D.C. Decomposition of chestnut oak (Quercus prinus) leaves and nitrogen mineralization in an urban environment. Biol. Fertil. Soils 2005, 41, 343–349. [Google Scholar] [CrossRef]
- Tan, X.; Kan, L.; Su, Z.; Liu, X.; Zhang, L. The composition and diversity of soil bacterial and fungal communities along an urban-to-rural gradient in South China. Forests 2019, 10, 797. [Google Scholar] [CrossRef]
- Yin, S.; Wang, F.; Xiao, Y.; Xue, S. Comparing cooling efficiency of shading strategies for pedestrian thermal comfort in street canyons of traditional shophouse neighbourhoods in Guangzhou, China. Urban Clim. 2022, 43, 101165. [Google Scholar] [CrossRef]
- Mackintosh, T.J.; Davis, J.A.; Thompson, R.M. Impacts of multiple stressors on ecosystem function: Leaf decomposition in constructed urban wetlands. Environ. Pollut. 2016, 208, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Vourlitis, G.L.; van der Veen, E.L.; Cangahuala, S.; Jaeger, G.; Jensen, C.; Fissore, C.; Wood, E.M.; Abraham, J.K.; Whittemore, K.S.; Slaven, E.; et al. Examining decomposition and nitrogen mineralization in five common urban habitat types across southern California to inform sustainable landscaping. Urban Sci. 2022, 6, 61. [Google Scholar] [CrossRef]
- Nikula, S.; Vapaavuori, E.; Manninen, S. Urbanization-related changes in European aspen (Populus tremula L.): Leaf traits and litter decomposition. Environ. Pollut. 2010, 158, 2132–2142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, X.; Wang, Q. Characteristics of N mineralization in urban soils of Hefei, East China. Pedosphere 2010, 20, 236–244. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, X.; Yang, J.; Tan, S.; Liao, S.; Wu, F. Nitrogen and phosphorus resorption and stoichiometric characteristics of different tree species in a mid-subtropical common-garden, China. Chin. J. Appl. Ecol. 2021, 32, 1154–1162. (In Chinese) [Google Scholar]
- Chen, Y.; Zhang, Y.; Cao, J.; Fu, S.; Hu, S.; Wu, J.; Zhao, J.; Liu, Z. Stand age and species traits alter the effects of understory removal on litter decomposition and nutrient dynamics in subtropical Eucalyptus plantations. Glob. Ecol. Conserv. 2019, 20, e00693. [Google Scholar] [CrossRef]
- Bao, S. Analysis in Soil and Agrochemistry, 3rd ed.; Agricultural Press: Beijing, China, 2000; pp. 30–78. (In Chinese) [Google Scholar]
- Bossio, D.A.; Scow, K.M. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Szefer, P.; Carmona, C.P.; Chmel, K.; Konečná, M.; Libra, M.; Molem, K.; Novotný, V.; Segar, S.T.; Švamberková, E.; Topliceanu, T.; et al. Determinants of litter decomposition rates in a tropical forest: Functional traits, phylogeny and ecological succession. Oikos 2017, 126, 1101–1111. [Google Scholar] [CrossRef]
- Tanjung, F.A.; Kuswardani, R.A.; Idumah, C.I.; Siregar, J.P.; Karim, A. Characterization of mechanical and thermal properties of esterified lignin modified polypropylene composites filled with chitosan fibers. Polym. Polym. Compos. 2022, 30, 09673911221082482. [Google Scholar] [CrossRef]
- Donnelly, P.K.; Entry, J.A.; Crawford, D.L.; Cromack, K. Cellulose and lignin degradation in forest soils: Response to moisture, temperature, and acidity. Microb. Ecol. 1990, 20, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Yin, R.; Xu, Z.; Zhang, L.; You, C.; Liu, Y.; Li, H.; Wang, L.; Liu, S.; Zhang, Y.; et al. Forest gaps slow lignin and cellulose degradation of fir (Abies faxoniana) twig litter in an alpine forest. Geoderma 2022, 424, 116010. [Google Scholar] [CrossRef]
- See, C.R.; Luke McCormack, M.; Hobbie, S.E.; Flores-Moreno, H.; Silver, W.L.; Kennedy, P.G. Global patterns in fine root decomposition: Climate, chemistry, mycorrhizal association and woodiness. Ecol. Lett. 2019, 22, 946–953. [Google Scholar] [CrossRef]
- Taylor, B.R.; Parkinson, D.; Parsons, W.F. Nitrogen and lignin content as predictors of litter decay rates: A microcosm test. Ecology 1989, 70, 97–104. [Google Scholar] [CrossRef]
- Bonanomi, G.; Idbella, M.; Zotti, M.; Santorufo, L.; Motti, R.; Maisto, G.; de Marco, A. Decomposition and temperature sensitivity of fine root and leaf litter of 43 mediterranean species. Plant Soil 2021, 464, 453–465. [Google Scholar] [CrossRef]
- Siqueira, D.P.; de Carvalho, G.C.M.W.; de Souza Silva, J.G.; Caldeira, M.V.W.; Barroso, D.G. Litter decomposition and nutrient release for two tropical N-fixing species in Rio de Janeiro, Brazil. J. For. Res. 2022, 33, 487–496. [Google Scholar] [CrossRef]
- Spohn, M.; Berg, B. Import and release of nutrients during the first five years of plant litter decomposition. Soil Biol. Biochem. 2023, 176, 108878. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Lv, X.; Wang, R.; Wang, D.; Zhang, F. Contrasting dynamics and factor controls in leaf compared with different-diameter fine root litter decomposition in secondary forests in the Qinling Mountains after 5 years of whole-tree harvesting. Sci. Total Environ. 2022, 838, 156194. [Google Scholar] [CrossRef]
- Lin, C.; Yang, Y.; Guo, J.; Chen, G.; Xie, J. Fine root decomposition of evergreen broadleaved and coniferous tree species in mid-subtropical China: Dynamics of dry mass, nutrient and organic fractions. Plant Soil 2011, 338, 311–327. [Google Scholar] [CrossRef]
- Song, X.; Li, Q.; Gu, H. Effect of nitrogen deposition and management practices on fine root decomposition in Moso bamboo plantations. Plant Soil 2017, 410, 207–215. [Google Scholar] [CrossRef]
- Aerts, A.; de Caluwe, H. Nutritional and plant-mediated controls on leaf litter decomposition of Carex species. Ecology 1997, 78, 244–260. [Google Scholar] [CrossRef]
- Ge, X.; Zeng, L.; Xiao, W.; Huang, Z.; Geng, X.; Tan, B. Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review. Acta. Ecol. Sin. 2013, 33, 102–108. [Google Scholar] [CrossRef]
- Cusack, D.F.; Chou, W.W.; Yang, W.H.; Harmon, M.E.; Silver, W.L.; Lidet, T. Controls on long-term root and leaf litter decomposition in neotropical forests. Global Chang. Biol. 2009, 15, 1339–1355. [Google Scholar] [CrossRef]
- Wambsganss, J.; Freschet, G.T.; Beyer, F.; Bauhus, J.; Scherer-Lorenzen, M. Tree diversity, initial litter quality, and site conditions drive early-stage fine-root decomposition in European forests. Ecosystems 2022, 25, 1493–1509. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Pihlatie, M.; Korhonen, J.F.; Levula, J.; Frumau, A.K.; Ibrom, A.; Lembrechts, J.J.; Morillas, L.; Horváth, L.; Jones, S.K.; et al. Climatic controls on leaf litter decomposition across European forests and grasslands revealed by reciprocal litter transplantation experiments. Biogeosciences 2016, 13, 1621–1633. [Google Scholar] [CrossRef]
- Solly, E.F.; Schöning, I.; Boch, S.; Kandeler, E.; Marhan, S.; Michalzik, B.; Müller, J.; Zscheischler, J.; Trumbore, S.E.; Schrumpf, M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant Soil 2014, 382, 203–218. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Cao, J.; Fu, S.; Wang, J.; Lambers, H.; Liu, Z. Divergent responses of fine root decomposition to removal of understory plants and overstory trees in subtropical Eucalyptus urophylla plantations. Plant Soil 2022, 476, 639–652. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Li, X.; Yang, Z.; Xiong, D.; Xu, C.; Wanek, W.; Yang, Y. Soil warming delays leaf litter decomposition but exerts no effect on litter nutrient release in a subtropical natural forest over 450 days. Geoderma 2022, 427, 116139. [Google Scholar] [CrossRef]
- Ruan, H.; Li, Y.; Zou, X. Soil communities and plant litter decomposition as influenced by forest debris: Variation across tropical riparian and upland sites. Pedobiologia 2005, 49, 529–538. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, C.; Wang, F.; Zhao, G.; Pu, G.; Ma, X.; Tian, X. Effects of nitrogen addition on litter decomposition, soil microbial biomass, and enzyme activities between leguminous and non-leguminous forests. Ecol. Res. 2013, 28, 793–800. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Zhang, W.; Shao, Y.; Duan, H.; Chen, B.; Wei, X.; Fan, H. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Argiroff, W.A.; Zak, D.R.; Upchurch, R.A.; Pellitier, P.T.; Belke, J.P. Fungal community composition and genetic potential regulate fine root decay in northern temperate forests. Mol. Ecol. 2023, 32, 2005–2021. [Google Scholar] [CrossRef]
- Su, Z.; Su, B.; Wu, Y.; Zhang, Y.; Wang, J.; Chen, Y.; Shangguan, Z. A less complex but more specialized microbial network resulted in faster fine-root decomposition in young stands of Robinia pseudoacacia. Appl. Soil Ecol. 2023, 182, 104735. [Google Scholar] [CrossRef]
- Li, A.; Fahey, T.J.; Pawlowska, T.E.; Fisk, M.C.; Burtis, J. Fine root decomposition, nutrient mobilization and fungal communities in a pine forest ecosystem. Soil Biol. Biochem. 2015, 83, 76–83. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. officinarum | E. decipiens | F-Value | p-Value | |
|---|---|---|---|---|
| pH | 5.31 ± 0.12 | 4.48 ± 0.07 | 27.93 | 0.003 |
| SOC (mg g−1) | 22.90 ± 2.20 | 13.10 ± 1.20 | 15.33 | 0.008 |
| TN (mg g−1) | 1.58 ± 0.21 | 1.00 ± 0.16 | 4.80 | 0.071 |
| TP (mg g−1) | 0.27 ± 0.02 | 0.22 ± 0.05 | 0.98 | 0.360 |
| C:N | 14.81 ± 1.19 | 13.85 ± 1.87 | 0.19 | 0.681 |
| N:P | 5.98 ± 0.64 | 4.91 ± 0.67 | 1.32 | 0.295 |
| C:P | 86.29 ± 4.48 | 68.24 ± 13.45 | 1.62 | 0.250 |
| ST | 20.41 ± 1.38 | 20.51 ± 1.41 | 0.52 | 0.500 |
| SMC | 26.40 ± 2.00 | 22.05 ± 1.56 | 11.73 | 0.014 |
| C. officinarum | E. decipiens | F-Value | p-Value | ||
|---|---|---|---|---|---|
| Leaf litter | C (%) | 51.12 ± 0.49 | 45.23 ± 1.20 | 20.73 | 0.004 |
| N (mg g−1) | 8.28 ± 0.42 | 10.58 ± 0.20 | 24.33 | 0.003 | |
| P (mg g−1) | 0.40 ± 0.03 | 0.67 ± 0.04 | 33.91 | 0.001 | |
| C:N | 62.20 ± 2.93 | 42.75 ± 0.87 | 40.41 | 0.001 | |
| N:P | 20.84 ± 0.47 | 16.04 ± 0.83 | 25.54 | 0.002 | |
| C:P | 1299.14 ± 84.49 | 687.52 ± 50.35 | 38.67 | 0.001 | |
| Fine root | C (%) | 45.70 ± 1.77 | 38.36 ± 1.23 | 11.58 | 0.014 |
| N (mg g−1) | 10.81 ± 1.86 | 3.27 ± 0.24 | 16.17 | 0.007 | |
| P (mg g−1) | 1.66 ± 0.14 | 0.83 ± 0.09 | 25.13 | 0.002 | |
| C:N | 46.56 ± 9.03 | 119.25 ± 9.77 | 29.85 | 0.002 | |
| N:P | 6.56 ± 1.18 | 3.97 ± 0.19 | 4.70 | 0.073 | |
| C:P | 281.3 ± 25.81 | 476.11 ± 52.21 | 11.19 | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Chen, S.; Zhang, B.; Ma, X.; Liu, X.; Huang, Y.; Zhang, Y. Divergent Decomposition Patterns of Leaf Litter and Fine Roots from an Urban Forest in Mid-Subtropical China. Forests 2023, 14, 1741. https://doi.org/10.3390/f14091741
Chen Y, Chen S, Zhang B, Ma X, Liu X, Huang Y, Zhang Y. Divergent Decomposition Patterns of Leaf Litter and Fine Roots from an Urban Forest in Mid-Subtropical China. Forests. 2023; 14(9):1741. https://doi.org/10.3390/f14091741
Chicago/Turabian StyleChen, Yuanqi, Shengfu Chen, Bohan Zhang, Xiaotong Ma, Xiaotong Liu, Ying Huang, and Yu Zhang. 2023. "Divergent Decomposition Patterns of Leaf Litter and Fine Roots from an Urban Forest in Mid-Subtropical China" Forests 14, no. 9: 1741. https://doi.org/10.3390/f14091741