
Responses of Plant Species Diversity and Biomass to Forest Management Practices after Pine Wilt Disease

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Plant Data Collection
2.3. Environmental Data Measurement
2.4. Biomass Estimation
2.5. Data Analysis
3. Results
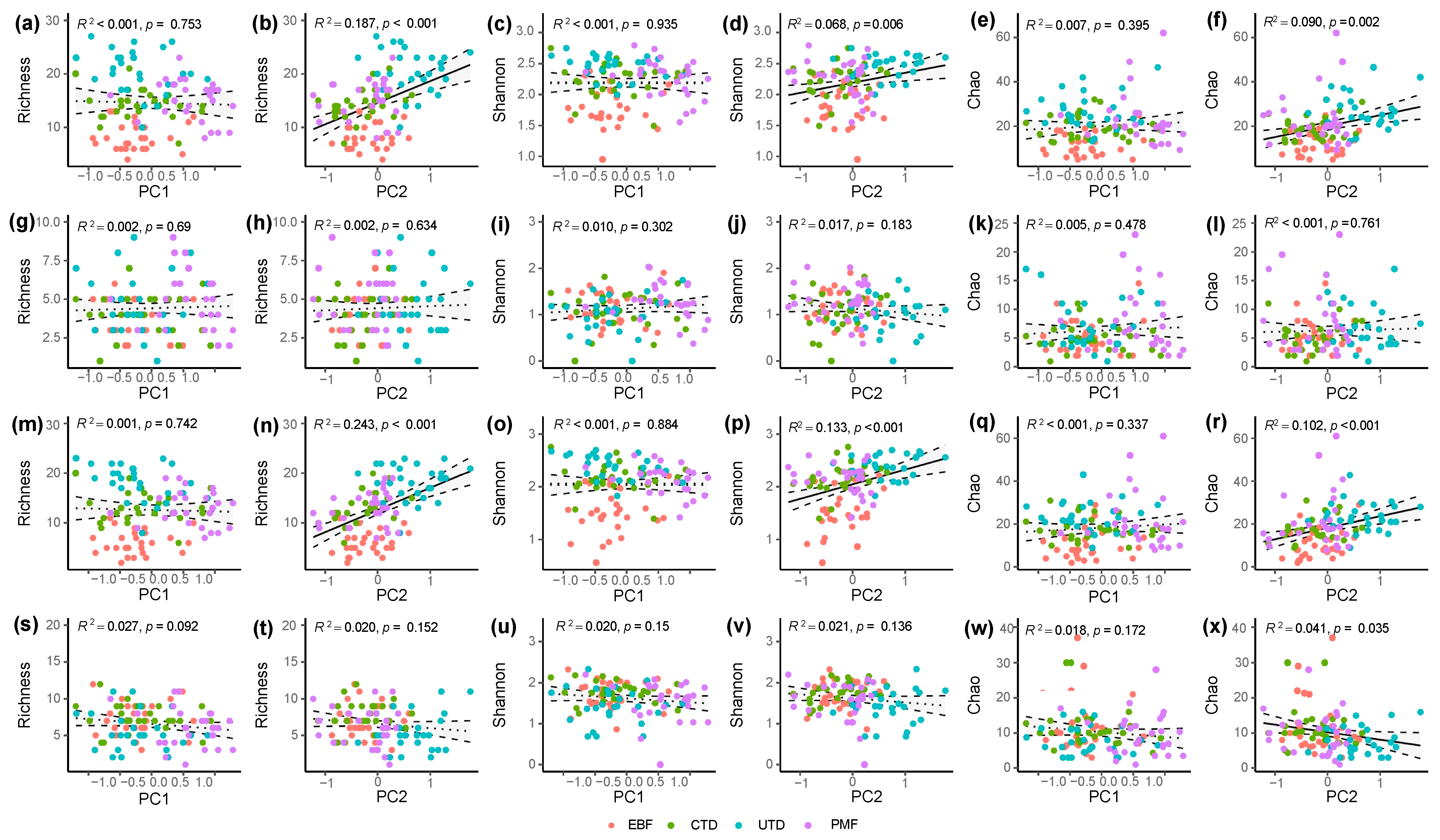
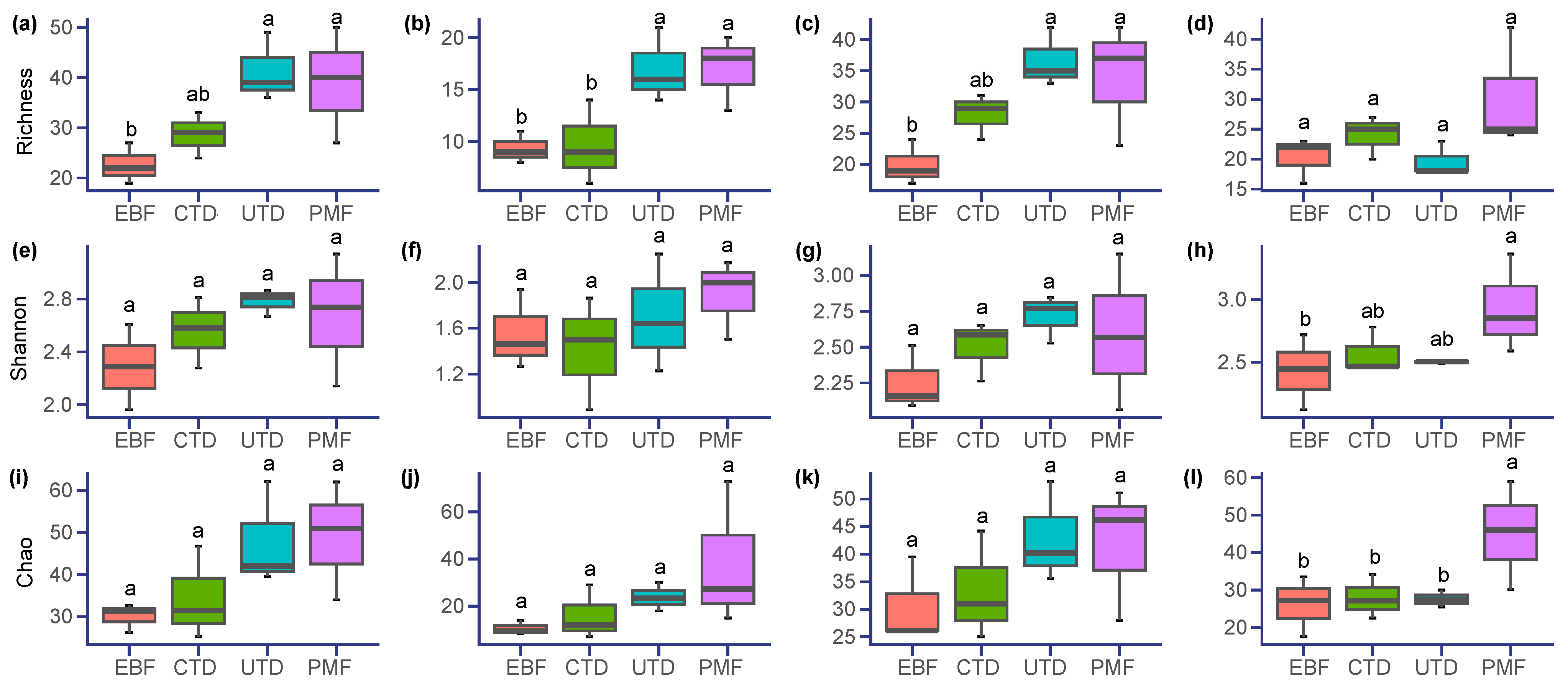
3.1. Differences in Plant Alpha Diversity across Forest Types
3.2. Differences in Plant Species Composition across Forest Types
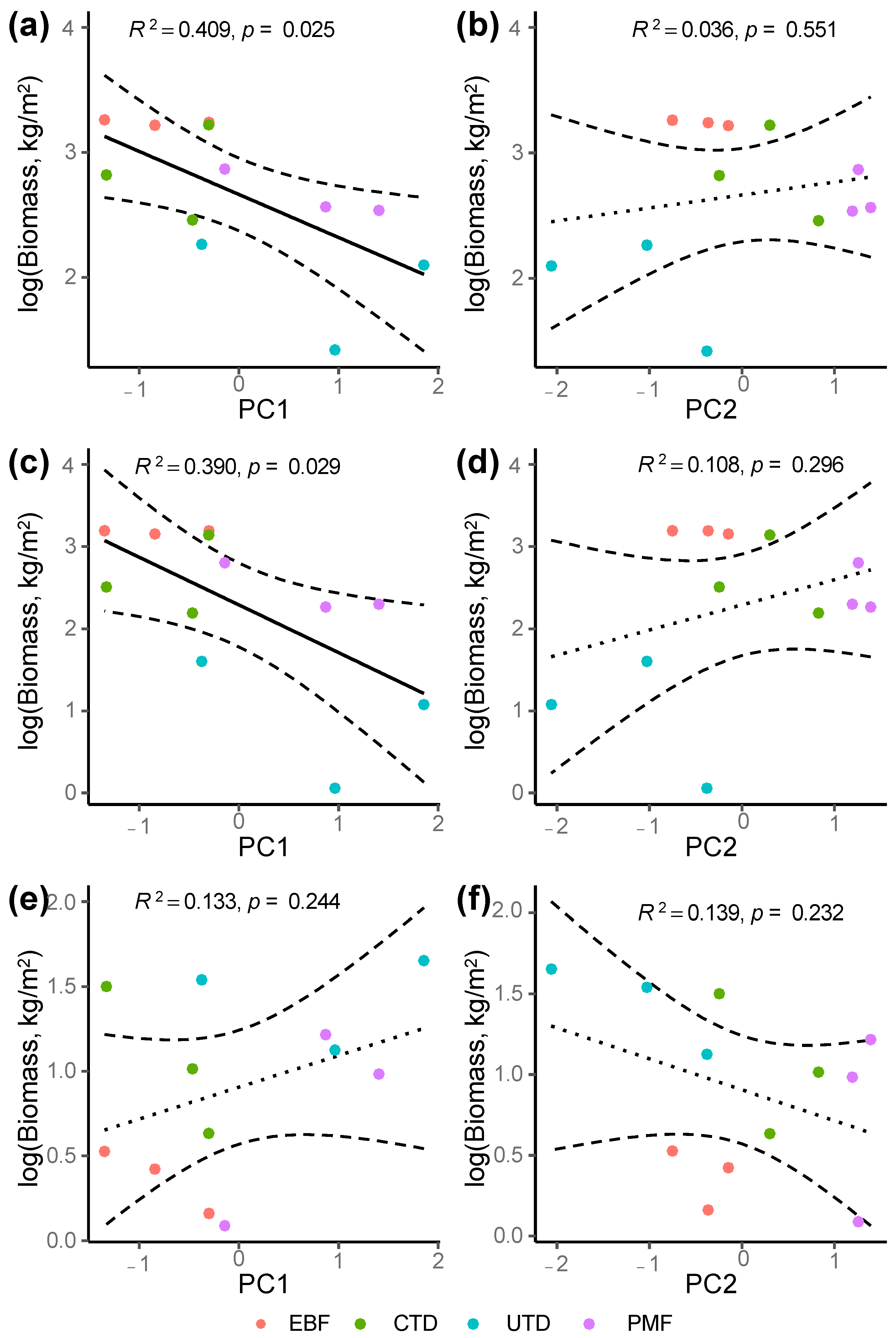
3.3. Differences in Plant Biomass across Forest Types
4. Discussion
4.1. Impact of the Removal of Diseased Pine Trees on Alpha Diversity
4.2. Impact of the Removal of Diseased Pine Trees on Species Composition
4.3. Impact of the Removal of Diseased Pine Trees on Biomass
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






References
- Webster, J.; Mota, M. Pine Wilt Disease: Global Issues, Trade and Economic Impact; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- de la Fuente, B.; Saura, S.; Beck, P.S.A.; Fortin, M.-J. Predicting the spread of an invasive tree pest: The pine wood nematode in Southern Europe. J. Appl. Ecol. 2018, 55, 2374–2385. [Google Scholar] [CrossRef]
- Yu, M.J.; Xu, X.H.; Ding, P. Economic loss versus ecological gain: The outbreaks of invaded pinewood nematode in China. Biol. Invasions 2011, 13, 1283–1290. [Google Scholar] [CrossRef]
- Kim, B.-N.; Kim, J.H.; Ahn, J.-Y.; Kim, S.; Cho, B.-K.; Kim, Y.-H.; Min, J. A short review of the pinewood nematode, Bursaphelenchus xylophilus. Toxicol. Env. Health 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente, B.; Beck, P.S.A. Management measures to control pine wood nematode spread in Europe. J. Appl. Ecol. 2019, 56, 2577–2580. [Google Scholar] [CrossRef]
- Filipiak, A. The pine wilt disease. Sylwan 2008, 152, 9–19. [Google Scholar]
- Yang, B.J. The History, Dispersal and Potential Threat of Pine Wood Nematode in China; Brill Academica Publishers: Leiden, The Netherlands, 2003. [Google Scholar]
- Fonseca, L.; Cardoso, J.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine Wilt Disease: A threat to European forestry. Eur. J. Plant Pathol. 2011, 133, 89–99. [Google Scholar] [CrossRef]
- Tang, X.; Yuan, Y.; Li, X.; Zhang, J. Maximum Entropy Modeling to Predict the Impact of Climate Change on Pine Wilt Disease in China. Front. Plant Sci. 2021, 12, 652500. [Google Scholar] [CrossRef]
- Kwon, T.S.; Shin, J.H.; Lim, J.H.; Kim, Y.K.; Lee, E.J. Management of pine wilt disease in Korea through preventative silvicultural control. Forest. Ecol. Manag. 2011, 261, 562–569. [Google Scholar] [CrossRef]
- Robinet, C.; Castagnone-Sereno, P.; Mota, M.; Roux, G.; Sarniguet, C.; Tassus, X.; Jactel, H.; Marini, L. Effectiveness of clear-cuttings in non-fragmented pine forests in relation to EU regulations for the eradication of the pine wood nematode. J. Appl. Ecol. 2020, 57, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Gordillo, L.F.; Kim, Y. A simulation of the effects of early eradication of nematode infected trees on spread of pine wilt disease. Eur. J. Plant Pathol. 2011, 132, 101–109. [Google Scholar] [CrossRef]
- Hao, X.; Liu, X.; Chen, J.; Wang, B.; Li, Y.; Ye, Y.; Ma, W.; Ma, L. Effects on community composition and function Pinus massoniana infected by Bursaphelenchus xylophilus. BMC Microbiol. 2022, 22, 157. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Shi, J.; Huang, R.; Wang, Z.; Luo, Y. Effects of pine wilt disease invasion on soil properties and Masson pine forest communities in the Three Gorges reservoir region, China. Ecol. Evol. 2015, 5, 1702–1716. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, M. Development of secondary pine forests after pine wilt disease in western Japan. J. Veg. Sci. 1996, 75, 729–738. [Google Scholar] [CrossRef]
- Dalling, J.W.; Hubbell, S.P. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J. Ecol. 2002, 90, 557–568. [Google Scholar] [CrossRef]
- Jin, Y.; Russo, S.E.; Yu, M.J. Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest. J. Ecol. 2018, 106, 1634–1645. [Google Scholar] [CrossRef]
- Hu, G.; Xu, X.; Wang, Y.; Lu, G.; Feeley, K.J.; Yu, M. Regeneration of different plant functional types in a Masson pine forest following pine wilt disease. PLoS ONE 2012, 7, e36432. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, C.; Lee, K.S.; Bolan, N.S.; Naidu, R. Carbon storage and soil CO2 efflux rates at varying degrees of damage from pine wilt disease in red pine stands. Sci. Total. Environ. 2013, 465, 273–278. [Google Scholar] [CrossRef]
- Kim, C.; Jang, K.-S.; Kim, J.-B.; Byun, J.-K.; Lee, C.-H.; Jeon, K.-S. Relationship between soil properties and incidence of pine wilt disease at stand level. Landsc. Ecol. Eng. 2010, 6, 119–124. [Google Scholar] [CrossRef]
- Gao, R.; Luo, Y.; Wang, Z.; Yu, H.; Shi, J. Patterns of biomass, carbon, and nitrogen storage distribution dynamics after the invasion of pine forests by Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) in the three Gorges Reservoir Region. J. For. Res. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Reu, J.C.; Catano, C.P.; Spasojevic, M.J.; Myers, J.A. Beta diversity as a driver of forest biomass across spatial scales. Ecology 2022, 103, e3774. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of Zhejiang (New Edition). Flora of Zhejiang (New Edition); Zhejiang Science and Technology Press: Hangzhou, China, 2021. [Google Scholar]
- Liu, J.; Zhong, Y.; Zhong, L.; Wei, B.; Zheng, S.; Xie, Y.; Jin, Y.; Yu, M. The asymmetric relationships of the distribution of conspecific saplings and adults in forest fragments. J. Plant Ecol. 2020, 13, 398–404. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Zeng, Y.; Lei, P.; Deng, X.; Peng, C. Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south China. For. Ecol. Manag. 2016, 372, 291–302. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity and independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. An improved nonparametric lower bound of species richness via a modified good-turing frequency formula. Biometrics 2014, 70, 671–682. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Walsh, D.C. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Menéndez-Gutiérrez, M.; Alonso, M.; Jiménez, E.; Toval, G.; Mansilla, P.; Abelleira, A.; Abelleira-Sanmartín, A.; Díaz, R. Interspecific variation of constitutive chemical compounds in Pinus spp. xylem and susceptibility to pinewood nematode (Bursaphelenchus xylophilus). Eur. J. Plant Pathol. 2017, 150, 939–953. [Google Scholar] [CrossRef]
- Bonello, P.; Campbell, F.T.; Cipollini, D.; Conrad, A.O.; Farinas, C.; Gandhi, K.J.K.; Hain, F.P.; Parry, D.; Showalter, D.N.; Villari, C.; et al. Invasive tree pests devastate ecosystems—A proposed new response framework. Front. For. Glob. Change 2020, 3, 2. [Google Scholar] [CrossRef]
- Frost, C.M.; Allen, W.J.; Courchamp, F.; Jeschke, J.M.; Saul, W.C.; Wardle, D.A. Using network theory to understand and predict biological invasions. Trends Ecol. Evol. 2019, 34, 831–843. [Google Scholar] [CrossRef]
- Crystal-Ornelas, R.; Lockwood, J.L. Cumulative meta-analysis identifies declining but negative impacts of invasive species on richness after 20 yr. Ecology 2020, 101, e03082. [Google Scholar] [CrossRef] [PubMed]
- Akesson, A.; Curtsdotter, A.; Eklof, A.; Ebenman, B.; Norberg, J.; Barabas, G. The importance of species interactions in eco-evolutionary community dynamics under climate change. Nat. Commun. 2021, 12, 4759. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Tan, C.; Yang, Z.; Li, J.; Xiao, H.; Tong, Y. Regeneration and growth of tree seedlings and saplings in created gaps of different sizes in a subtropical secondary forest in southern China. For. Ecol. Manag. 2022, 511, 120143. [Google Scholar] [CrossRef]
- Ge, X.; Zeng, L.; Xiao, W.; Huang, Z.; Geng, X.; Tan, B. Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review. Acta Ecol. Sin. 2013, 33, 102–108. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bray–Curtis | Sums of Squares | R2 | F | p Value | |
|---|---|---|---|---|---|
| All species | |||||
| EBF vs. CTD | 0.657 | 4.375 | 0.302 | 22.496 | ≤0.001 |
| EBF vs. UTD | 0.805 | 5.964 | 0.391 | 33.317 | ≤0.001 |
| EBF vs. PMF | 0.790 | 5.460 | 0.346 | 27.459 | ≤0.001 |
| CTD vs. UTD | 0.472 | 1.698 | 0.163 | 10.095 | ≤0.001 |
| CTD vs. PMF | 0.567 | 2.782 | 0.221 | 14.0794 | ≤0.001 |
| UTD vs. PMF | 0.590 | 2.968 | 0.249 | 17.194 | ≤0.001 |
| Adults | |||||
| EBF vs. CTD | 0.622 | 4.276 | 0.291 | 21.349 | ≤0.001 |
| EBF vs. UTD | 0.870 | 7.230 | 0.398 | 34.430 | ≤0.001 |
| EBF vs. PMF | 0.810 | 5.679 | 0.358 | 28.955 | ≤0.001 |
| CTD vs. UTD | 0.752 | 3.760 | 0.226 | 15.150 | ≤0.001 |
| CTD vs. PMF | 0.432 | 1.260 | 0.094 | 5.378 | ≤0.001 |
| UTD vs. PMF | 0.735 | 3.851 | 0.233 | 15.782 | ≤0.001 |
| Saplings | |||||
| EBF vs. CTD | 0.724 | 3.244 | 0.202 | 13.173 | ≤0.001 |
| EBF vs. UTD | 0.799 | 4.406 | 0.272 | 19.433 | ≤0.001 |
| EBF vs. PMF | 0.795 | 5.460 | 0.346 | 27.459 | ≤0.001 |
| CTD vs. UTD | 0.434 | 1.699 | 0.163 | 10.095 | ≤0.001 |
| CTD vs. PMF | 0.611 | 3.244 | 0.240 | 16.446 | ≤0.001 |
| UTD vs. PMF | 0.561 | 2.676 | 0.225 | 15.061 | ≤0.001 |
| Seedlings | |||||
| EBF vs. CTD | 0.443 | 0.972 | 0.066 | 3.696 | 0.002 |
| EBF vs. UTD | 0.566 | 2.369 | 0.134 | 8.068 | ≤0.001 |
| EBF vs. PMF | 0.690 | 3.203 | 0.162 | 10.087 | ≤0.001 |
| CTD vs. UTD | 0.504 | 1.504 | 0.084 | 4.799 | ≤0.001 |
| CTD vs. PMF | 0.706 | 2.534 | 0.126 | 7.512 | ≤0.001 |
| UTD vs. PMF | 0.678 | 1.837 | 0.088 | 4.994 | ≤0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, W.; Wu, J.; Wei, B.; Guo, J.; Zhong, L.; Yu, M. Responses of Plant Species Diversity and Biomass to Forest Management Practices after Pine Wilt Disease. Forests 2023, 14, 1636. https://doi.org/10.3390/f14081636
Liu J, Liu W, Wu J, Wei B, Guo J, Zhong L, Yu M. Responses of Plant Species Diversity and Biomass to Forest Management Practices after Pine Wilt Disease. Forests. 2023; 14(8):1636. https://doi.org/10.3390/f14081636
Chicago/Turabian StyleLiu, Jinliang, Weiyong Liu, Jianbin Wu, Boliang Wei, Jing Guo, Lei Zhong, and Mingjian Yu. 2023. "Responses of Plant Species Diversity and Biomass to Forest Management Practices after Pine Wilt Disease" Forests 14, no. 8: 1636. https://doi.org/10.3390/f14081636