
A New Method to Calibrate Cardinal Temperatures for Eucalyptus Plantation †

Abstract
:
1. Introduction
2. Material and Methods
2.1. Experimental Data and Climatic Classification of Sites
2.2. The Models
2.3. Analytical Approach
3. Results and Discussion
3.1. Stem Dry Mass Production and Air Temperature
3.2. Growth Rate Modifier and Models
3.3. Cardinal Temperature and Process-Based Models
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa, M.G.S.; Mazzafera, P.; Balbuena, T.S. Insights into Temperature Modulation of the Eucalyptus Globulus and Eucalyptus Grandis Antioxidant and Lignification Subproteomes. Phytochemistry 2017, 137, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Juri, S.; Toshinori, Y.; Hori, C.; Nagata, A.; Kudoh, Y.; Ooi, T.; Taguchi, S. Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase (RuBisCO)-Mediated de Novo Synthesis of Glycolate-Based Polyhydroxyalkanoate in Escherichia Coli. J. Biosci. Bioeng. 2019, 128, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.Y.; Miličić, G.; Thieulin-Pardo, G.; Bracher, A.; Maxwell, A.; Ciniawsky, S.; Mueller-Cajar, S.; Engen, J.R.; Hartl, F.U.; Wendler, P.; et al. Mechanism of Enzyme Repair by the AAA+ Chaperone Rubisco Activase. Mol. Cell 2017, 67, 744–756.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, D.H. The short-term temperature-dependency of CO2 photosynthetic responses of two Vitis vinifera cultivars grown in a hot climate. Environ. Exp. Bot. 2018, 147, 125–137. [Google Scholar]
- Bassow, S.L.; Ford, E.D.; Kiester, A.R. A Critique of Carbon-Based Tree Growth Models. In Process Modeling of Forest Growth Responses to Environmental Stress; Timber Press: Portland, OR, USA, 1990; pp. 50–57. [Google Scholar]
- Rauscher, H.M.; Isebrands, J.G.; Host, G.E.; Dickson, R.E.; Dickmann, D.I.; Crow, T.R.; Michael, D.A. ECOPHYS: An Ecophysiological Growth Process Model for Juvenile Poplar. Tree Physiol. 1990, 7, 255–281. [Google Scholar] [CrossRef]
- McMurtrie, R.E.; Rook, D.A.; Kelliher, F.M. Modelling the Yield of Pinus Radiata on a Site Limited by Water and Nitrogen. For. Ecol. Manag. 1990, 30, 381–413. [Google Scholar] [CrossRef]
- Korol, R.L.; Running, S.W.; Milner, K.S. Incorporating Intertree Competition into an Ecosystem Model. Can. J. For. Res. 1995, 25, 413–424. [Google Scholar] [CrossRef]
- Landsberg, J.J.; Waring, R.H. A Generalised Model of Forest Productivity Using Simplified Concepts of Radiation-Use Efficiency, Carbon Balance and Partitioning. For. Ecol. Manag. 1997, 95, 209–228. [Google Scholar] [CrossRef]
- Battaglia, M.; Sands, P.; White, D.; Mummery, D. CABALA: A Linked Carbon, Water and Nitrogen Model of Forest Growth for Silvicultural Decision Support. For. Ecol. Manag. 2004, 193, 251–282. [Google Scholar]
- Rouan, L.; Audebert, A.; Luquet, D.; Roques, S.; Dardou, A.; Gozé, E. Cardinal Temperatures Variability within a Tropical Japonica Rice Diversity Panel. Plant Prod. Sci. 2018, 21, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Baath, G.S.; Kakani, V.G.; Gowda, P.H.; Rocateli, A.C.; Northup, B.K.; Singh, H.; Katta, J.R. Guar Responses to Temperature: Estimation of Cardinal Temperatures and Photosynthetic Parameters. Ind. Crops Prod. 2019, 145, 111940. [Google Scholar] [CrossRef]
- Andreucci, M.P.; Moot, D.J.; Black, A.D.; Sedcole, R. A Comparison of Cardinal Temperatures Estimated by Linear and Nonlinear Models for Germination and Bulb Growth of Forage Brassicas. Eur. J. Agron. 2016, 81, 52–63. [Google Scholar] [CrossRef]
- Daibes, L.F.; Cardoso, V.J.M. Seed Germination of a South American Forest Tree Described by Linear Thermal Time Models. J. Therm. Biol. 2018, 76, 156–164. [Google Scholar] [CrossRef]
- Ryan, M.G. Temperature and Tree Growth. Tree Physiol. 2010, 30, 667–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paine, C.E.; Timothy Toby, R.M.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to Fit Nonlinear Plant Growth Models and Calculate Growth Rates: An Update for Ecologists: Nonlinear Plant Growth Models. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Warr, R.L.; Collins, D.H. Bayesian Nonparametric Models for Combining Heterogeneous Reliability Data. Proc. Inst. Mech. Eng. Part O J. Risk Reliab. 2014, 228, 166–175. [Google Scholar] [CrossRef]
- Luo, Q. Temperature Thresholds and Crop Production: A Review. Clim. Chang. 2011, 109, 583–598. [Google Scholar] [CrossRef]
- Sharif, B.; Makowski, D.; Plauborg, F.; Olesen, J.E. Comparison of Regression Techniques to Predict Response of Oilseed Rape Yield to Variation in Climatic Conditions in Denmark. Eur. J. Agron. 2017, 82, 11–20. [Google Scholar] [CrossRef]
- Kim, S.; Kano, M.; Nakagawa, H.; Hasebe, S. Input Variable Scaling for Statistical Modeling. Comput. Chem. Eng. 2015, 74, 59–65. [Google Scholar] [CrossRef]
- Shoaib, S.A.; Marshall, L.; Sharma, A. Attributing Uncertainty in Streamflow Simulations Due to Variable Inputs via the Quantile Flow Deviation Metric. Adv. Water Resour. 2018, 116, 40–55. [Google Scholar] [CrossRef]
- Souza, L.C.; Souza, R.M.C.R.; Amaral, G.J.A.; Silva Filho, T.M. A Parametrized Approach for Linear Regression of Interval Data. Knowl. Based Syst. 2017, 131, 149–159. [Google Scholar] [CrossRef]
- González, M.; Minuesa, C. del Puerto, I. Maximum Likelihood Estimation and Expectation–Maximization Algorithm for Controlled Branching Processes. Comput. Stat. Data Anal. 2016, 93, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Binkley, D.; Campor, O.C.; Alvares, C.; Carneiro, R.L.; Cegatta, I.; Stape, J.L. The Interactions of Climate, Spacing and Genetics on Clonal Eucalyptus Plantations across Brazil and Uruguay. For. Ecol. Manag. 2017, 405, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.S.; Gonçalves, J.L.M.; Sparovek, G. Koppen’s Climate Classification Map for Brazil. Meteorol. Zeitschrif 2013, 22, 711–728. Available online: http://www.lerf.eco.br/img/publicacoes/Alvares_etal_2014.pdf (accessed on 1 July 2023).
- Elli, E.F.; Sentelhas, P.C.; Freitas, C.H.; Carneiro, R.L.; Alvares, C.A. Intercomparison of Structural Features and Performance of Eucalyptus Simulation Models and Their Ensemble for Yield Estimations. For. Ecol. Manag. 2019, 450, 117493. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Alvares, C.A.; Higac, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; Ferras, S.F.; Lima, W.P.; Brancalion, P.H.S.; Hubner, A.; et al. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Flores, T.B.; Alvares, C.A.; Souza, V.C.; Stape, J.L. Eucalyptus No Brasil: Zonemaneto Climático e Guia Para Identificação; IPEF: Piracicaba, Brazil, 2016.
- Campoe, O.C.; Alvares, C.A.; Carneiro, R.L.; Binkley, D.; Ryan, G.; Hubbard, R.M.; Stahl, J.; Moreira, G.; Moraes, F.; Stape, J.L. Climate and genotype influences on carbon fluxes and partitioning in Eucalyptus plantations. For. Ecol. Manag. 2020, 475, 118445. [Google Scholar] [CrossRef]
- Schumacher, F.X.; Hall, F.S. Logarithmic Expression of Tree Volume. J. Agric. Res. 1933, 47, 719–734. [Google Scholar]
- Ratkowsky, D.A. Handbook of Nonlinear Regression Models; M. Dekker: New York, NY, USA, 1990. [Google Scholar]
- Yin, X.; Martin, J.K.; McLaren, G.; Visperas, R.M. A Nonlinear Model for Crop Development as a Function of Temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.S.; Rubilar, R.; Kimberley, M.O.; Kriticos, D.J.; Emhart, V.; Mardones, O.; Acevedo, M.; Pincheira, M.; Stape, J.; Fox, T. Using Seasonal Measurements to Inform Ecophysiology: Extracting Cardinal Growth Temperatures for Process-Based Growth Models of Five Eucalyptus Species/Crosses from Simple Field Trials. N. Z. J. For. Sci. 2014, 44, 9. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Weinian, Z. Forts of Quadratic Polynomials under Iteration. J. Comput. Appl. Math. 2018, 331, 1–10. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, B.-h.; Hao, Y.; Yang, R.; Wang, Y.-a. Effects of Short-Term Heat Stress on PSII and Subsequent Recovery for Senescent Leaves of Vitis Vinifera, L. Cv. Red Globe. J. Integr. Agric. 2018, 17, 2683–2693. [Google Scholar] [CrossRef]
- Holliday, R. Plant Population and Crop Yield. Nature 1960, 186, 22–24. [Google Scholar] [CrossRef]
- Goudriaan, J.; Monteith, J.L. A Mathematical Function for Crop Growth Based on Light Interception and Leaf Area Expansion. Ann. Bot. 1990, 66, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Charnes, A.; Cooper, W.W.; Rhodes, E. Measuring the Efficiency of Decision Making Units. Eur. J. Oper. Res. 1978, 2, 429–444. [Google Scholar] [CrossRef]
- Nash, J.C.; Ravi, V.; Gabor, G. Expanded Replacement and Extension of the ‘optim’ Function. 2018. Available online: https://cran.rstudio.com/web/packages/optimx/optimx.pdf (accessed on 1 November 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. 2014. Available online: http://www.Rproject.org/ (accessed on 6 June 2022).
- Ocheltree, T.W.; Nippert, J.B.; Prasad, P.V.V. Stomatal Responses to Changes in Vapor Pressure Deficit Reflect Tissue-Specific Differences in Hydraulic Conductance: Stomatal Sensitivity of C3 and C4 Grasses. Plant Cell Environ. 2014, 37, 132–139. [Google Scholar] [CrossRef]
- Hänninen, H.; Kramer, K.; Tanino, K.; Zhang, R.; Wu, J.; Fu, Y.H. Experiments Are Necessary in Process-Based Tree Phenology Modelling. Trends Plant Sci. 2019, 24, 199–209. [Google Scholar] [CrossRef] [PubMed]
- King, G.; Fonti, P.; Nievergelt, D.; Büntgen, U.; Frank, D. Climatic Drivers of Hourly to Yearly Tree Radius Variations along a 6 °C Natural Warming Gradient. Agric. For. Meteorol. 2013, 168, 36–46. [Google Scholar] [CrossRef]
- Rawal, D.S.; Sabine, K.; Keatley, M.R.; Nitschke, C.R. Climatic and Photoperiodic Effects on Flowering Phenology of Select Eucalypts from South-Eastern Australia. Agric. For. Meteorol. 2015, 214–215, 231–242. [Google Scholar] [CrossRef]
- Svystun, T.; Bhalerao, R.P.; Jönsson, A.M. Modelling Populus Autumn Phenology: The Importance of Temperature and Photoperiod. Agric. For. Meteorol. 2019, 271, 346–354. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Ranalli, P. Improvement of Crop Plants for Industrial End Uses; Springer: Dordrecht, The Netherlands, 2007; 533p. [Google Scholar]
- Long, S.P.; Zhu, X.-G.; Naidu, S.L.; Ort, D.R. Can Improvement in Photosynthesis Increase Crop Yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Sands, P.J.; Landsberg, J.J. Parameterisation of 3-PG for Plantation Grown Eucalyptus Globulus. For. Ecol. Manag. 2002, 163, 273–292. [Google Scholar] [CrossRef]
- Caldeira, D.R.M.; Alvares, C.A.; Campoe, O.C.; Hakamada, R.E.; Guerrini, I.A.; Cegatta, I.R.; Stape, J.L. Multisite Evaluation of the 3-PG Model for the Highest Phenotypic Plasticity Eucalyptus Clone in Brazil. For. Ecol. Manag. 2020, 462, 117989. [Google Scholar] [CrossRef]
- Information, E.; Portal, N. Legal Amazon|IBGE. 2022. Available online: https://www.ibge.gov.br/en/geosciences/environmental-information/vegetation/17927-legal-amazon.html?=&t=o-que-e (accessed on 13 April 2022).
- Zoldan, D.; Band, R.S.; Guy, C.L.; Porat, R. Understanding Chilling Tolerance Traits Using Arabidopsis Chilling-Sensitive Mutants. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 159–173. [Google Scholar] [CrossRef]
- Raju, S.K.K.; Barnes, A.C.; Schnable, J.C.; Roston, R.L. Low-Temperature Tolerance in Land Plants: Are Transcript and Membrane Responses Conserved? Plant Sci. 2018, 276, 73–86. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Celestian, S.B.; Martin, C.A. Rhizosphere Surface, and Air Temperature Patterns at Parking Lots in Phoenix, Arizona, U.S. J. Arboric. 2004, 30, 245–252. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | State/Country | Meteorological Station in Situ (S) | Meteorological Station from INMET or INIA * (I) | S ↔ I | ||||
|---|---|---|---|---|---|---|---|---|
| Lat | Long | Alt | Lat | Long | Alt | Distance | ||
| Graus | Meters | Graus | Meters | Kilometers | ||||
| 16 * | Uruguay | −32.2 | −57.8 | 50 | −34.3 | −57.7 | 72 | 112.9 |
| 20 | Sao Paulo—Brazil | −22.4 | −47.0 | 633 | −22.0 | −47.0 | 633 | 17.7 |
| 22 | Parana—Brazil | −24.2 | −50.5 | 888 | −24.0 | −50.0 | 1106 | 31.4 |
| 23 | Santa Catarina—Brazil | −27.5 | −50.1 | 870 | −27.0 | −51.0 | 982 | 53.5 |
| 25 * | Uruguay | −33.3 | −57.9 | 37 | −34.3 | −57.7 | 47 | 103.8 |
| 29 | Maranhao—Brazil | −3.4 | −43.1 | 81 | −4.0 | −43.0 | 91 | 46.7 |
| 30 | Minas Gerais—Brazil | −17.3 | −43.8 | 848 | −17.0 | −44.0 | 646 | 64.6 |
| 31 | Bahia—Brazil | −16.3 | −39.6 | 200 | −16.0 | −39.0 | 88 | 47.6 |
| Model | Name | f(x) | Area | Absolute Error | |||
|---|---|---|---|---|---|---|---|
| Equation (2) | Beta Function | 2.2744 | 37.6508 | 19.9640 | 18.86794 | <6.5 × 10−9 | |
| Equation (3) | Quadratic Polynomial | −1.5923 | 0.2732 | −0.0072 | 15.69885 | <1.7 × 10−13 | |
| Equation (4) | Inverse Quadratic Polynomial | 5.3897 | −0.4481 | 0.0115 | 22.20823 | <2.9 × 10−7 | |
| Equation (5) | Exp Quadratic Polynomial | −2.8590 | 0.3094 | −0.0083 | 19.92385 | <5.9 × 10−5 | |
| Equation (6) | Exp Model | 8.2905 | −15.2678 | 0.4681 | 26.00404 | <1.7 × 10−3 | |
| Equation (7) | Rate Polynomial | 35.5476 | −2.1410 | 0.0773 | 20.36569 | <2.1 × 10−7 |
| Month | 2nd Degree Polynomial | Beta Function | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Area | Absolute Error | Area | Absolute Error | |||||||
| January | −11.5092 | 1.1100 | −0.0246 | 8.4520 | <9.4 × 10−14 | 16.1375 | 23.3863 | 31.4918 | 8.0729 | <4.9 × 10−5 |
| February | −15.7086 | 1.4359 | −0.0309 | 7.5211 | <8.4 × 10−14 | 15.6712 | 23.4399 | 31.7432 | 8.4867 | <2.8 × 10−5 |
| March | −14.8584 | 1.4004 | −0.0309 | 7.5279 | <8.4 × 10−14 | 13.7724 | 21.6694 | 30.0155 | 8.5889 | <2.2 × 10−5 |
| April | −7.1868 | 0.7713 | −0.0182 | 9.8281 | <1.1 × 10−13 | 13.7724 | 21.6694 | 30.0155 | 8.5889 | <2.2 × 10−5 |
| May | −3.3387 | 0.4562 | −0.0120 | 11.9704 | <1.3 × 10−13 | 6.0409 | 18.2299 | 30.9412 | 13.1905 | <2.4 × 10−5 |
| June | −1.9104 | 0.3395 | −0.0099 | 13.3144 | <1.5 × 10−13 | 3.0467 | 16.2665 | 30.0540 | 14.3065 | <2.6 × 10−5 |
| July | −1.9210 | 0.3409 | −0.0100 | 13.3400 | <1.5 × 10−13 | 3.0467 | 16.2665 | 30.0540 | 14.3065 | <2.6 × 10−5 |
| August | −2.0265 | 0.3219 | −0.0086 | 14.2415 | <1.6 × 10−13 | 6.8377 | 18.8862 | 31.0061 | 12.8763 | <1.4 × 10−6 |
| September | −3.5280 | 0.4690 | −0.0122 | 11.8531 | <1.3 × 10−13 | 6.5255 | 17.8706 | 29.5968 | 12.2376 | <1.6 × 10−5 |
| October | −6.0212 | 0.6695 | −0.0160 | 10.4198 | <1.2 × 10−13 | 8.5334 | 22.1008 | 36.9952 | 14.9869 | <7.4 × 10−5 |
| November | −9.3220 | 0.9343 | −0.0212 | 9.0602 | <1 × 10−13 | 13.2544 | 22.8786 | 32.7458 | 10.3517 | <9 × 10−6 |
| December | −7.1279 | 0.7536 | −0.0175 | 10.1607 | <1.1 × 10−13 | 13.8997 | 22.4568 | 31.1807 | 9.1860 | <5.5 × 10−6 |
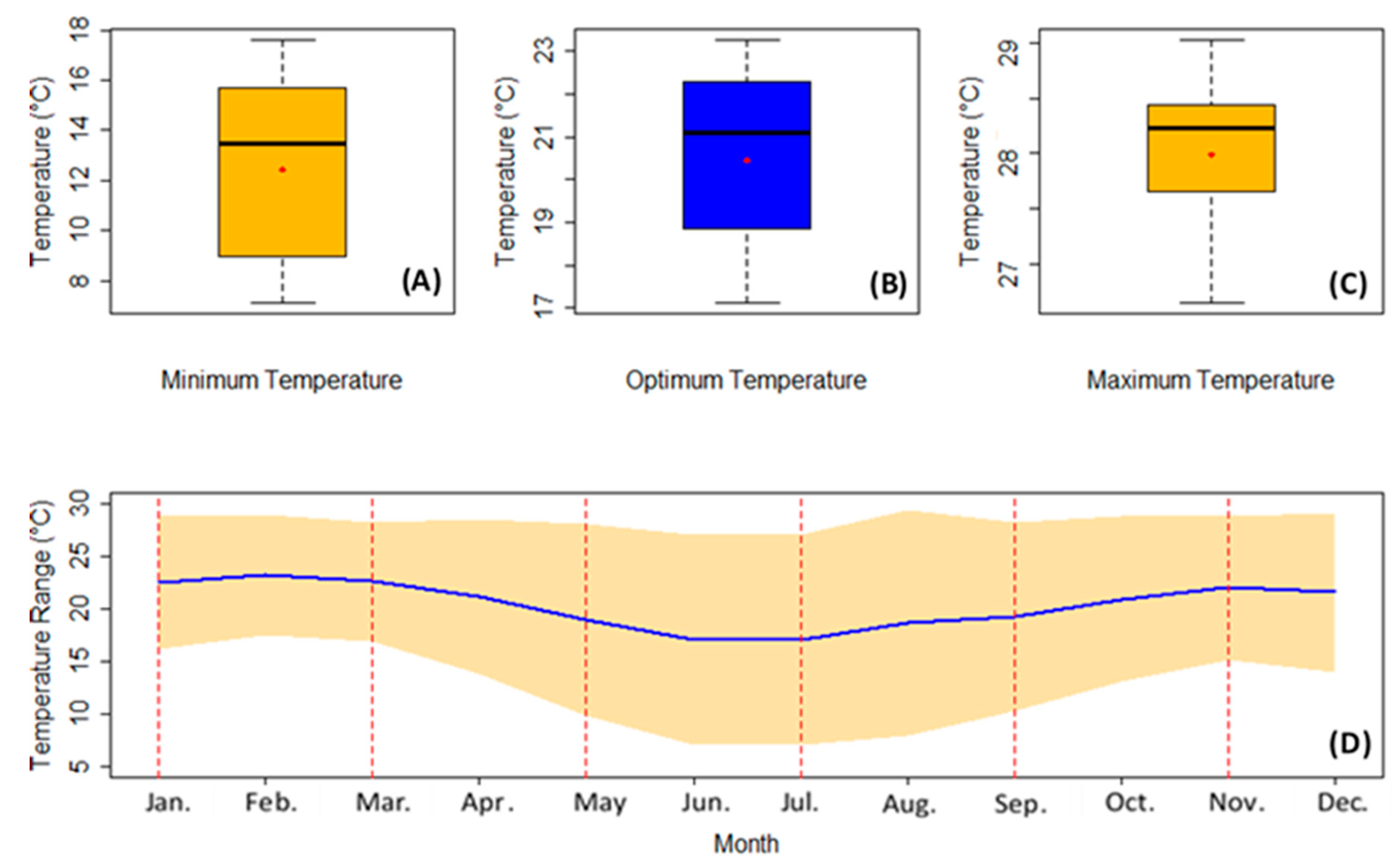
| Minimum Temperature (minT—°C) | Optimum Temperature (optT—°C) | Maximum Temperature (maxT—°C) | |
|---|---|---|---|
| Month | minT | ||
| January | 16.2 | 22.5 | 28.9 |
| February | 17.6 | 23.3 | 28.9 |
| March | 17.0 | 22.7 | 28.3 |
| April | 13.8 | 21.2 | 28.6 |
| May | 9.9 | 19.0 | 28.1 |
| June | 7.1 | 17.1 | 27.2 |
| July | 7.1 | 17.1 | 27.2 |
| August | 8.0 | 18.8 | 29.5 |
| September | 10.3 | 19.3 | 28.3 |
| October | 13.1 | 21.0 | 28.8 |
| November | 15.2 | 22.1 | 28.9 |
| December | 14.0 | 21.6 | 29.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, T.B.; Montes, C.R.; Campoe, O.C. A New Method to Calibrate Cardinal Temperatures for Eucalyptus Plantation. Forests 2023, 14, 1631. https://doi.org/10.3390/f14081631
Queiroz TB, Montes CR, Campoe OC. A New Method to Calibrate Cardinal Temperatures for Eucalyptus Plantation. Forests. 2023; 14(8):1631. https://doi.org/10.3390/f14081631
Chicago/Turabian StyleQueiroz, Túlio Barroso, Cristian Rodrigo Montes, and Otávio Camargo Campoe. 2023. "A New Method to Calibrate Cardinal Temperatures for Eucalyptus Plantation" Forests 14, no. 8: 1631. https://doi.org/10.3390/f14081631