
The Mechanical Stability of Pure Norway Spruce Stands along an Altitudinal Gradient in the Czech Republic

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Source
- (1)
- nutrient-rich—this represents mesotrophic soils without a significant soil water influence.
- (2)
- acidic—this represents oligotrophic soils with no significant soil water effect.
2.2. Aggregation of Tree-Level Data
2.3. HDR-Age Relationship Modelling
2.4. Modelling HDR–Age Relationship by Ecological Series (ES)
2.5. CR–Age Relationship Modelling
3. Results
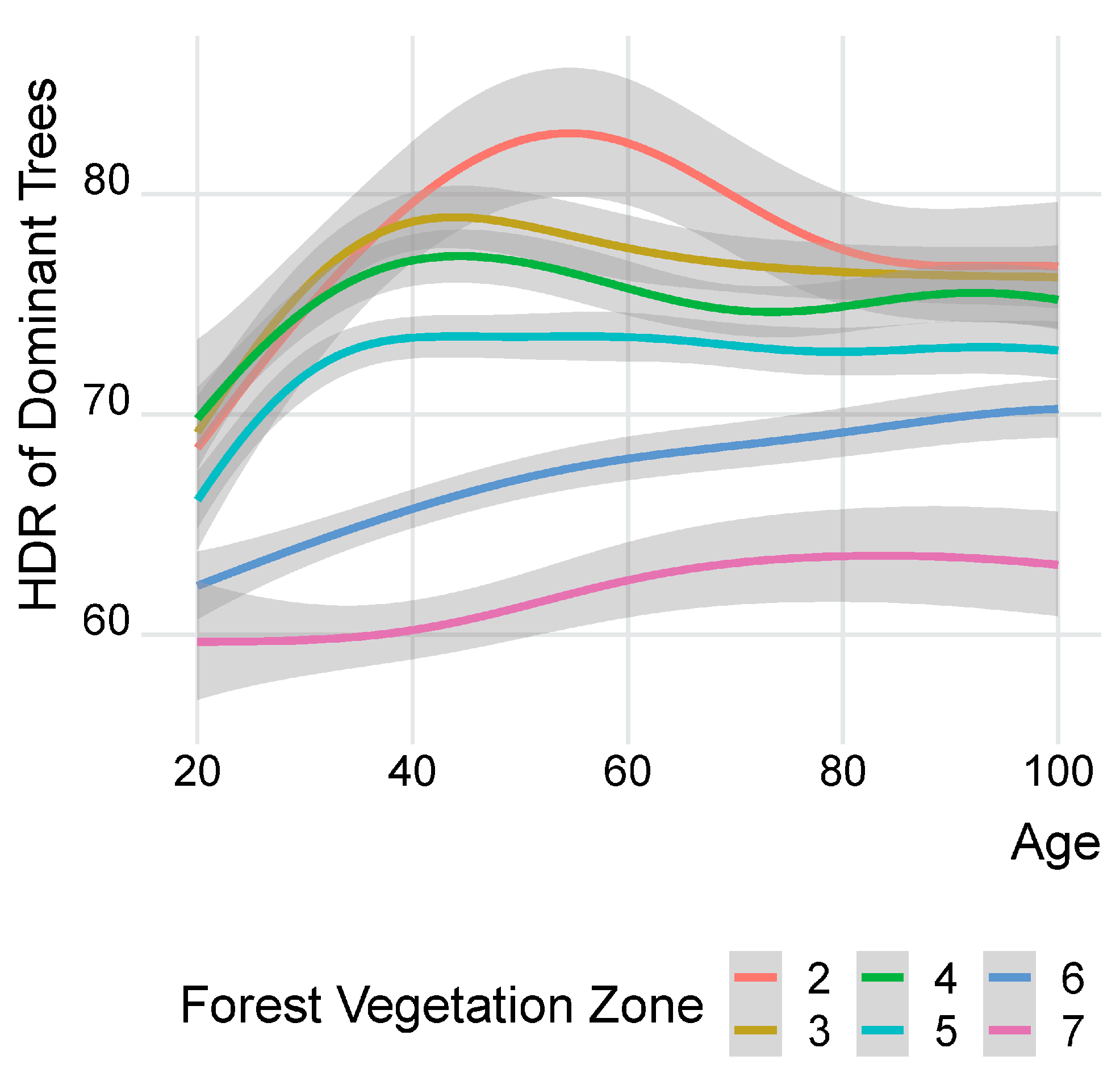
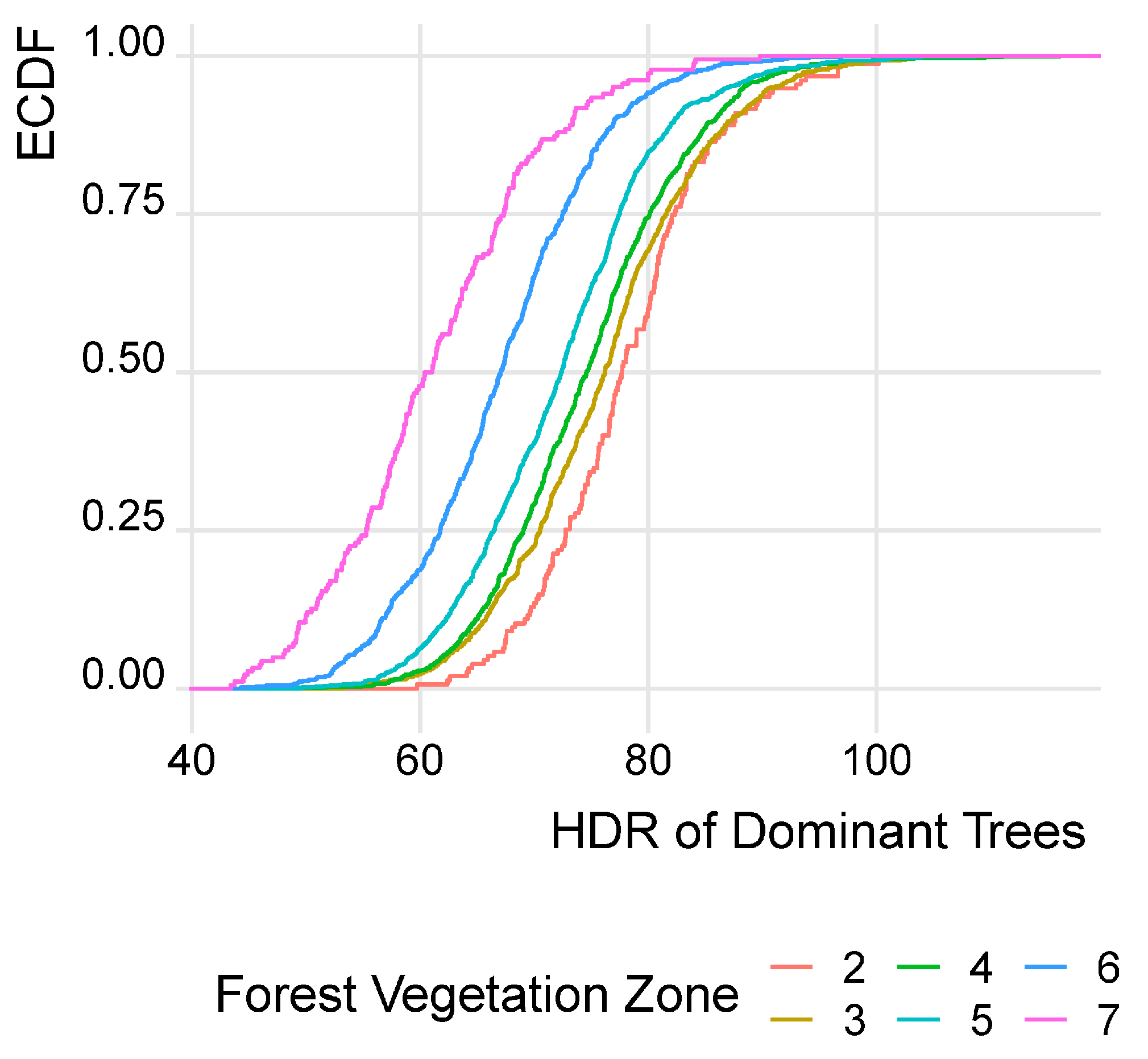
3.1. Development of Dominant Trees’ HDR by Forest Vegetation Zones (FVZ)
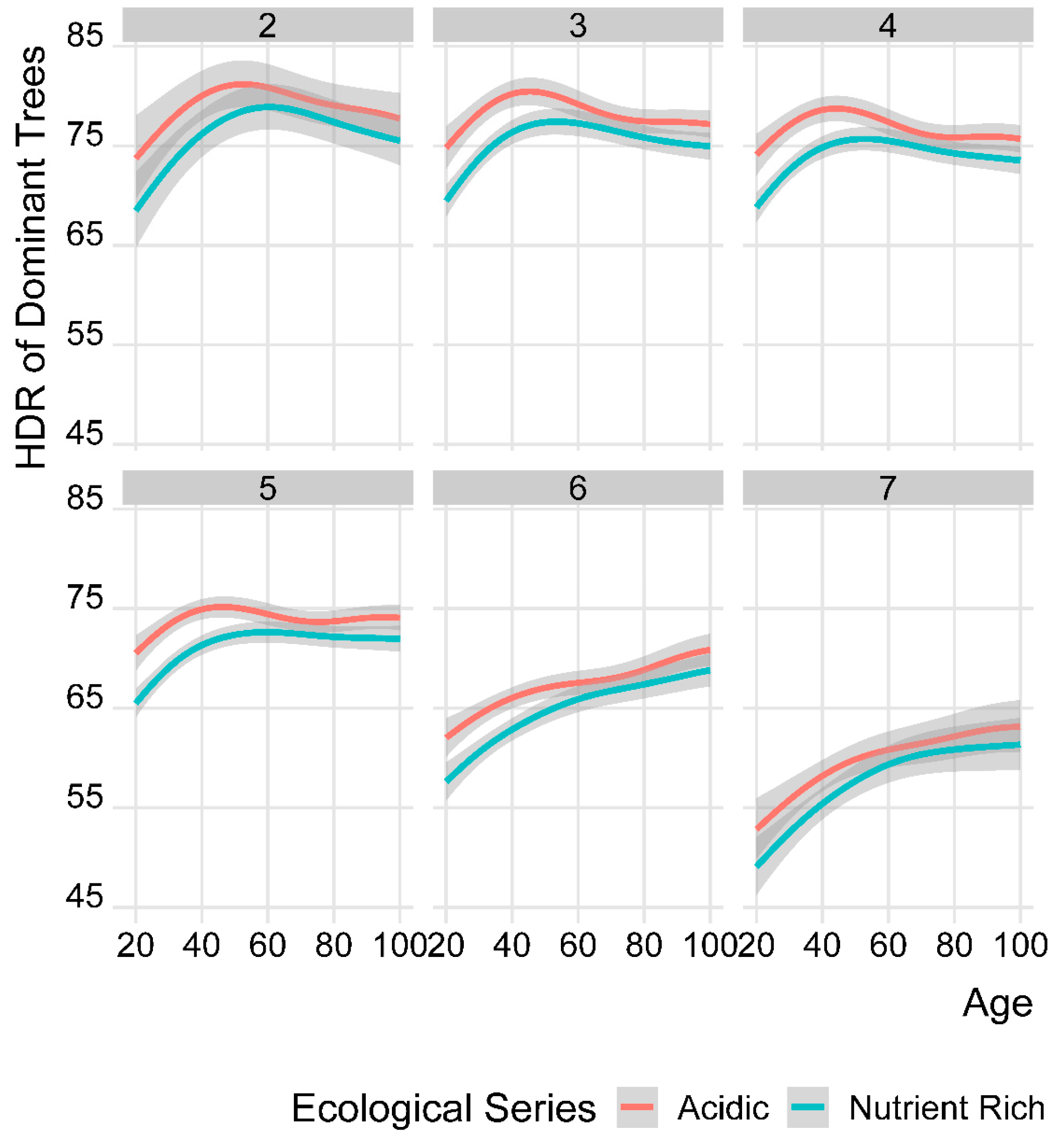
3.2. Development of Dominant Trees’ HDR in Acidic and Nutrient-Rich Ecological Series (ES)
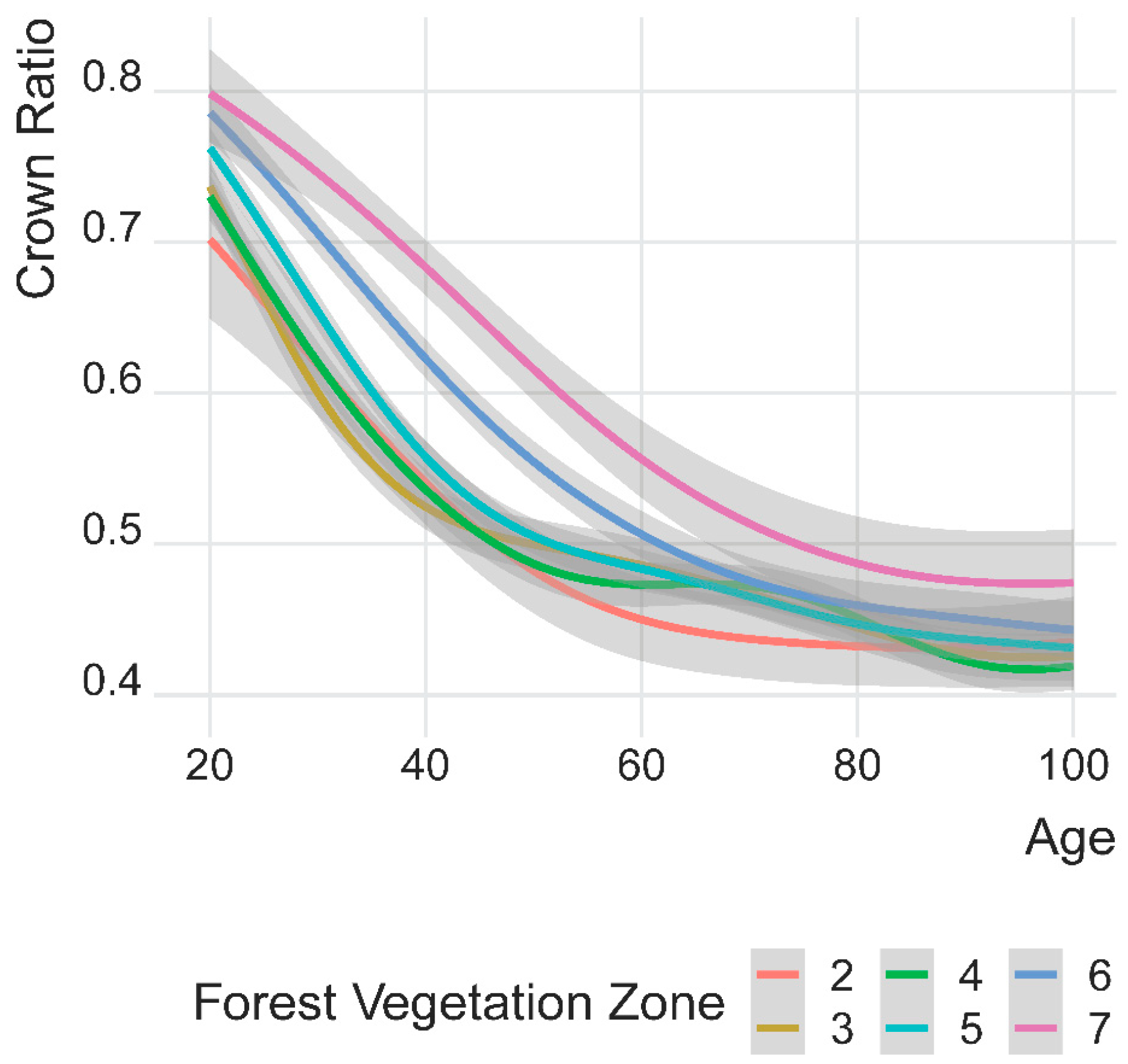
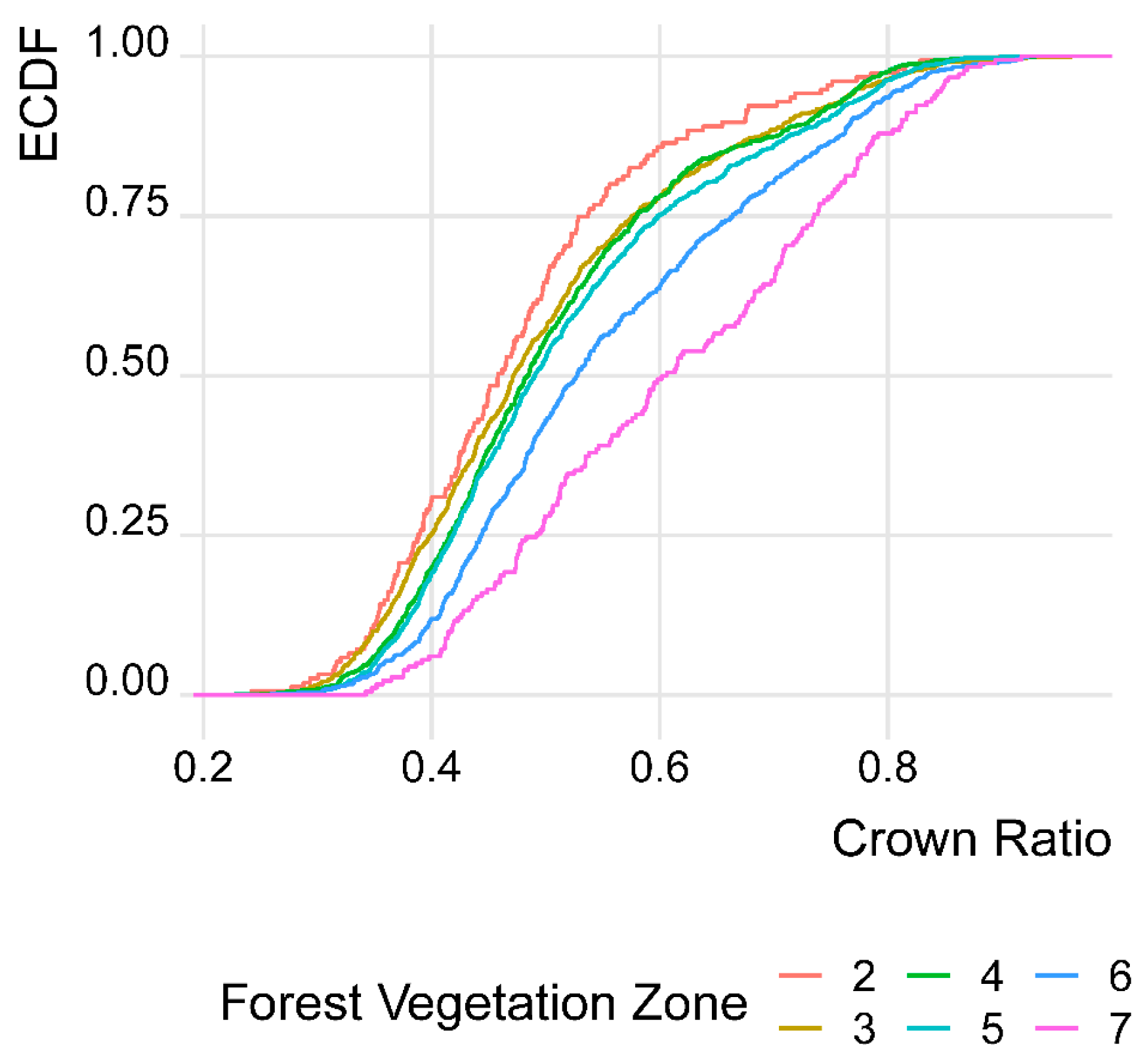
3.3. Development of Mean Crown Ratio (CR) by Forest Vegetation Zones (FVZs)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kohler, M.; Sohn, J.; Nägele, G.; Bauhus, J. Can drought tolerance of Norway spruce (Picea abies (L.) Karst.) be increased through thinning. Eur. J. For. Res. 2010, 129, 1109–1118. [Google Scholar] [CrossRef]
- Hlásny, T.; Křístek, Š.; Holuša, J.; Trombik, J.; Urbaňcová, N. Snow disturbances in secondary Norway spruce forests in Central Europe: Regression modeling and its implications for forest management. For. Ecol. Manag. 2011, 262, 2151–2161. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Change 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Steckel, M.; del Río, M.; Heym, M.; Aldea, J.; Bielak, K.; Brazaitis, G.; Černý, J.; Coll, L.; Collet, C.; Ehbrecht, M.; et al. Species mixing reduces drought susceptibility of Scots pine (Pinus sylvestris L.) and oak (Quercus robur L.; Quercus petraea (Matt.) Liebl.)—Site water supply and fertility modify the mixing effect. For. Ecol. Manag. 2020, 461, 117908. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Hummel, S.; Albrecht, A. Assessing natural hazards in forestry for risk management: A review. Eur. J. For. Res. 2011, 130, 329–351. [Google Scholar] [CrossRef]
- Cukor, J.; Vacek, Z.; Linda, R.; Sharma, R.P.; Vacek, S. Afforested farmland vs. forestland: Effects of bark stripping by Cervus elaphus and climate on production potential and structure of Picea abies forests. PLoS ONE 2019, 14, e0221082. [Google Scholar] [CrossRef] [Green Version]
- Vacek, Z.; Cukor, J.; Linda, R.; Vacek, S.; Šimůnek, V.; Brichta, J.; Gallo, J.; Prokůpková, A. Bark stripping, the crucial factor affecting stem rot development and timber production of Norway spruce forests in Central Europe. For. Ecol. Manag. 2020, 474, 118360. [Google Scholar] [CrossRef]
- Cavin, L.; Mountford, E.P.; Peterken, G.F.; Jump, A.S. Extreme drought alters competitive dominance within and between tree species in a mixed forest stand. Funct. Ecol. 2013, 27, 1424–1435. [Google Scholar] [CrossRef]
- Bottero, A.; Forrester, D.I.; Cailleret, M.; Kohnle, U.; Gessler, A.; Michel, D.; Bose, A.K.; Bauhus, J.; Bugmann, H.; Cuntz, M.; et al. Growth resistance and resilience of mixed silver fir and Norway spruce forests in central Europe: Contrasting responses to mild and severe droughts. Glob. Change Biol. 2021, 27, 4403–4419. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistant carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suvanto, S.; Peltoniemi, M.; Tuominen, S.; Strandström, M.; Lehtonen, A. High-resolution mapping of forest vulnerability to wind for disturbance-aware forestry. For. Ecol. Manag. 2019, 453, 117619. [Google Scholar] [CrossRef]
- Gardiner, B. Wind damage to forests and trees: A review with an emphasis on planted and managed forests. J. For. Res. 2021, 26, 248–266. [Google Scholar] [CrossRef]
- Łaska, G. The disturbance and vegetation dynamics: A review and an alternative framework. Plant Ecol. 2001, 157, 77–99. [Google Scholar] [CrossRef]
- Tsvetanov, M.; Dountchev, A.; Panayotov, M.; Zhelev, P.; Bebi, P.; Yurukov, S. Short- and long-term natural regeneration after windthrow disturbances in Norway spruce forests in Bulgaria. iForest 2018, 11, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Forzieri, G.; Girardello, M.; Ceccherini, G.; Spinoni, J.; Feyen, L.; Hartmann, H.; Beck, P.S.A.; Camps-Valls, G.; Chirici, G.; Mauri, A.; et al. Emergent vulnerability to climate-driven disturbances in European forests. Nat. Commun. 2021, 12, 1081. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R. Mapping the forest disturbance regimes of Europe. Nat. Sustain. 2021, 4, 63–70. [Google Scholar] [CrossRef]
- Schuck, A.; Schelhaas, M.-J. Storm damage in Europe—An overview. In Living with Storm Damage to Forests; Gardiner, B., Schuck, A., Schelhaas, M.-J., Orazio, C., Blennow, K., Nicoll, B., Eds.; European Forest Institute: Joensu, Finland, 2013; pp. 15–23. [Google Scholar]
- MZe. Report about Stage of Forests and Forest Management in the Czech Republic in 2021; Ministry of Agriculture of the Czech Republic: Praha, Czech Republic, 2021; p. 47. (In Czech) [Google Scholar]
- Forzieri, G.; Pecchi, M.; Girardello, M.; Mauri, A.; Klaus, M.; Nikolov, C.; Rüetschi, M.; Gardiner, B.; Tomaštík, J.; Small, D.; et al. A spatially explicit database of wind disturbances in European forests over the period 2000–2018. Earth Syst. Sci. Data 2020, 12, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Change 2012, 3, 203–207. [Google Scholar] [CrossRef]
- Konôpka, B.; Zach, P.; Kulfan, J. Wind—An important ecological factor and destructive agent in forests. For. J. 2016, 62, 123–130. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Change Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Change 2014, 4, 806–810. [Google Scholar] [CrossRef] [Green Version]
- Zell, J.; Hanewinkel, M. How treatment, storm events and changed climate affect productivity of temperate forests in SW Germany. Reg. Environ. Change 2015, 15, 1531–1542. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. Camb. Phylos. Soc. 2016, 91, 760–781. [Google Scholar] [CrossRef]
- Reyer, C.P.O.; Bathgate, S.; Blennow, K.; Borges, J.G.; Bugmann, H.; Delzon, S.; Faias, S.P.; Garcia-Gonzalo, J.; Gardiner, B.; Gonzalez-Olabarria, J.R.; et al. Are forest disturbances amplifying or canceling out climate change-induced productivity changes in European forests? Environ. Res. Lett. 2017, 12, 034027. [Google Scholar] [CrossRef]
- Díaz-Yáñez, O.; Mola-Yudego, B.; Eriksen, R.; González-Olabarria, J.R.; Carcaillet, C. Assessment of the main natural disturbances on Norwegian forest based on 20 years of national inventory. PLoS ONE 2016, 11, e0161361. [Google Scholar] [CrossRef] [Green Version]
- Dodan, M.; Perić, S. Windthrow resistance of Norway spruce (Picea abies L. Karst.) forest cultures—preliminary results. Seefor-South-East Eur. For. 2019, 10, 77–88. [Google Scholar] [CrossRef]
- Gardiner, B.; Blennow, K.; Carnus, J.-M.; Fleischer, P.; Ingemarson, F.; Landmann, G.; Linder, M.; Marzano, M.; Nicoll, B.; Orazio, C.; et al. Destructive Storms in European Forests: Past and Forthcoming Impacts; Final report to European Commission—DG Environment: Brussels, Belgium, 2010; p. 138. [Google Scholar] [CrossRef]
- Slodičák, M.; Novák, J. Silvicultural measures to increase the mechanical stability of pure secondary Norway spruce stands before conversion. For. Ecol. Manag. 2006, 224, 252–257. [Google Scholar] [CrossRef]
- del Río, M.; Bravo-Oviedo, A.; Pretzsch, H.; Löf, M.; Ruiz-Peinado, R. A review of thinning effects on Scots pine stands: From growth and yield to new challenges under global change. For. Syst. 2017, 26, eR03S. [Google Scholar] [CrossRef]
- Vacek, Z.; Prokůpková, A.; Vacek, S.; Cukor, J.; Bílek, L.; Gallo, J.; Bulušek, D. Silviculture as a tool to support stability and diversity of forests under climate change: Study from Krkonoše Mountains. Cent. Eur. For. J. 2020, 66, 116–129. [Google Scholar] [CrossRef]
- Romeiro, J.M.N.; Eid, T.; Anton-Fernandez, C.; Kangas, A.; Tromborg, E. Natural disturbances risks in European boreal and temperate forests and their links to climate change—A review of modelling approaches. For. Ecol. Manag. 2022, 509, 120071. [Google Scholar] [CrossRef]
- Pařez, J. Škody Sněhem a Větrem ve Smrkových Porostech Probírkových Pokusných Ploch v Období 1959–1968 [Damages Caused by Snow and Wind in Pure Norway Spruce Stands at Silvicultural Experiments in the Period of 1959–1968]; VÚLHM: Jíloviště-Strnady, Czech Republic, 1972; p. 97. (In Czech) [Google Scholar]
- Chroust, L. Tvar kmene a velikost korun při výchově smrkových porostů ve vztahu ke škodám působeným sněhem a větrem [Stem shape and crown size in during Norway spruce tending related to damages elicited by snow and wind]. Práce VÚLHM 1980, 56, 31–52. (In Czech) [Google Scholar]
- Slodičák, M. Resistance of young spruce stands to snow and wind damage in dependence on thinning. Commun. Inst. For. Cech. 1987, 15, 75–86. [Google Scholar]
- Nykänen, M.L.; Peltola, H.; Quine, C.; Kellomäki, S.; Broadgate, M. Factors affecting snow damage of trees with particular reference to European conditions. Silva Fenn. 1997, 31, 193–213. [Google Scholar]
- Mason, B.; Valinger, E. Managing forests to reduce storm damage. In Living with Storm Damage to Forests. What Science Can Tell Us; Gardiner, B., Schuck, A., Schelhaas, M.-J., Orazio, C., Blennow, K., Nicoll, B., Eds.; European Forest Institute: Joensuu, Finland, 2013; pp. 87–96. [Google Scholar]
- Vacchiano, G.; Derose, R.J.; Shaw, J.D.; Svoboda, M.; Motta, R. A density management diagram for Norway spruce in the temperate European montane region. Eur. J. For. Res. 2013, 132, 535–549. [Google Scholar] [CrossRef]
- Gauthier, M.-M.; Tremblay, S. Precommercial thinning as a silvicultural option for treating very dense conifer stands. Scand. J. For. Res. 2018, 33, 446–454. [Google Scholar] [CrossRef]
- Dušek, D.; Novák, J.; Kacálek, D.; Slodičák, M. Norway spruce production and static stability in IUFRO thinning experiments in the Czech Republic. J. For. Sci. 2021, 67, 185–194. [Google Scholar] [CrossRef]
- Bragg, D.C.; Shelton, M.G.; Zeide, B. Impacts and management implications of ice storms on forests in the southern United States. For. Ecol. Manag. 2003, 186, 99–123. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Kenefic, L.S.; Seymour, R.S.; Phillips, L.M. Long-term effects of precommercial thinning on the stem dimensions, form and branch characteristics of red spruce and balsam fir crop trees in Maine, USA. Silva Fenn. 2009, 43, 397–409. [Google Scholar]
- Zhang, X.; Wang, H.; Cchin, S.; Zhang, J. Effects of competition, age and climate on tree slenderness of Chinese fir plantations in southern China. For. Ecol. Manag. 2020, 458, 117815. [Google Scholar] [CrossRef]
- Swift, D.E.; Knight, W.; Béland, M.; Boureima, I.; Bourque, C.P.-A.; Meng, F.R. Stand dynamics and tree quality response to precommercial thinning in a northern hardwood forest of the Acadian forest region: 23 years of intermediate results. Scand. J. For. Res. 2017, 32, 45–59. [Google Scholar] [CrossRef]
- Klinar, B.; Klopčič, M.; Bončina, A. Individual tree damage due to abiotic natural disturbances on European beech sites in Slovenia with the main focus on snow damage. Acta Silvae Et. Ligni 2020, 122, 53–69, (in Slovenian with English abstract and summary). [Google Scholar] [CrossRef]
- Brüchert, F.; Becker, G.; Speck, T. The mechanics of Norway spruce (Picea abies L. Karst) mechanical properties of standing trees from different thinning regimes. For. Ecol. Manag. 2000, 135, 45–62. [Google Scholar] [CrossRef]
- Seifert, T.; Pretzsch, H.; Bücking, M. Coppice with spruce from high forest? Part II: Year ring width, stem taper and branchiness of long crowned Norway spruce. Forst. Holz. 2003, 59, 63–68. [Google Scholar]
- Pretzsch, H. Tree growth as affected by stem and crown structure. Trees 2021, 35, 947–960. [Google Scholar] [CrossRef]
- Katrevičs, J.; Džeriņa, B.; Neimane, U.; Desaine, I.; Bigača, Z.; Jansons, A. Production and profitability of low density Norway spruce (Picea abies L. Karst.) plantation at 50 years age: Case study from eastern Latvia. Agron. Res. 2018, 16, 113–121. [Google Scholar] [CrossRef]
- Albrecht, A.; Hanewinkel, M.; Bauhus, J.; Kohnle, U. How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modelling based on long-term observations. Eur. J. For. Res. 2012, 131, 229–247. [Google Scholar] [CrossRef]
- Piticar, M.A. The effects of thinning on a spruce stand, over a period of 40 years of monitoring. Adv. Agric. Bot. 2016, 8, 77–86. [Google Scholar]
- Peltola, H.; Kellomaki, S.; Hassinen, A.; Granander, M. Mechanical stability of Scots pine, Norway spruce and birch: An analysis of tree-pulling experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Rust, S. Analysis of regional variation of height growth and slenderness in populations of six urban tree species using a quantile regression approach. Urban For. Urban Green. 2014, 13, 336–343. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S.; Kučera, M. A nonlinear mixed-effects height-to-diameter ratio model or several tree species based on Czech National Forest Inventory data. Forests 2019, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Jiang, L.; Wang, J. The influence of climate, soil physicochemical properties and tree size inequality on tree slenderness in mixed forests of Northeastern China. For. Ecol. Manag. 2023, 529, 120719. [Google Scholar] [CrossRef]
- Cremer, K.W.; Borough, C.J.; McKinnell, F.H.; Carter, P.R. Effects of stocking and thinning on wind damage in plantations. New Zealand J. For. Sci. 1982, 12, 224–268. [Google Scholar]
- Bayar, E.; Deligoz, A. Effects of precommercial thinning on growth parameters in Pinus nigra Arn. subsp. Pallasiana (Lamb.) Holmboe Stand. J. For. Res. 2020, 31, 1803–1811. [Google Scholar] [CrossRef]
- Slodičák, M.; Novák, J.; Dušek, D. Management of Norway spruce stands in the Western Carpathians. In The Carpathians: Integrating Nature and Society towards Sustainability. Environmental Science and Engineering; Kozak, J., Ed.; Springer: Berlin, Heidelberg, 2013. [Google Scholar] [CrossRef]
- Jelonek, T.; Walkowiak, R.; Jakubowski, M.; Tomczak, A. Indices of tree stability in Scots pine stands damaged by wind. Sylwan 2013, 157, 323–329. [Google Scholar]
- Slodičák, M.; Novák, J. Růst, Struktura a Statická Stabilita Smrkových Porostů s Různým Režimem Výchovy. [Growth, Structure and Static Stability of Norway Spruce Stands with Different Thinning Regimes]; Folia Forestalia Bohemica 3; Lesnická Práce: Kostelec nad Černými lesy, Czech, 2006; p. 128. ISBN 978-80-86386-91-1. (In Czech) [Google Scholar]
- Vospernik, S.; Monserud, R.A.; Sterba, H. Do individual-tree growth models correctly represent height:diameter ratios of Norway spruce and Scots pine? For. Ecol. Manag. 2010, 260, 1735–1753. [Google Scholar] [CrossRef] [Green Version]
- Wonn, H.T.; O’Hara, K.L. Height:diameter ratios and stability relationships for four Northern Rocky Mountain tree species. West. J. Appl. For. 2001, 16, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Bošeľa, M.; Konôpka, B.; Šebeň, V.; Vladovič, J.; Tobin, B. Modelling height to diameter ratio—An opportunity to increase Norway spruce stand stability in the Western Carpathians. For. J. 2014, 60, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devasting outbreak of bark beetle in the Czech Republic: Drivers, impacts, and management implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Albrecht, A.; Kohnle, U.; Hanewinkel, M.; Bauhus, J. Storm damage of Douglas-fir unexpectedly high compared to Norway spruce. Ann. For. Sci. 2013, 70, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; König, L.; Kroneke, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark beetle outbreaks in Europe: State of knowledge and ways forward for management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- FMI. National Forest Inventory in the Czech Republic 2001–2004; Forest Management Institute: Brandýs nad Labem, Czech Republic, 2007; p. 224. (In Czech) [Google Scholar]
- Viewegh, J.; Kusbach, A.; Mikeska, M. Czech forest ecosystem classification. J. For. Sci. 2003, 49, 74–82. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 12 April 2023).
- Levakovic, A. Analytical form of growth laws. Glas. Za Sumske Pokuse 1935, 4, 189–282. [Google Scholar]
- Wood, S. Generalized Additive Models, 2nd ed.; Chapman & Hall/CRC: New York, NY, USA, 2017; p. 476. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, E.J.; Miller, D.L.; Simpson, G.L.; Ross, N. Hierarchical generalized additive models in ecology: An introduction with mgcv. PeerJ 2019, 7, e6876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S. Thin plate regression splines. J. R. Stat. Society. Ser. B 2003, 65, 95–114. [Google Scholar] [CrossRef]
- Faraway, J. Extending the Linear Model with R, 2nd ed.; Chapman & Hall/CRC: New York, NY, USA, 2016; p. 413. [Google Scholar] [CrossRef]
- Harezlak, J.; Ruppert, D.; Wand, M.P. Semiparametric Regression with R; Springer: New York, NY, USA, 2018; p. 331. [Google Scholar] [CrossRef]
- Rigby, R.A.; Stasinopoulos, M.D.; Heller, G.Z.; Bastiani, F.D. Distributions for Modeling Location, Scale, and Shape Using GAMLSS in R; Chapman & Hall/CRC: New York, NY, USA, 2020; p. 588. [Google Scholar] [CrossRef]
- Gschwantner, T.; Lanz, A.; Vidal, C.; Bošeľa, M.; Cosmo, L.; Fridman, J.; Gasparini, P.; Kuliesis, A.; Tomter, S.; Schadauer, K. Comparison of methods used in European National Forest Inventories for the estimation of volume increment: Towards harmonisation. Ann. For. Sci. 2016, 73, 807–821. [Google Scholar] [CrossRef] [Green Version]
- Chirici, G.; McRoberts, R.E.; Winter, S.; Bertini, R.; Brandli, U.-B.; Asensio, I.A.; Bastrup-Birk, A.; Rondeaux, J.; Barsoum, N.; Marchetti, M. National Forest Inventory contributions to forest biodiversity monitoring. For. Sci. 2012, 58, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, I.; Nunes, L.; Kovac, M.; Bonheme, I.; Cañellas, I.; Rego, F.C.; Dias, S.; Duarte, I.; Notarangelo, M.; Rizzo, M.; et al. The conservation status assessment of Natura 2000 forest habitats in Europe: Capabilities, potentials and challenges of national forest inventories data. Ann. For. Sci. 2019, 76, 34. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, M.A.; Edwards, D.M.; Jensen, F.S.; van der Jagt, A.P.N.; Ditchburn, B.R.; Sievanen, T.; Gasparini, P. Harmonising, improving and using social and recreational data in National Forest Inventories across Europe. Ann. For. Sci. 2020, 77, 76. [Google Scholar] [CrossRef]
- Šebeň, V.; Bošeľa, M.; Konôpka, B.; Pajtík, J. Indices of tree competition in dense spruces stand originated from natural regeneration. Lesn. Časopis For. J. 2013, 59, 172–179. [Google Scholar]
- Konôpka, B.; Konôpka, J. Static stability of forest stands in the seventh altitudinal vegetation zone in Slovakia. J. For. Sci. 2003, 49, 474–481. [Google Scholar]
- Homeier, J.; Breckle, S.W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich Ecuadorian Montane rain forest. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Lines, E.R.; Zavala, M.A.; Purves, D.W.; Coomes, D.A. Predictable changes in aboveground allometry of trees along gradients of temperature, aridity and competition. Glob. Ecol. Biogeogr. 2012, 21, 1017–1028. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Liu, S.; Zhang, Y.; Li, J. Variation in height-diameter allometry of ponderosa pine along competition, climate, and species diversity gradients in the western United States. For. Ecol. Manag. 2021, 497, 119477. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Xia, R. Using random forest to disentangle the effects of environmental conditions on height-to-diameter ratio of Engelmann spruce. New For. 2023. (online first). [Google Scholar] [CrossRef]
- Černý, J.; Pokorný, R.; Vejpustková, M.; Šrámek, V.; Bednář, P. Air temperature is the main driving factor of radiation use efficiency and carbon storage of mature Norway spruce stands under global climate change. Int. J. Biometeorol. 2020, 64, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Orman, O.; Adamus, M.; Foremník, K. Norway spruce sapling plasticity in their response of architecture and growth to light gradient decreases with altitude in subalpine stands. For. Ecol. Manag. 2023, 536, 120898. [Google Scholar] [CrossRef]
- Bošeľa, M.; Máliš, F.; Kulla, L.; Šebeň, V.; Deckmyn, G. Ecologically based height growth model and derived raster maps of Norway spruce site index in the Western Carpathians. Eur. J. For. Res. 2013, 132, 691–705. [Google Scholar] [CrossRef]
- Konôpka, J.; Konôpka, B. Static stability of spruce stands subjected to target trees method tending. Rep. For. Res. 2020, 65, 82–92. [Google Scholar]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbertin, M. Influence of stand structure and site factors on wind damage comparing the storms Vivian and Lothar. For. Snow Landsc. Res. 2002, 77, 187–205. [Google Scholar]
- Bouchard, M.; Pothier, D.; Ruel, J.-C. Stand-replacing windthrow in the boreal forests of eastern Quebec. Can. J. For. Res. 2009, 39, 481–487. [Google Scholar] [CrossRef]
- Zeide, B.; Vanderschaaf, C. The effect of density on the height-diameter relationship. In Proceedings of the Eleventh Biennial Southern Silvicultural Research Conference; Outcalt, K.W., Ed.; General Technical Report SRS-48; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 463–466. [Google Scholar]
- Valinger, E.; Fridman, J. Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. For. Ecol. Manag. 2011, 262, 398–403. [Google Scholar] [CrossRef]
- Brüchert, F.; Gardiner, B. The effect of wind exposure on the tree aerial architecture and biomechanics of Sitka spruce (Picea sitchensis, Pinaceae). Am. J. Bot. 2006, 93, 1512–1521. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Huuskonen, S.; Hynynen, J.; Siipilehto, J.; Niemistö, P. Tree-level differences in Norway spruce and Scots pine growth after extreme thinning treatments. Scand. J. For. Res. 2022, 31, 109–118. [Google Scholar] [CrossRef]
- Yousefpour, R.; Hanewinkel, M.; Le Moguédec, G. Evaluating the stability of management strategies of pure Norway spruce forests in the Black Forest area of southwest Germany for adaptation to or mitigation of climate change. Environ. Manag. 2010, 45, 387–402. [Google Scholar] [CrossRef]
- Pasztor, F.; Matulla, C.; Zuvela-Aloise, M.; Rammer, W.; Lexer, M.J. Developing predictive models of wind damage in Austrian forests. Ann. For. Sci. 2014, 72, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Steckel, M.; Heym, M.; Biber, P.; Ammer, C.; Ehbrecht, M.; Bielak, K.; Bravo, F.; Ordóñez, C.; Collet, C.; et al. Stand growth and structure of mixed-species and monospecific stands of Scots pine (Pinus sylvestris L.) and oak (Q. robur L.; Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2020, 139, 349–367. [Google Scholar] [CrossRef] [Green Version]
- Vospernik, S.; Heym, M.; Pretzsch, H.; Pach, M.; Steckel, M.; Aldea, J.; Brazaitis, G.; Bravo-Oviedo, A.; del Río, M.; Löf, M.; et al. Tree species growth response to climate in mixtures of Quercus robur/Quercus petraea and Pinus sylvestris across Europe—A dynamic, sensitive equilibrium. For. Ecol. Manag. 2023, 530, 120753. [Google Scholar] [CrossRef]
- del Río, M.; Pretzsch, H.; Ruiz-Peinado, R.; Jactel, H.; Coll, L.; Löf, M.; Aldea, J.; Ammer, C.; Avdagić, A.; Barbeito, I.; et al. Emerging stability of forest productivity by mixing two species buffers temperature destabilizing effect. J. Appl. Ecol. 2022, 59, 2730–2741. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Vegetation Zone (FVZ) | Mean Annual Air Temperature (°C) | Mean Sum of Annual Precipitation (mm) | Growing Season Length (Days) |
|---|---|---|---|
| 2nd Beech—Oak zone | 7.5–8.0 | 600–650 | 160–165 |
| 3rd Oak—Beech zone | 6.5–7.5 | 650–700 | 150–160 |
| 4th—Beech zone | 6.0–6.5 | 700–800 | 140–150 |
| 5th Fir—Beech zone | 5.5–6.0 | 800–900 | 130–140 |
| 6th Spruce—Beech zone | 4.5–5.5 | 900–1050 | 115–130 |
| 7th Beech—Spruce zone | 4.0–4.5 | 1050–1200 | 100–115 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dušek, D.; Novák, J.; Černý, J. The Mechanical Stability of Pure Norway Spruce Stands along an Altitudinal Gradient in the Czech Republic. Forests 2023, 14, 1558. https://doi.org/10.3390/f14081558
Dušek D, Novák J, Černý J. The Mechanical Stability of Pure Norway Spruce Stands along an Altitudinal Gradient in the Czech Republic. Forests. 2023; 14(8):1558. https://doi.org/10.3390/f14081558
Chicago/Turabian StyleDušek, David, Jiří Novák, and Jakub Černý. 2023. "The Mechanical Stability of Pure Norway Spruce Stands along an Altitudinal Gradient in the Czech Republic" Forests 14, no. 8: 1558. https://doi.org/10.3390/f14081558