
Afforestation Alters the Molecular Composition of Soil Organic Matter in the Central Loess Plateau of China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection
2.3. Soil Organic Matter Biomarker Extraction and Analysis
2.4. Solid-State Nuclear Magnetic Resonance Analysis
2.5. Biomarker Parameters and Calculations
2.6. Statistical Analyses
3. Results
3.1. MBC and SOC Concentrations under Four Treatments
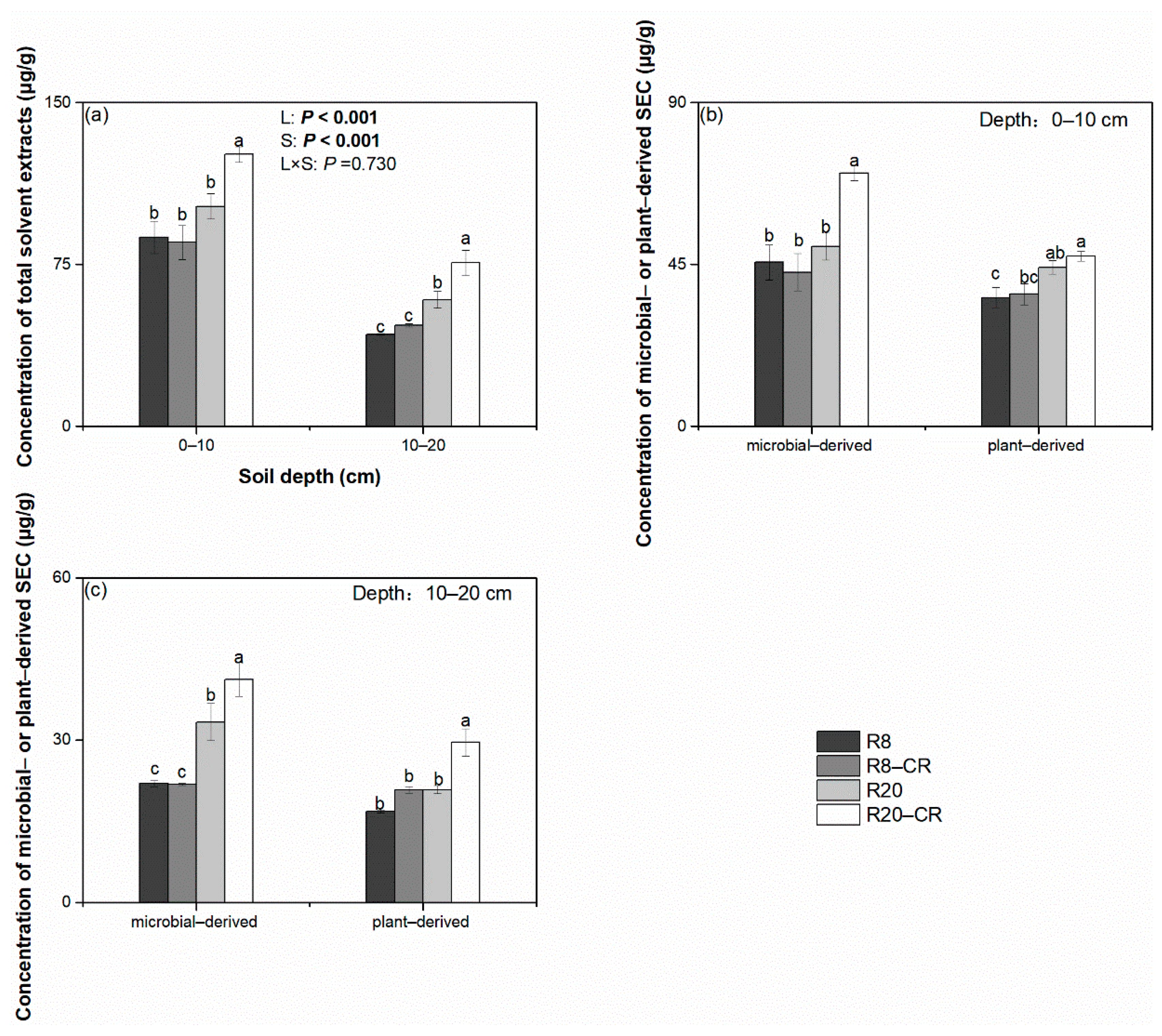
3.2. Solvent-Extractable Compounds under Four Treatments
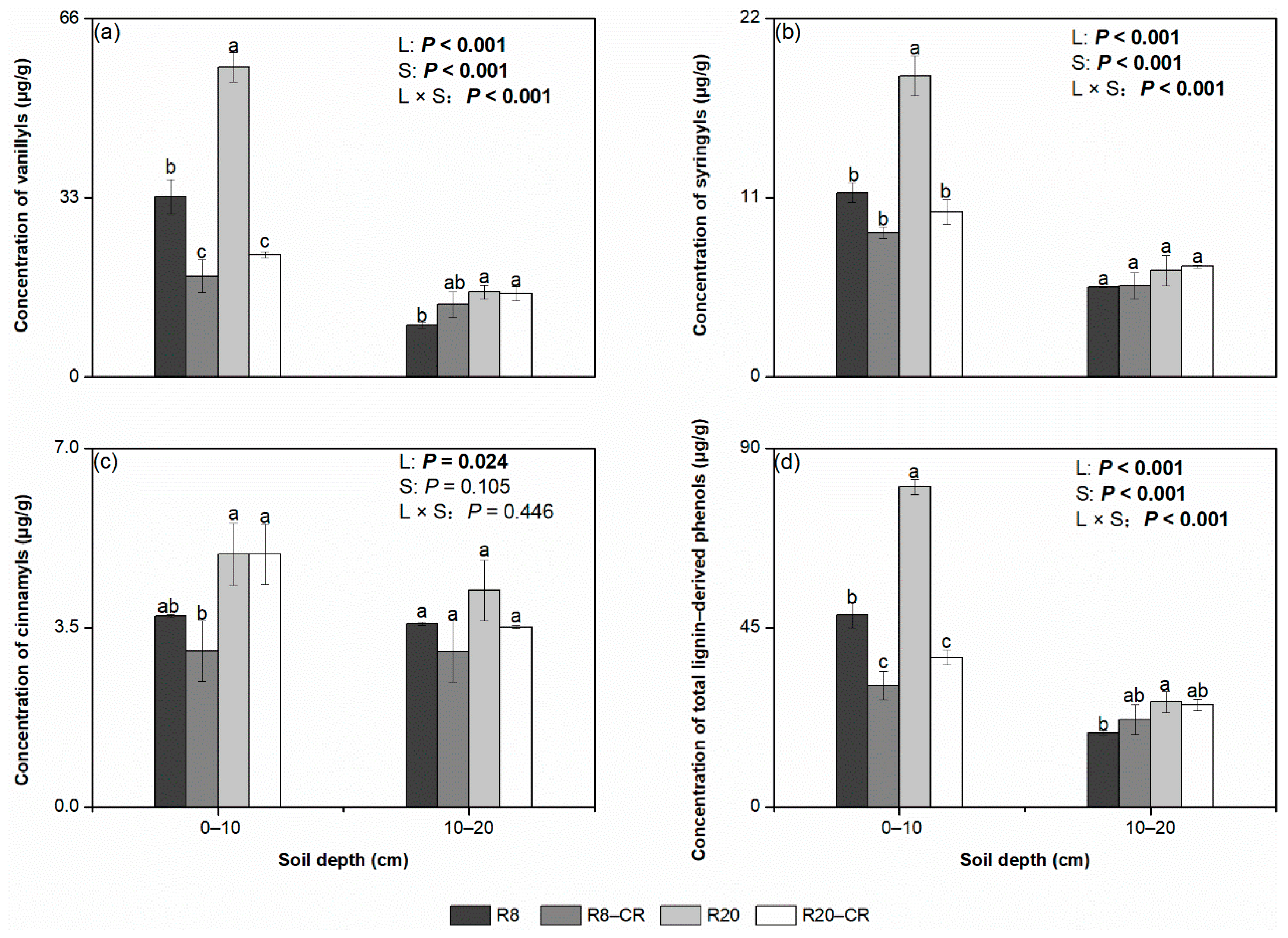
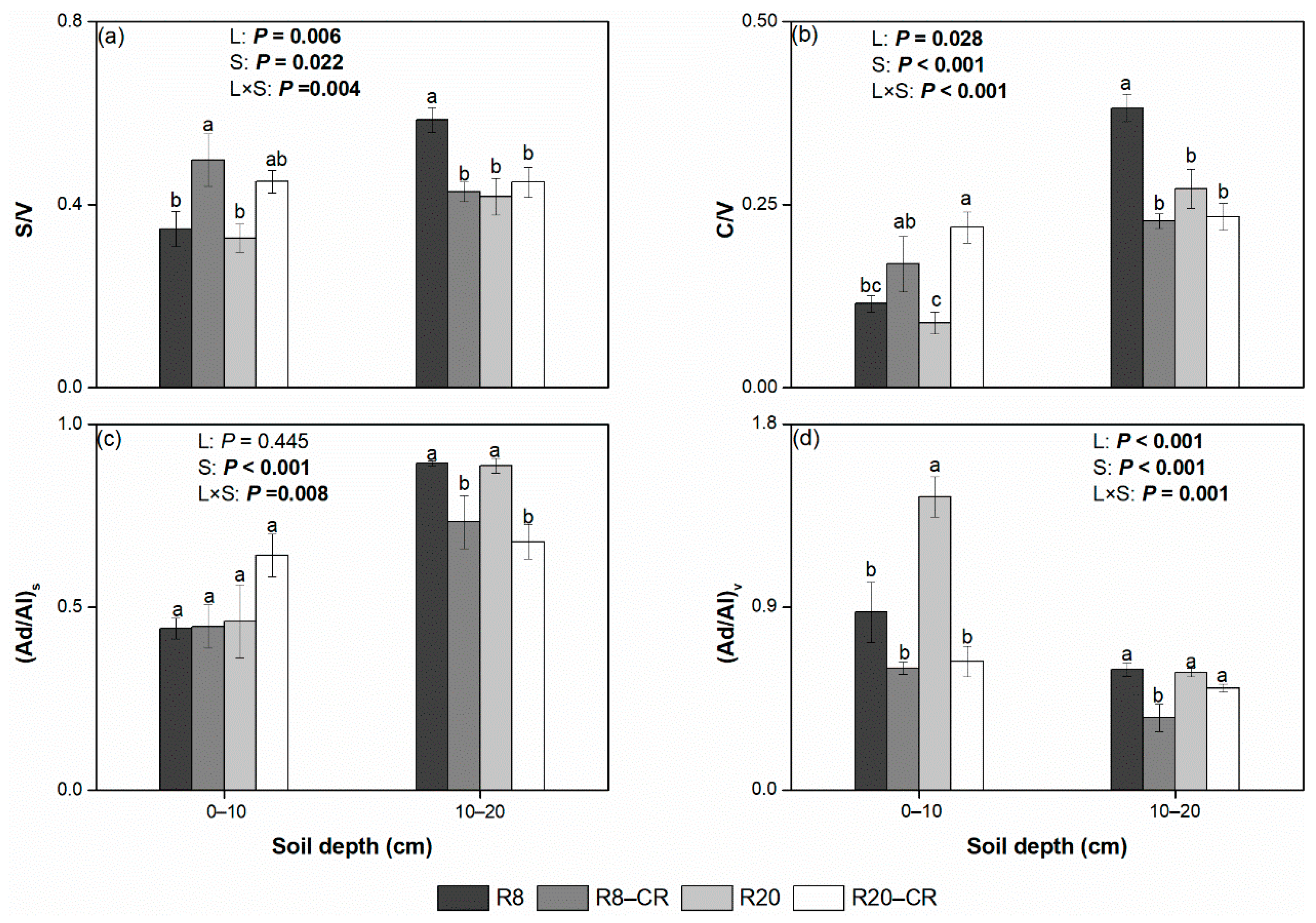
3.3. Lignin-Derived Phenols under Four Treatments
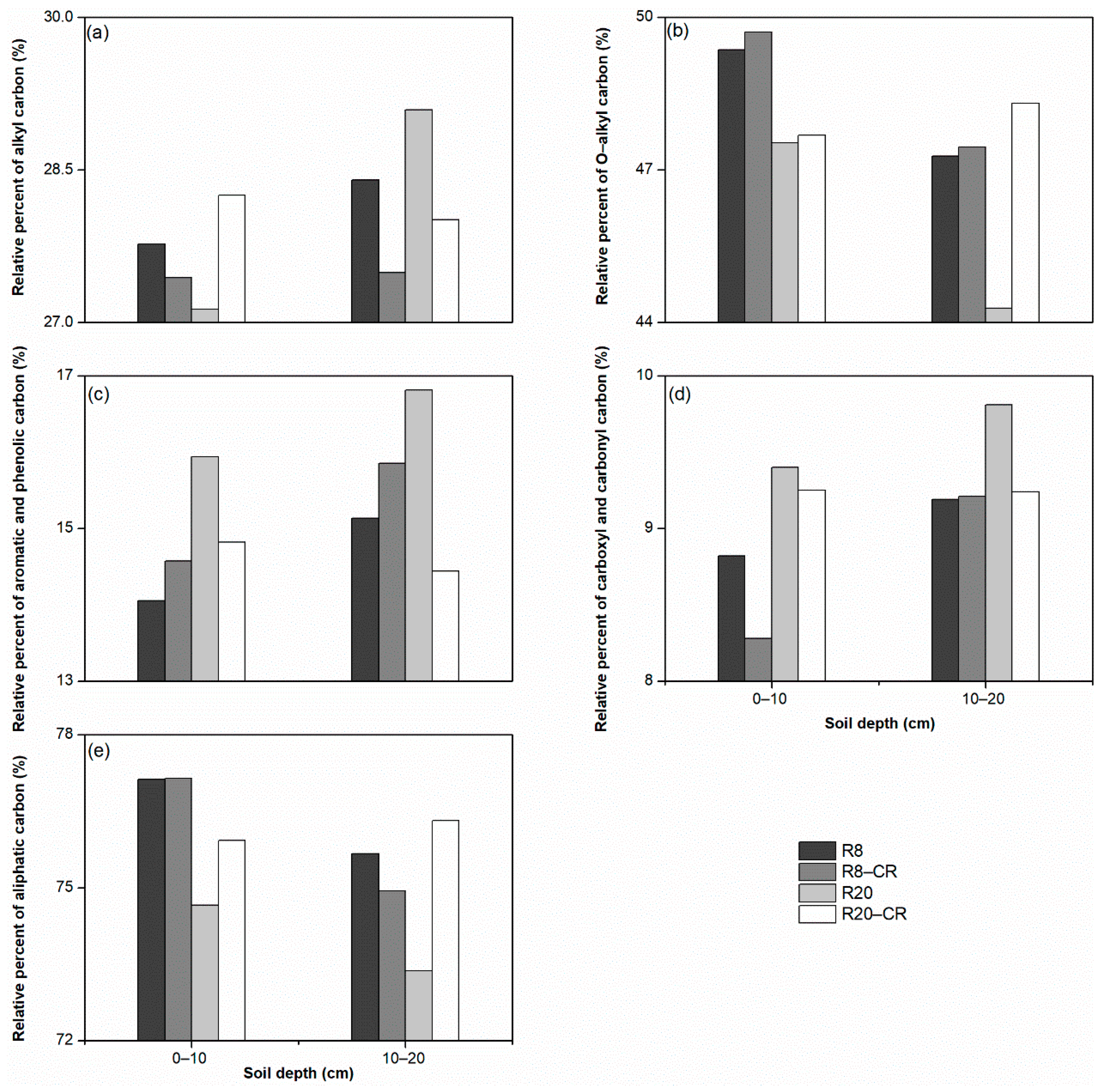
3.4. Chemical Composition of SOM Measured by NMR
4. Discussion
4.1. Effect of Land Use Types on SOC and MBC Concentrations and SOC Stocks
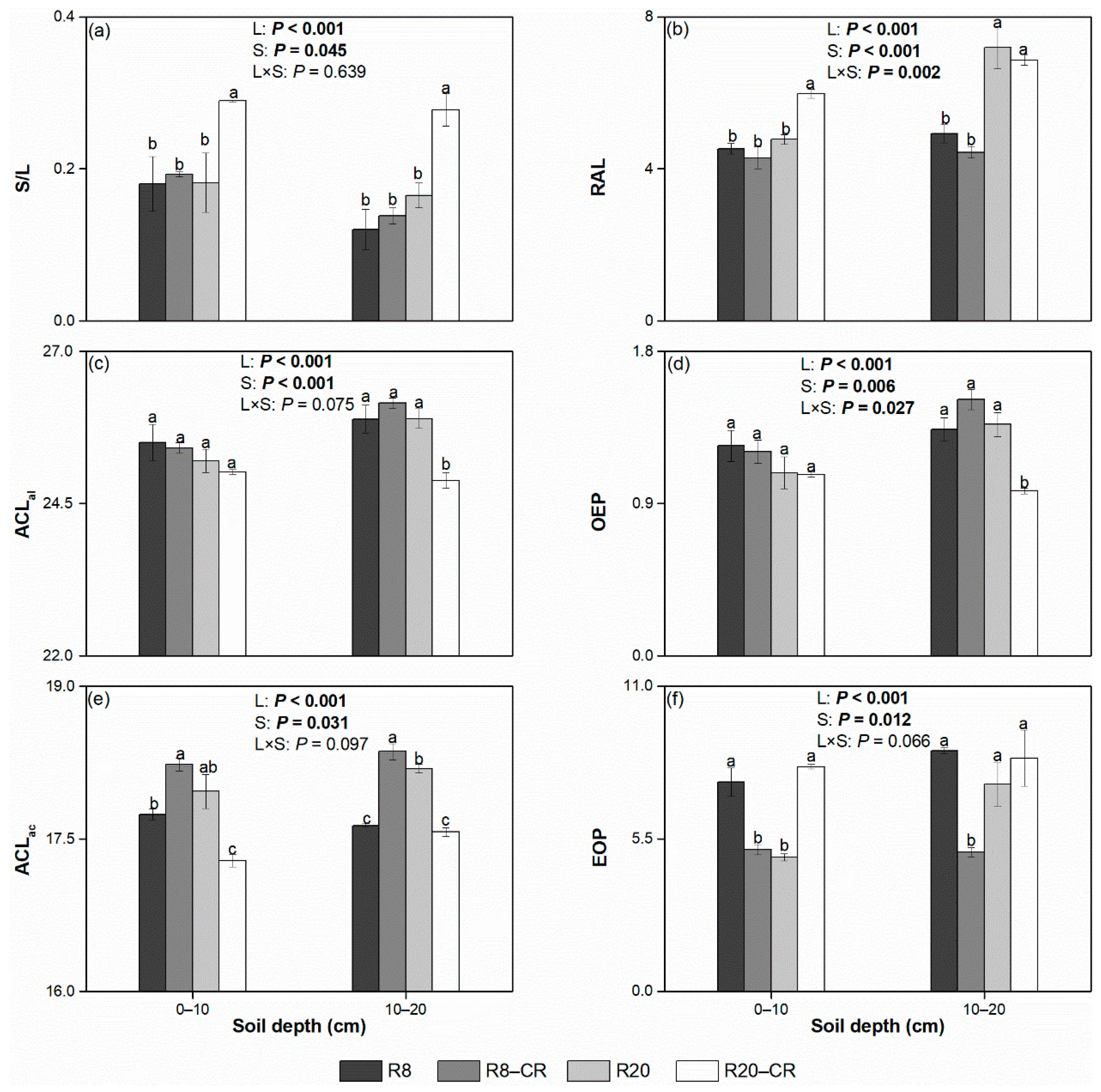
4.2. Effect of Land Use Types on the Solvent-Extractable Compounds
4.3. Effect of Land Use Types on Lignin-Derived Phenols
4.4. Effect of Land Use Types on the NMR Chemical Composition of SOM
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Spaccini, R.; Piccolo, A.; Conte, P.; Haberhauer, G.; Gerzabek, M.H. Increased soil organic carbon sequestration through hydrophobic protection by humic substances. Soil Biol. Biochem. 2002, 34, 1839–1851. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Gardi, C.; Sconosciuto, F. Evaluation of carbon stock variation in Northern Italian soils over the last 70 years. Sustain. Sci. 2007, 2, 237–243. [Google Scholar] [CrossRef]
- Minasny, B.; Sulaeman, Y.; Mcbratney, A.B. Is soil carbon disappearing? The dynamics of soil organic carbon in Java. Glob. Chang. Biol. 2011, 17, 1917–1924. [Google Scholar] [CrossRef]
- Savarese, C.; Drosos, M.; Spaccini, R.; Cozzolino, V.; Piccolo, A. Molecular characterization of soil organic matter and its extractable humic fraction from long-term field experiments under different cropping systems. Geoderma 2021, 383, 114700. [Google Scholar] [CrossRef]
- Baldock, J.A.; Skjemstad, J.O. Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org. Geochem. 2000, 31, 697–710. [Google Scholar] [CrossRef]
- Pisani, O.; Hills, K.M.; Courtier-Murias, D.; Simpson, A.J.; Mellor, N.J.; Paul, E.A.; Morris, S.J.; Simpson, M.J. Molecular level analysis of long term vegetative shifts and relationships to soil organic matter composition. Org. Geochem. 2013, 62, 7–16. [Google Scholar] [CrossRef]
- Eglinton, G.; Calvin, M. Chemical fossils. Sci. Am. 1967, 216, 32–43. [Google Scholar] [CrossRef]
- Otto, A.; Simpson, M.J. Evaluation of CuO oxidation parameters for determining the source and stage of lignin degradation in soil. Biogeochemistry 2006, 80, 121–142. [Google Scholar] [CrossRef]
- Feng, X.J.; Simpson, M.J. The distribution and degradation of biomarkers in Alberta grassland soil profiles. Org. Geochem. 2007, 38, 1558–1570. [Google Scholar] [CrossRef]
- Otto, A.; Shunthirasingham, C.; Simpson, M.J. A comparison of plant and microbial biomarkers in grassland soils from the Prairie Ecozone of Canada. Org. Geochem. 2005, 36, 425–448. [Google Scholar] [CrossRef]
- Schäfer, I.K.; Lanny, V.; Franke, J.; Eglinton, T.I.; Zech, M.; Vysloužilová, B.; Zech, R. Leaf waxes in litter and topsoils along a European transect. Soil 2016, 2, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Baldock, J.A.; Preston, C.M. Chemistry of Carbon Decomposition Processes in Forests as Revealed by Solid-State Carbon-13 Nuclear Magnetic Resonance. Carbon Forms Funct. For. Soils 1995, 89–117. [Google Scholar] [CrossRef]
- Pisani, O.; Haddix, M.L.; Conant, R.T.; Paul, E.A.; Simpson, M.J. Molecular composition of soil organic matter with land-use change along a bi-continental mean annual temperature gradient. Sci. Total Environ. 2016, 573, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Simpson, M.J.; Simpson, A.J.; Kingery, W.L. Solid-State 13C Nuclear Magnetic Resonance (NMR) analysis of soil organic matter. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Wei, X.R.; Qiu, L.P.; Shao, M.A.; Zhang, X.C.; Gale, W.J. The accumulation of organic carbon in mineral soils by afforestation of abandoned Farmland. PLoS ONE 2012, 7, e32054. [Google Scholar] [CrossRef]
- Song, X.S.; Shi, S.M.; Lu, S.; Ren, R.X.; He, C.X.; Meng, P.; Zhang, J.S.; Yin, C.J.; Zhang, X. Changes in soil chemical properties following afforestation of cropland with Robinia pseudoacacia in the southeastern Loess Plateau of China. For. Ecol. Manag. 2021, 487, 118993. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Powlson, D.S. The effects of biocidal treatments on metabolism in soil-V: A method for measuring soil biomass. Soil Biol. Biochem. 1976, 8, 209–213. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Wiley: Hoboken, NJ, USA, 1982. [Google Scholar]
- Schmidt, M.; Knicker, H.; Hatcher, P.G.; Kogel-Knabner, I. Improvement of 13C and 15N CPMAS NMR spectra of bulk soils, particle size fractions and organic material by treatment with 10% hydrofluoric acid. Eur. J. Soil Sci. 1997, 48, 319–328. [Google Scholar] [CrossRef]
- Ellert, B.H.; Bettany, J.R. Calculation of organic matter and nutrients stored in soils under contrasting management regimes. Can. J. Soil Sci. 1995, 75, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Hoefs, M.J.L.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S. The influence of oxic degradation on the sedimentary biomarker record I: Evidence from Madeira Abyssal plain turbidites. Geochim. Cosmochim. Acta 2002, 66, 2719–2735. [Google Scholar] [CrossRef]
- Bush, R.T.; McInerney, F.A. Leaf wax n-alkane distributions in and across modern plants: Implications for paleoecology and chemotaxonomy. Geochim. Cosmochim. Acta 2013, 117, 161–179. [Google Scholar] [CrossRef]
- Guo, Z.; Han, J.; Li, J.; Xu, Y.; Wang, X. Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 2019, 14, e0211163. [Google Scholar]
- Henrique, M.R.; Ribeiro, D.F.; Fátima, A.; Ernesto, V.; João, C.; Roland, B.; Fernanda, C. Carbon mineralization kinetics in an organically managed Cambic Arenosol amended with organic fertilizers. J. Plant Nutr. Soil Sci. 2010, 173, 39–45. [Google Scholar]
- Jackson, L.E.; Calderon, F.J.; Steenwerth, K.L.; Scow, K.M.; Rolston, D.E. Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma 2003, 114, 305–317. [Google Scholar] [CrossRef]
- Liu, Z.P.; Shao, M.A.; Wang, Y.Q. Effect of environmental factors on regional soil organic carbon stocks across the Loess Plateau region, China. Agric. Ecosyst. Environ. 2011, 142, 184–194. [Google Scholar] [CrossRef]
- Chang, R.Y.; Fu, B.J.; Liu, G.H.; Wang, S.; Yao, X.L. The effects of afforestation on soil organic and inorganic carbon: A case study of the Loess Plateau of China. Catena 2012, 95, 145–152. [Google Scholar] [CrossRef]
- Girona-García, A.; Badía-Villas, D.; Jiménez-Morillo, N.J.; González-Pérez, J.A. Changes in soil organic matter composition after Scots pine afforestation in a native European beech forest revealed by analytical pyrolysis (Py-GC/MS). Sci. Total Environ. 2019, 691, 1155–1161. [Google Scholar] [CrossRef]
- Ma, L.X.; Ju, Z.Q.; Fang, Y.Y.; Vancov, T.; Gao, Q.Q.; Wu, D.; Zhang, A.P.; Wang, Y.N.; Hu, C.S.; Wu, W.L.; et al. Soil warming and nitrogen addition facilitates lignin and microbial residues accrual in temperate agroecosystems. Soil Biol. Biochem. 2022, 170, 108693. [Google Scholar] [CrossRef]
- Naafs, D.F.W.; van Bergen, P.F.; Boogert, S.J.; de Leeuw, J.W. Solvent-extractable lipids in an acid andic forest soil; variations with depth and season. Soil Biol. Biochem. 2004, 36, 297–308. [Google Scholar] [CrossRef]
- Amblès, A.; Jacquesy, J.C.; Jambu, P.; Joffre, J.; Maggi-Churin, R. Polar lipid fraction in soil: A kerogen-like matter. Org. Geochem. 1991, 17, 341–349. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Jolivet, C.; Angers, D.A.; Chantigny, M.H.; Andreux, F.; Arrouays, D. Carbohydrate dynamics in particle—Size fractions of sandy spodosols following forest conversion to maize cropping. Soil Biol. Biochem. 2006, 38, 2834–2842. [Google Scholar] [CrossRef]
- Boeni, M.; Bayer, C.; Dieckow, J.; Conceição, P.C.; Dick, D.P.; Knicker, H.; Salton, J.C.; Macedo, M.C.M. Organic matter composition in density fractions of Cerrado Ferralsols as revealed by CPMAS 13C NMR: Influence of pastureland, cropland and integrated crop-livestock. Agric. Ecosyst. Environ. 2014, 190, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Nierop, K.G.J.; Verstraten, J.M. Organic matter formation in sandy subsurface horizons of Dutch coastal dunes in relation to soil acidification. Org. Geochem. 2003, 34, 499–513. [Google Scholar] [CrossRef]
- Bull, I.D.; van Bergen, P.F.; Nott, C.J.; Poulton, P.R.; Evershed, R.P. Organic geochemical studies of soils from the Rothamsted classical experiments—V. The fate of lipids in different long-term experiments. Org. Geochem. 2000, 31, 389–408. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, C.W.; Prater, I.; Angst, Š.; Peterse, F.; Nierop, K.G.J. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Commun. Biol. 2019, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Angst, G.; Mueller, K.E.; Nierop, K.G.; Simpson, M.J. Plant-or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Yang, Y.; Dou, Y.X.; Wang, B.R.; Wang, Y.Q.; Liang, C.; An, S.S.; Soromotin, A.; Kuzyakov, Y. Increasing contribution of microbial residues to soil organic carbon in grassland restoration chronosequence. Soil Biol. Biochem. 2022, 170, 108688. [Google Scholar] [CrossRef]
- Gao, W.T.; Wang, Q.T.; Zhu, X.M.; Liu, Z.F.; Li, N.; Xiao, J.; Sun, X.P.; Yin, H.J. The vertical distribution pattern of microbial- and plant-derived carbon in the rhizosphere in alpine coniferous forests. Rhizosphere 2021, 20, 100436. [Google Scholar] [CrossRef]
- Tückmantel, T.; Leuschner, C.; Preusser, S.; Kandeler, E.; Angst, G.; Mueller, C.W.; Meier, I.C. Root exudation patterns in a beech forest: Dependence on soil depth, root morphology, and environment. Soil Biol. Biochem. 2017, 107, 188–197. [Google Scholar] [CrossRef]
- Jansen, B.; Wiesenberg, G. Opportunities and limitations related to the application of plant-derived lipid molecular proxies in soil science. Soil 2017, 3, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Q.; Anderson, B.J.; Vogeler, I.; Schwendenmann, L. Long-chain n-alkane and n-fatty acid characteristics in plants and soil—Potential to separate plant growth forms, primary and secondary grasslands? Sci. Total Environ. 2018, 645, 1567–1578. [Google Scholar] [CrossRef]
- Schulten, H.R.; Schnitzer, M. Aliphatics in soil organic matter in fine-clay fractions. Soil Sci. Soc. Am. J. 1990, 54, 98–105. [Google Scholar] [CrossRef]
- Wiesenberg, G.L.; Dorodnikov, M.; Kuzyakov, Y. Source determination of lipids in bulk soil and soil density fractions after four years of wheat cropping. Geoderma 2010, 156, 267–277. [Google Scholar] [CrossRef]
- Zech, M.; Buggle, B.; Leiber, K.; Markovic, S.; Glaser, B.; Hambach, U.; Huwe, B.; Stevens, T.; Sümegi, P.; Wiesenberg, G.; et al. Reconstructing Quaternary vegetation history in the Carpathian Basin, SE Europe, using n-alkane biomarkers as molecular fossils—Problems and possible solutions, potential and limitations. J. Quat. Sci. 2009, 58, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Thevenot, M.; Dignac, M.; Rumpel, C. Fate of lignins in soils: A review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar] [CrossRef]
- Tamilarasan, K.; Sellamuthu, P.; Gurunathan, D.B. Integration of Lignin Removal from Black Liquor and Biotransformation Process. In Bioremediation: Applications for Environmental Protection and Management. Energy, Environment, and Sustainability; Varjani, S., Agarwal, A., Gnansounou, E., Gurunathan, B., Eds.; Springer: Singapore, 2018; pp. 77–97. [Google Scholar]
- Abiven, S.; Heim, A.; Schmidt, M.W.I. Lignin content and chemical characteristics in maize and wheat vary between plant organs and growth stages: Consequences for assessing lignin dynamic in soils. Plant Soil 2011, 343, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Ruckamp, D.; Martius, C.; Braganca, M.A.L.; Amelung, W. Lignin patterns in soil and termite nests of the Brazilian Cerrado. Appl. Soil Ecol. 2011, 48, 45–52. [Google Scholar] [CrossRef]
- Guggenberger, G.; Christensen, B.T.; Zech, W. Land-use effects on the composition of organic matter in particle-size separates of soil: I. Lignin and carbohydrate signature. Eur. J. Soil Sci. 1994, 45, 449–458. [Google Scholar] [CrossRef]
- Hedges, J.I.; Mann, D.C. The characterization of plant tissues by their lignin in oxidation products. Geochim. Cosmochim. Acta 1979, 43, 1803–1807. [Google Scholar] [CrossRef]
- Hedges, J.I.; Ertel, J.R. Characterization of lignin by gas capillary chromatography of cupric oxidation products. Anal. Chem. 1982, 54, 174–178. [Google Scholar] [CrossRef]
- Bahri, H.; Dignac, M.F.; Rumpel, C.; Rasse, D.P.; Chenu, C.; Mariotti, A. Lignin turnover kinetics in an agricultural soil in monomer specific. Soil Biol. Biochem. 2006, 38, 1977–1988. [Google Scholar] [CrossRef]
- Rumpel, C.; Eusterhues, K.; Kögel-Knabner, I. Location and chemical composition of stabilized organic carbon in topsoil and subsoil horizons of two acid forest soils. Soil Biol. Biochem. 2004, 36, 177–190. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.R.; Song, Z.C.; Yang, Y.J.; Wang, J.X.; You, Y.M.; Zhang, X.; Shi, Z.M.; Nong, Y.; Ming, A.G.; et al. Introducing nitrogen-fixing tree species and mixing with Pinus massoniana alters and evenly distributes various chemical compositions of soil organic carbon in a planted forest in southern China. For. Ecol. Manag. 2019, 449, 117477. [Google Scholar] [CrossRef]
- Zech, W.; Ziegler, F.; Kögel-Knabner, I.; Haumaier, L. Humic substances distribution and transformation in forest soil. Sci. Total Environ. 1992, 117/118, 155–174. [Google Scholar] [CrossRef]
- Dieckow, J.; Bayer, C.; Conceição, C.; Zanatta, J.A.; Martin-Neto, L.; Milorid, D.B.M.; Salton, J.C.; Macedo, M.M.; Mielniczuk, J.; Hernani, L.C. Land use, tillage, texture and organic matter stock and composition in tropical and subtropical Brazilian soils. Eur. J. Soil Sci. 2009, 60, 240–249. [Google Scholar] [CrossRef]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.G.; Khanna, P.K. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Tree Height (m) | Diameter at Breast Height (cm) | Understory Vegetation/Crop Type | Soil Bulk Density (g·cm−3) |
|---|---|---|---|---|
| R8 | 9.52 | 10.31 | Potentilla reptans L. var. sericophylla Franch., Carex breviculmis R. Br., and Setaria viridis (L.) Beauv. | 1.35 (0.16) |
| R8-CR | – | – | Wheat and oilseed rape rotation | 1.58 (0.13) |
| R20 | 9.71 m | 16.3 | Potentilla reptans L. var. sericophylla Franch., Carex breviculmis R. Br., and Setaria viridis (L.) Beauv. | 1.35 (0.18) |
| R20-CR | – | – | Wheat and oilseed rape rotation | 1.43 (0.23) |
| Soil Depth (cm) | Treatment | MBC (g·kg−1) | SOC (g·kg−1) |
|---|---|---|---|
| 0–10 | R8 | 0.30 (0.05) b | 9.98 (0.84) ab |
| R8-CR | 0.62 (0.11) a | 7.12 (0.46) bc | |
| R20 | 0.72 (0.06) a | 12.55 (1.04) a | |
| R20-CR | 0.78 (0.06) a | 5.25 (0.64) c | |
| 10–20 | R8 | 0.19 (0.02) b | 6.34 (0.13) a |
| R8-CR | 0.74 (0.08) a | 8.02 (0.68) a | |
| R20 | 0.78 (0.13) a | 8.36 (0.94) a | |
| R20-CR | 0.91 (0.06) a | 7.02 (0.12) a |
| Treatment | SOC Stock (Mg·ha−1) |
|---|---|
| R8 | 21.11 (1.24) ab |
| R8-CR | 20.53 (1.81) b |
| R20 | 27.24 (2.56) a |
| R20-CR | 15.96 (0.98) b |
| Soil Depth (cm) | Treatment | n-Alkanes | n-Alkanols | n-Alkanoic Acids | Carbohydrates | Steroids | Short-Chain Aliphatic Lipids | Long-Chain Aliphatic Lipids |
|---|---|---|---|---|---|---|---|---|
| μg·g−1 | ||||||||
| 0–10 | R8 | 9.35 (0.48) a | 10.47 (1.20) c | 35.78 (3.83) c | 19.73 (1.21) a | 12.33 (1.34) b | 32.17 (3.92) c | 23.44 (1.44) b |
| R8-CR | 8.68 (0.93) a | 9.57 (0.88) c | 37.84 (4.51) bc | 16.16 (1.13) b | 13.01 (0.67) ab | 32.34 (4.00) bc | 23.76 (2.27) b | |
| R20 | 8.19 (0.28) a | 14.58 (0.97) b | 49.24 (3.16) b | 14.72 (1.38) b | 15.11 (0.86) ab | 42.90 (3.34) b | 29.10 (1.00) a | |
| R20-CR | 9.77 (0.14) a | 18.60 (0.74) a | 63.87 (2.48) a | 18.24 (0.34) ab | 15.47 (0.80) a | 60.39 (2.74) a | 31.86 (0.57) a | |
| 10–20 | R8 | 5.65 (0.33) b | 3.92 (0.06) b | 17.83 (0.23) b | 9.70 (0.34) ab | 5.58 (0.17) c | 16.18 (0.26) c | 11.22 (0.46) b |
| R8-CR | 5.66 (0.33) b | 4.74 (0.18) b | 20.46 (0.38) b | 8.92 (0.28) b | 6.98 (0.24) b | 17.00 (0.37) c | 13.86 (0.36) b | |
| R20 | 6.39 (0.38) b | 4.82 (0.12) b | 30.67 (3.68) a | 10.96 (0.47) a | 5.82 (0.30) bc | 26.85 (3.54) b | 15.03 (0.49) a | |
| R20-CR | 10.20 (1.16) a | 9.22 (0.62) a | 37.16 (3.07) a | 10.89 (0.60) a | 8.23 (0.60) a | 35.20 (2.88) a | 21.38 (1.96) a | |
| Soil Depth (cm) | Treatment | Alkyl C/O-Alkyl Ratio | Aromatic/O-Alkyl Ratio | Aliphatic/Aromatic Ratio |
|---|---|---|---|---|
| 0–10 | R8 | 0.56 | 0.28 | 5.49 |
| R8-CR | 0.55 | 0.29 | 5.30 | |
| R20 | 0.57 | 0.34 | 4.68 | |
| R20-CR | 0.59 | 0.31 | 5.12 | |
| 10–20 | R8 | 0.60 | 0.32 | 5.00 |
| R8-CR | 0.58 | 0.33 | 4.73 | |
| R20 | 0.66 | 0.38 | 4.36 | |
| R20-CR | 0.58 | 0.30 | 5.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Guo, J.; Wang, X.; Du, Z.; Ren, R.; Lu, S.; He, C. Afforestation Alters the Molecular Composition of Soil Organic Matter in the Central Loess Plateau of China. Forests 2023, 14, 1502. https://doi.org/10.3390/f14071502
Song X, Guo J, Wang X, Du Z, Ren R, Lu S, He C. Afforestation Alters the Molecular Composition of Soil Organic Matter in the Central Loess Plateau of China. Forests. 2023; 14(7):1502. https://doi.org/10.3390/f14071502
Chicago/Turabian StyleSong, Xueshu, Jingwen Guo, Xiao Wang, Zhangliu Du, Rongxiu Ren, Sen Lu, and Chunxia He. 2023. "Afforestation Alters the Molecular Composition of Soil Organic Matter in the Central Loess Plateau of China" Forests 14, no. 7: 1502. https://doi.org/10.3390/f14071502