
Effects of Changing Restoration Years on Soil Nutrient Traits and Plant Community Diversity in a Phosphate Mining Area
Abstract
:1. Introduction
2. Materials and Methods
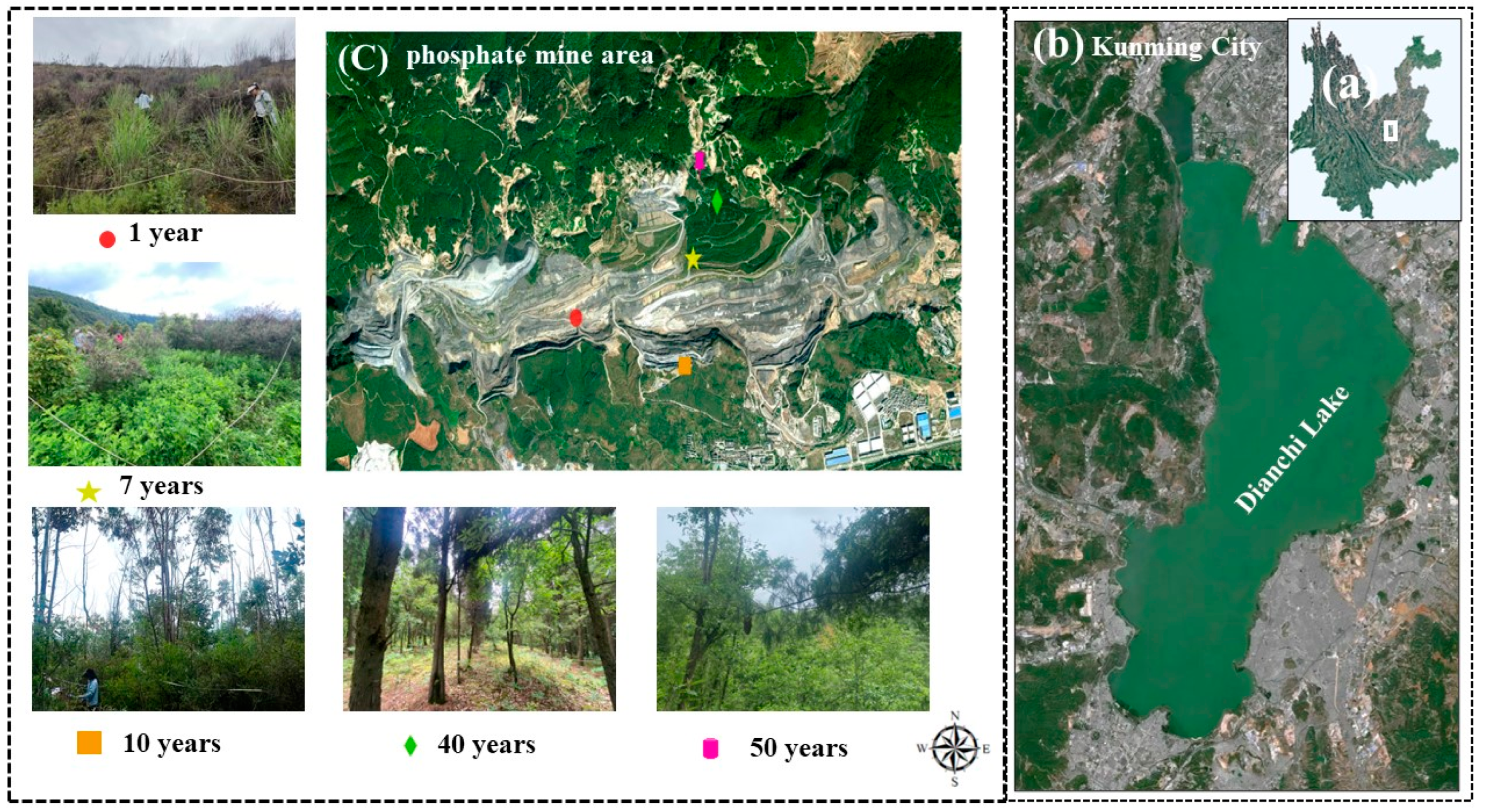
2.1. Study Site Description
2.2. Sample Site Survey
2.3. Soil Collection and Determination Indicators
2.4. Soil determination Methods
2.5. Statistical Analysis
3. Results
3.1. Plant Community Composition
3.2. Species Diversity in Communities
3.3. Nutrient Characteristics of Soils
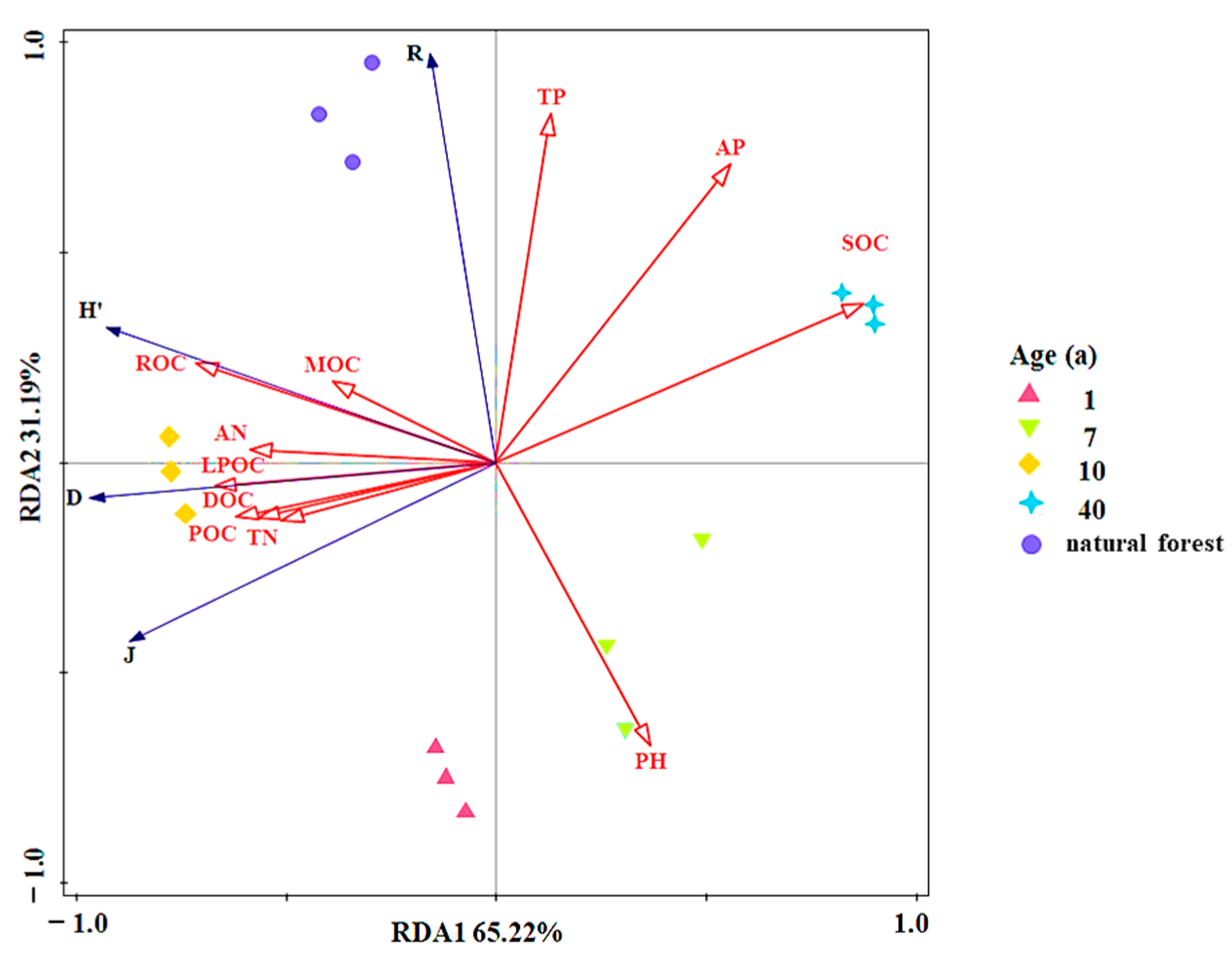
3.4. Relation between Plant Diversity to Soil Nutrients and Restoration Years
4. Discussion
4.1. Species Composition and Diversity in Phosphate Mining Areas at Different Restoration Years
4.2. Soil Nitrogen and Phosphorus Nutrient Characteristics in Different Restoration Years
4.3. Effects of Different Restoration Years on Plant Community Diversity and Soil Organic Carbon
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malaviya, S.; Munsi, M.; Oinam, G.; Joshi, P. Landscape approach for quantifying land use land cover change (1972–2006) and habitat diversity in a mining area in Central India (Bokaro, Jharkhand). Environ. Monit. Assess. 2010, 170, 215–229. [Google Scholar] [CrossRef]
- Adlassnig, W.; Weiss, Y.S.; Sassmann, S.; Steinhauser, G.; Hofhansl, F.; Baumann, N.; Lichtscheidl, I.K.; Lang, L. The copper spoil heap Knappenberg, Austria, as a model for metal habitats—Vegetation, substrate and contamination. Sci Total Environ. 2016, 563–564, 1037–1049. [Google Scholar] [CrossRef]
- Gastauer, M.; Souza, F.P.; Ramos, S.J.; Caldeira, C.F.; Silva, J.R.; Siqueir, J.O.; Furtini Neto, A.E. Mine land rehabilitation in Brazil: Goals and techniques in the context of legal requirements. Ambio 2019, 48, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M.; Bhatia, S.; Shenoy, K. Vegetation-environment relationships in a South Asian tropical montane grassland ecosystem: Restoration implications. Trop. Ecol. 2015, 56, 201. [Google Scholar] [CrossRef]
- Daily, G.C. Restoring Value to the World’s degraded lands. Science 1995, 269, 350–354. [Google Scholar] [CrossRef]
- Singh, R.S.; Tripathi, N.; Chaulya, S.K. Ecological study of revegetated coal mine spoil of an Indian dry tropical ecosystem along an age gradient. Biodegradation 2012, 23, 837–849. [Google Scholar] [CrossRef]
- Martínez-Ruiz, C.; Fernández-Santos, B.; Putwain, P.D.; Fernández-Gómez, M.J. Natural and man-induced revegetation on mining wastes: Changes in the floristic composition during early succession. Ecol. Eng. 2007, 30, 286–294. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Bai, X.; Zhou, Y.B.; Zhu, W.X.; Yin, Y. Variations of soil microbial communities accompanied by different vegetation restoration in an open-cut iron mining area. Sci. Total Environ. 2020, 704, 135243. [Google Scholar] [CrossRef]
- Li, S.; Liber, K. Influence of different revegetation choices on plant community and soil development nine years after initial planting on a reclaimed coal gob pile in the Shanxi mining area, China. Sci. Total Environ. 2018, 618, 1314–1323. [Google Scholar] [CrossRef]
- Putten, W.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedback: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Thakur, M.P.; Putten, W.H.; Wilschut, R.A.; Veen, G.F.; Kardol, P.; van Ruijven, J.; Allan, E.; Roscher, C.; van Kleunen, M.; Bezemer, T.M. Plant-soil feedbacks and temporal dynamics of plant diversity-productivity relationships. Trends Ecol. Evol. 2021, 36, 651–661. [Google Scholar] [CrossRef]
- Kulmatiski, A.; Beard, K.H.; Stevens, J.R.; Cobbold, S.M. Plant-soil feedbacks: A meta-analytical review. Ecol. Lett. 2008, 11, 980–992. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Z.; Jia, J.Y.; Zhang, J.L. Plant soil feedback theory and its applications and agricultural ecosystems. Acta Ecologica Sinica. 2021, 41, 14. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Wang, S.L.; Yang, Y. Concentrations and resorption patterns of 13 nutrients in different plant functional types in the karst region of south-western China. Ann. Bot. 2014, 113, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, X.; Xu, C.Y.; Hao, M.H.; Choe, C.H.; He, H.J. Thinning can increase shrub diversity and decrease herb diversity by regulating light and soil environments. Front Plant Sci. 2022, 13, 948648. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhao, Z.Q.; Niu, S.Y.; Bai, Z.K. The reclaimed coal mine ecosystem diverges from the surrounding ecosystem and reaches a new self-sustaining state after 20–23 years of succession in the Loess Plateau area, China. Sci. Total Environ. 2020, 727, 138739. [Google Scholar] [CrossRef]
- Suding, K.N.; Gross, K.L.; Houseman, G.R. Alternative states and positive feedbacks in restoration ecology. Trends Ecol. Evol. 2004, 19, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Juwarkar, A.A.; Singh, L.; Singh, S.K.; Jambhulkar, H.P.; Thawale, P.R.; Kanfade, H. Natural vs. reclaimed forests—A case study of successional change, reclamation technique and phytodiversity. Int. J. Min. Reclam. Environ. 2014, 29, 1–23. [Google Scholar] [CrossRef]
- Chiquoine, L.P.; Abella, S.R.; Bowker, M.A. Rapidly restoring biological soil crusts and ecosystem functions in a severely disturbed desert ecosystem. Ecol. Appl. 2016, 26, 1260–1272. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Li, X.Y.; Liu, X.M. Restoration of nutrient in phosphate mined-out areas: A case study of the phosphate mine in kunyang, Yunnan. Genom. Appl. Biol. 2017, 36, 8. [Google Scholar]
- Zhou, J.H.; Yu, L.; Wang, Y.S.; Xiao, Y.N.; Zhu, Y.L. Effects of Magnolia multiflora. with different growth years on soil nutrient changes in Phosphorus mine slope. Soil Water Conserv. China 2022, 4, 38–41. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; Zhong, M.C. Ecosystem carbon storage and soil organic carbon stability in pure and mixed stands of Cunninghamia lanceolata and Michelia macclurei. Plant Soil. 2013, 370, 295–304. [Google Scholar] [CrossRef]
- Fang, J.Y.; Wang, X.P.; Shen, Z.H.; Tang, Z.Y.; He, J.S.; Yu, D.; Jiang, Y.; Wang, Z.H.; Zheng, C.Y.; Zhu, J.L.; et al. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533–548. [Google Scholar] [CrossRef]
- Bao, S.D. Agricultural Soil Analysis, 3rd ed.; China Agricuture Press: Beijing, China, 2000. [Google Scholar]
- Marriott, E.E.; Wander, M.M. Total and labile soil organic matter in organic and conve entional farming systems. Soil Sci. Soc. Am. J. 2006, 70, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.; He, B.Y.; Li, C.; Hui, H.Y.; Liu, B.; Zhang, X.X. Analysis of grassland community-level plant functional traits and functional diversity at different times during restoration. Acta Prataculturae Sin. 2023, 32, 16–25. [Google Scholar] [CrossRef]
- Auclair, A.N.; Goff, F.G. Diversity relations of upland forests in the western great lakes area. Univ. Chic. Press J. 1971, 105, 946. [Google Scholar] [CrossRef]
- Li, T.; Wu, M.H.; Duan, C.Q.; Li, S.Y.; Liu, C.E. The effect of different restoration approaches on vegetation development in metal mines. Sci. Total Environ. 2022, 806, 150626. [Google Scholar] [CrossRef]
- Walker, L.R.; Walker, J.; Hobbs, R.J. Linking Restoration and Ecological Succession; Springer series on environmental management; Springer: New York, NY, USA, 2007. [Google Scholar]
- Feng, H.B.; Zhou, J.W.; Zhou, A.G.; Bai, G.Y.; Li, Z.X.; Chen, H.N.; Su, D.H.; Han, X. Grassland ecological restoration based on the relationship between vegetation and its below-ground habitat analysis in steppe coal mine area. Sci. Total Environ. 2021, 778, 146221. [Google Scholar] [CrossRef]
- Reed, C.C.; Berhe, A.A.; Moreland, K.C.; Wilcox, J.W.; Sullivan, B. Restoring function: Positive responses of carbon and nitrogen to 20 years of hydrologic restoration in montane meadows. Ecol. Appl. 2022, 32, e2677. [Google Scholar] [CrossRef]
- Yun, J.Y.; Guo, C.Y.; Li, J.C.; Wang, X.; Li, Q.R. Colonized wild plants in the reclamation area of the antaibao opencast coal mine. Arid. Zone Res. 2016, 33, 399–409. [Google Scholar] [CrossRef]
- Sun, C.; Chai, Z.; Liu, G.; Sha, X. Changes in species diversity patterns and spatial heterogeneity during the secondary succession of grassland vegetation on the loess plateau, China. Front Plant Sci. 2017, 8, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Nobile, C.M.; Bravin, M.N.; Becquer, T.; Paillat, J.M. Phosphorus sorption and availability in an andosol after a decade of organic or mineral fertilizer applications: Importance of pH and organic carbon modifications in soil as compared to phosphorus accumulation. Chemosphere 2020, 239, 124709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, X.; Li, Z.; Xu, C.H.; Luo, W. Improvements in soil quality with vegetation succession in subtropical China karst. Sci. Total Environ. 2021, 775, 145876. [Google Scholar] [CrossRef]
- Smethurst, P.; Holz, G.; Moroni, M.; Baillie, C. Nitrogen management in Eucalyptus nitens plantations. For. Ecol. Manag. 2004, 193, 63–80. [Google Scholar] [CrossRef]
- Li, H.; Wu, Y.; Liu, S.; Zhao, W.Z.; Xiao, J.F.; Winosiecki, L.A.; Vågen, T.G.; Xu, J.C.; Yin, X.Y.; Wang, F.; et al. The Grain-for-Green project offsets warming-induced soil organic carbon loss and increases soil carbon stock in Chinese Loess Plateau. Sci. Total Environ. 2022, 837, 155469. [Google Scholar] [CrossRef]
- Gu, L.P.; Kong, J.J.; Chen, K.; Guo, Y.Q. Monitoring soil biological properties during the restoration of a phosphate mine under different tree species and plantation types. Ecotoxicol. Environ. Saf. 2019, 180, 130–138. [Google Scholar] [CrossRef]
- Yamileth, D.H.; Elena, V.; Jesús, C.; Patrick, L.; Luis, F.C.; Juan, J.J. Evaluation of reclamation success in an open-pit coal mine using integrated soil physical, chemical and biological quality indicators. Ecol. Indic. 2019, 103, 182–193. [Google Scholar] [CrossRef]
- Wang, S.; Cao, Y.; Geng, B.; Yang, K.; Zhou, Z.K. Succession law and model of reconstructed soil quality in an open-pit coal mine dump of the loess area, China. J. Environ. Manag. 2022, 312, 114923. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.Y.; Liu, S.H.; Cheng, F.Y.; Su, X.K.; Dong, S.K.; Zhao, S.; Liu, G.H. Variability of environmental factors and the effects on vegetation diversity with different restoration years in a large open-pit phosphorite mine. Ecol. Eng. 2019, 127, 245–253. [Google Scholar] [CrossRef]
- Rüegg, J.; Quezada, J.C.; Santonja, M.; Ghazoul, J.; Kuzyakov, Y.; Buttler, A.; Guillaume, T. Drivers of soil carbon stabilization in oil palm plantations. Land Degrad. Dev. 2019, 30, 1904–1915. [Google Scholar] [CrossRef]
- Xu, H.W.; Qu, Q.; Wang, M.G.; Li, P.; Li, Y.Z.; Xue, S.; Liu, G.B. Soil organic carbon sequestration and its stability after vegetation restoration in the Loess Hilly Region, China. Land Degrad. Dev. 2020, 5, 568–580. [Google Scholar] [CrossRef]
- Brown, K.H.; Bach, E.M.; Drijber, R.A.; Hofmockel, K.S.; Jeske, E.S.; Sawyer, J.E.; Castellano, M.C. A long-term nitrogen fertilizer gradient has little effect on soil organic matter in a high-intensity maize production system. Glob. Chang. Biol. 2014, 20, 1339–1350. [Google Scholar] [CrossRef]
- Lajtha, K.; Bowden, R.D.; Crow, S.; Fekete, I.; Kotroczó, Z.; Plante, A.; Simpson, M.J.; Nadelhoffer, K.J. The detrital input and removal treatment (DIRT) network: Insights into soil carbon stabilization. Sci. Total Environ. 2018, 640–641, 1112–1120. [Google Scholar] [CrossRef]
- Rasmussen, C.; Heckman, K.; Wieder, W.R.; Keiluweit, M.; Lawrence, C.R.; Berhe, A.A.; Blankinship, J.C.; Crow, S.E.; Druhan, J.L.; Pries, C.E.H.; et al. Beyond clay: Towards an improved set of variables for predicting soil organic matter content. Biogeochemistry 2018, 137, 297–306. [Google Scholar] [CrossRef]
- Miltner, A.; Bombach, P.; Schmidt-Brücken, B.; Kästner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [Green Version]
- Panchal, P.; Preece, C.; Peñuelas, J.; Giri, J. Soil carbon sequestration by root exudate. Trends Plant Sci. 2022, 27, 749–757. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Restoration Year/a | Elevation/m | Slope Gradient/° | Slope Direction | Vegetation Coverage/% |
|---|---|---|---|---|
| 1 | 2113–2110 | 6 | southwest | 27% |
| 7 | 2057–2060 | 6 | west | 80% |
| 10 | 2038–2045 | 40 | southeast | 100% |
| 40 | 2218–2240 | 3 | southeast | 70% |
| natural forest | 2268–2273 | 40 | east | 100% |
| Number | Life Form | Species Name | Restoration Year/a | Natural Forest | |||
|---|---|---|---|---|---|---|---|
| 1 | 7 | 10 | 40 | ||||
| 1 | Tree | Pinus armandii Franch. | + | ||||
| 2 | Alnus cremastogyne Burk. | + | |||||
| 3 | Cupressus duclouxiana Hickel | + | |||||
| 4 | Quercus acutissima Carruth. | + | |||||
| 5 | Quercus aliena Bl. | + | |||||
| 6 | Eucalyptus pellita F.Muell. | + | |||||
| 7 | Shrub | Berberis silva-taroucana Schneid. | + | ||||
| 8 | Zanthoxylum simulans Hance. | + | |||||
| 9 | Buddleja officinalis Maxim. | + | |||||
| 10 | Cotoneaster pannosus Franch. | + | |||||
| 11 | Osyris wightiana Wall. ex Wight var. stipitata | + | |||||
| 12 | Lespedeza bicolor Turcz. | + | |||||
| 13 | Myrsine africana L. | + | |||||
| 14 | Indigofera tinctoria L. | + | |||||
| 15 | Cinnamomum camphora L. | + | + | ||||
| 16 | Buddleja asiatica Lour. | + | |||||
| 17 | Herb | Eupatorium coelestinum L. | + | + | + | ||
| 18 | Phytolacca americana L. | + | |||||
| 19 | Rubus ellipticus var. obcordatus (Franch.) Focke | + | |||||
| 20 | Pteris ensiformis Burm. | + | |||||
| 21 | Pteris aspericaulis var. cuspigera | + | |||||
| 22 | Rheum officinale Baill. | + | |||||
| 23 | Lophatherum gracile Brongn. | + | + | ||||
| 24 | Paeonia suffruticosa Andr. | + | |||||
| 25 | Erigeron canadensis L. | + | |||||
| 26 | Geranium wilfordii Maxim. | + | |||||
| 27 | Microbiota decussata Kom. | + | |||||
| 28 | Pinellia ternata (Thunb.) Makino | + | |||||
| 29 | Lonicera japonica Thunb. | + | |||||
| 30 | Medicago sativa L. | + | |||||
| 31 | Artemisia argyi Lévl. et Van. | + | |||||
| 32 | Imperata cylindrica (L.) Raeusch. | + | |||||
| 33 | Miscanthus sinensis Anderss. | + | |||||
| 34 | Rumex hastatus D. Don. | + | |||||
| Restoration Year/a | S | J | D |
|---|---|---|---|
| 1 | 4 | 1.31 | 0.68 |
| 7 | 5 | 0.98 | 0.67 |
| 10 | 8 | 1.34 | 0.83 |
| 40 | 9 | 0.66 | 0.44 |
| natural forest | 12 | 1.02 | 0.76 |
| pH | TN | AN | TP | AP | SOC | DOC | ROC | LPOC | POC | MOC | H′ | R | J | D | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H’ | −0.396 | 0.717 ** | 0.772 ** | −0.359 | −0.536 * | −0.913 ** | 0.052 | 0.236 | 0.217 | 0.572 * | 0.211 | 1 | −0.162 | 0.646 ** | 0.860 ** |
| R | −0.754 ** | −0.066 | 0.109 | 0.827 ** | 0.651 ** | 0.253 | −0.088 | 0.308 | −0.026 | −0.093 | 0.212 | −0.162 | 1 | 0.640 * | 0.202 |
| J | −0.851 ** | 0.426 | 0.569 * | 0.410 | 0.112 | −0.524 * | −0.005 | 0.438 | 0.126 | 0.089 | 0.006 | 0.646 ** | 0.640 * | 1 | 0.830 ** |
| D | −0.691 ** | 0.560 * | 0.625 | −0.031 | −0.031 | −0.654 ** | −0.307 | 0.088 | −0.169 | −0.127 | −0.527 * | 0.860 ** | 0.202 | 0.830 ** | 1 |
| Variable | Estimate (Standard Error) | ANOVA | Relative Importance | |
|---|---|---|---|---|
| F-Value | Pr (>F) | |||
| pH | −0.91 | 13.22 | *** | 26.5% |
| SOC | −0.20 | 14.58 | *** | 6.9% |
| TP | 0.96 | 25.96 | *** | 2.9% |
| AP | −0.60 | 24.65 | *** | 0.8% |
| age | −0.41 | 23.85 | *** | 7.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Kuang, S.; He, L.; Wang, C.; Zu, Y. Effects of Changing Restoration Years on Soil Nutrient Traits and Plant Community Diversity in a Phosphate Mining Area. Forests 2023, 14, 1468. https://doi.org/10.3390/f14071468
Xu C, Kuang S, He L, Wang C, Zu Y. Effects of Changing Restoration Years on Soil Nutrient Traits and Plant Community Diversity in a Phosphate Mining Area. Forests. 2023; 14(7):1468. https://doi.org/10.3390/f14071468
Chicago/Turabian StyleXu, Cui, Siping Kuang, Lu He, Chunxue Wang, and Yanqun Zu. 2023. "Effects of Changing Restoration Years on Soil Nutrient Traits and Plant Community Diversity in a Phosphate Mining Area" Forests 14, no. 7: 1468. https://doi.org/10.3390/f14071468