
The New Report of Root Rot on Fatsia japonica Caused by Phytophthora nicotianae in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Disease Investigation and Isolation
2.2. Morphological Identification
2.3. DNA Extraction and PCR Amplifcation
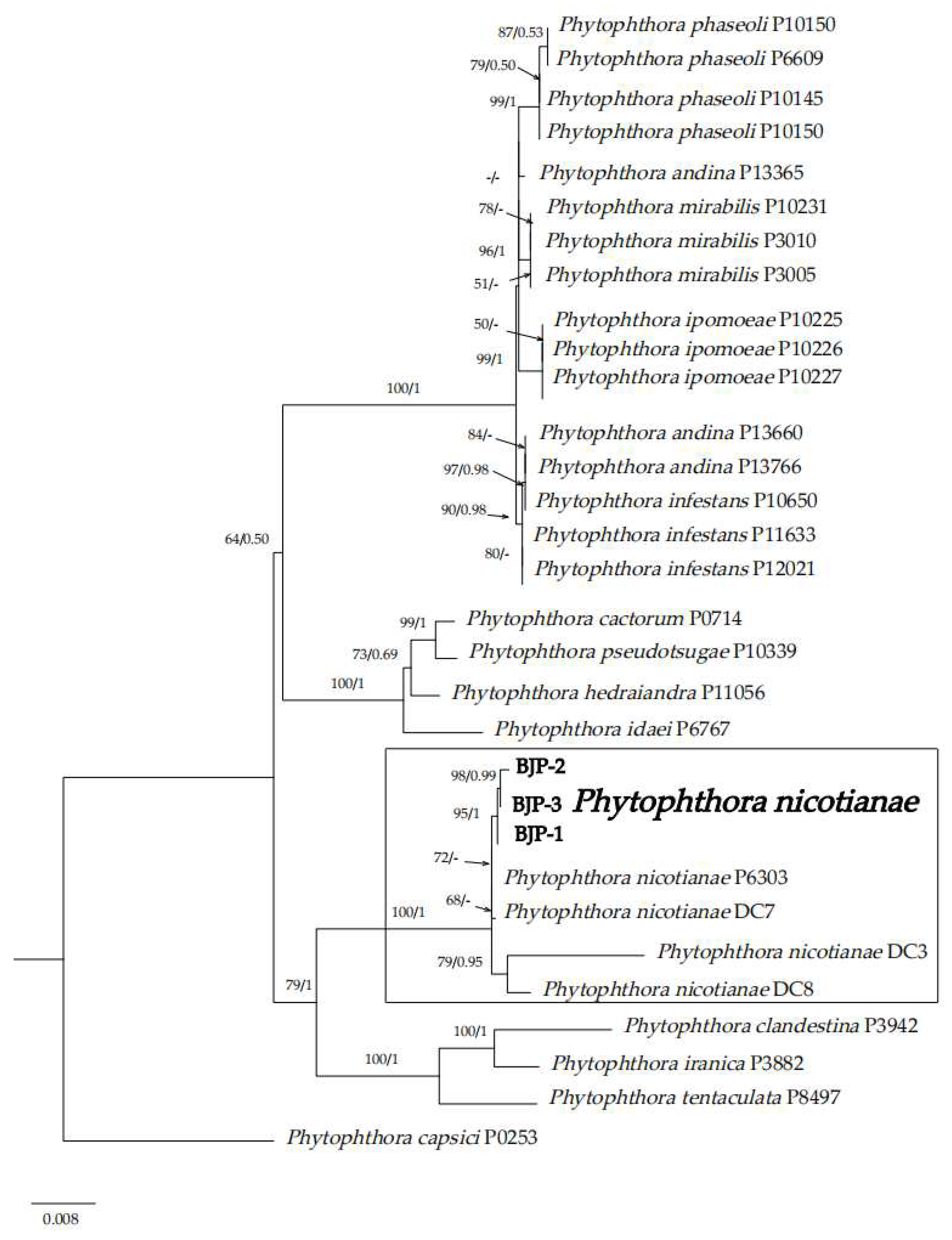
2.4. Phylogenetic Analyses
2.5. Pathogenicity Assays
3. Results
3.1. Natural Symptoms
3.2. Morphological Characteristics
3.3. Molecular Biology Identification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | DNA Target | GenBank Accession No. | Blast Match Sequence | |
|---|---|---|---|---|
| Reference Accession No. | Sequence Identity (%) | |||
| BJP-1 | ITS | OP735506 | P. nicotianae B2 (MT472132.1) | 99.88% (801/802) |
| LSU | OP738518 | P. nicotianae 22F9 (KX250514.1) | 100% (1246/1246) | |
| COXII | OP743911 | P. nicotianae P6303 (GU318304.1) | 99.73% (733/735) | |
| BJP-2 | ITS | OP735507 | P. nicotianae B2 (MT472132.1) | 99.63% (802/805) |
| LSU | OP738516 | P. nicotianae 22F9 (KX250514.1) | 100% (1248/1248) | |
| COXII | OP743912 | P. nicotianae P6303 (GU318304.1) | 99.66% (880/883) | |
| BJP-3 | ITS | OP735526 | P. nicotianae B2 (MT472132.1) | 100% (803/803) |
| LSU | OP738517 | P. nicotianae 22F9 (KX250514.1) | 100% (1245/1245) | |
| COXII | OP743913 | P. nicotianae P6303 (GU318304.1) | 99.61% (758/761) | |
3.4. Pathogenicity Tests of Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grishkovets, V.I.; Sobolev, E.A.; Shashkov, A.S.; Chirva, V.Y. Triterpene glycosides of Fatsia japonica. I. Isolation and structure of glycosides from Fatsia japonica seeds. Chem. Nat. Compd. 2000, 36, 166–169. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Feng, X.L.; Meng, Z.; Yang, B.X.; Gao, C.X.; Zhang, L.; Tian, J.K. The complete chloroplast genome sequence of Fatsia japonica (Apiales: Araliaceae) and the phylogenetic analysis. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 3050–3051. [Google Scholar] [CrossRef]
- Liang, Z.Y.; Gan, X.J.; Wang, D.P.; Liao, G.C.; Chen, H.P. Analysis and Comparison of Chemical Constituents of Essential oils from Stems, Leaves and Flowers (Fruits) of Fatsia japonica. J. Anhui Agric. Sci. 2012, 40, 8473–8475. [Google Scholar] [CrossRef]
- Xu, X.L.; Xiao, Q.G.; Yang, C.L.; Liu, Y.G. First Report of Anthracnose Caused by Colletotrichum karstii on Fatsia japonica in Sichuan, China. Plant Dis. 2020, 105, 216. [Google Scholar] [CrossRef] [PubMed]
- Hieno, A.; Li, M.Z.; Afandi, A.; Otsubo, K.; Suga, H.; Kageyama, K. Detection of the Genus Phytophthora and the Species Phytophthora nicotianae by LAMP with a QProbe. Plant Dis. 2020, 104, 2469–2480. [Google Scholar] [CrossRef] [PubMed]
- De Bary, H.A. Researches into the nature of the potato–fungus, Phytophthora infestans. J. Roy. Agric. Soc. 1876, 12, 239–269. Available online: https://BiotaNZ.landcareresearch.co.nz/references/1cb0e861-36b9-11d5-9548-00d0592d548c (accessed on 13 May 2023).
- Yang, X.; Tyler, B.M.; Hong, C.X. An expanded phylogeny for the genus Phytophthora. IMA Fungus 2017, 8, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Scanu, B.; Bakonyi, J.; Seress, D.; Kovács, G.; Durán, A.; von Stowasser, E.S.; Schena, L.; Mosca, S.; Thu, P.; et al. Nothophytophthora gen. nov. a new sister genus of Phytophthora from natural and semi-natural ecosystem. Pers. -Mol. Phylogeny Evol. Fungi 2017, 39, 143–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Bakhshi, M.; Balci, Y.; Broders, K.; Cheewangkoon, R.; Chen, S.; Fan, X.; Gramaje, D.; Halleen, F.; Jung, M.H.; et al. Genera of phytopathogenic fungi: GOPHY 4. Stud. Mycol. 2022, 101, 417–564. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; American Phytopathological Society (APS) Press: St. Paul, MN, USA, 1996; p. 592. [Google Scholar]
- Gallegly, M.E.; Hong, C.X. Phytophthora: Identifying Species by Morphology and DNA Fingerprints. J. Phytopathol. 2008, 157, 520. [Google Scholar] [CrossRef]
- Van Tran, Q.; Ha, C.V.; Vvedensky, V.V.; Linh Le, T.T.; Han, V.C. Pathogenicity and fungicide sensitivity of Phytophthora parvispora, a new pathogen causing gummosis and root rot disease on citrus trees. Microb. Pathog. 2023, 175, 105986. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.H. The taxonomy and biology of Phytophthora and Pythium. J. Bacteriol. Mycol. 2018, 6, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Agrios, G.N. Plant Pathology, 5th ed.; Academic Press: Cambridge, MA, USA, 2005; p. 922. [Google Scholar]
- Fernandes, P.; Colavolpe, M.B.; Serrazina, S.; Costa, R.L. European and American chestnuts: An overview of the main threats and control efforts. Front. Plant Sci. 2022, 13, 844–951. [Google Scholar] [CrossRef] [PubMed]
- Colavolpe, M.B.; Vaz Dias, F.; Serrazina, S.; Malhó, R.; Lourenço Costa, R. Castanea crenata Ginkbilobin-2-like Recombinant Protein Reveals Potential as an Antimicrobial against Phytophthora cinnamomi, the Causal Agent of Ink Disease in European Chestnut. Forests 2023, 14, 785. [Google Scholar] [CrossRef]
- Jung, T.; Vettraino, A.M.; Cech, T.; Vannini, A. The impact of invasive Phytophthora species on European forests. In Phytophthora: A Global Perspective; Lamour, K., Ed.; Plant Protection Series 2; CABI: Wallingford, UK, 2013; pp. 146–158. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez–Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and Pathogenicity of Phytophthora Species from Poplar Plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Sun, S.; Yang, Y.; Zhou, B.; Duan, C.; Shan, W.; Zhu, Z. A Novel Disease of Mung Bean, Phytophthora Stem Rot Caused by a New Forma Specialis of Phytophthora vignae. Plant Dis. 2021, 105, 2160–2168. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Burgess, T.; Dau, V.T.; Le, V.Q.; Trinh, T.L.; Pham, T.L.; Burgess, L.W. Phytophthora stem rot of purple passionfruit in Vietnam. Australas. Plant Dis. Notes 2015, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Browne, G.T.; Ott, N.J.; Forbes, H.; Yaghmour, M.; Milliron, L.K. First report of Phytophthora chlamydospora causing crown and root rot on almond in California. Plant Dis. 2020, 104, 2033. [Google Scholar] [CrossRef] [Green Version]
- Abuley, I.K.; Hansen, J.G. An epidemiological analysis of the dilemma of plant age and late blight (Phytophthora infestans) susceptibility in potatoes. Eur. J. Plant Pathol. 2021, 161, 645–663. [Google Scholar] [CrossRef]
- Chang, L.; Liu, K.; Wang, L.G.; Yang, X.L.; Sun, H. First report of Phytophthora nicotianae causing stem canker of Catalpa bungei (Chinese catalpa) in China. Plant Dis. 2021, 106, 1309. [Google Scholar] [CrossRef]
- Elliott, M.; Rollins, L.; Bourret, T.; Chastagner, G. First report of leaf blight caused by Phytophthora ramorum on periwinkle (Vinca minor) in Washington State, USA. Plant Dis. 2021, 105, 2023. [Google Scholar] [CrossRef]
- Merga, J.; Shamil, A. Characterization of Phytophthora capsici Foot Rot Disease of Black Pepper in Ethiopia. J. Plant Pathol. Microbiol. 2021, 12, 537. [Google Scholar]
- Pane, A.; Li Destri Nicosia, M.G.; Cacciola, S.O. First Report of Phytophthora citrophthora Causing Fruit Brown Rot of Feijoa in Italy. Plant Dis. 2001, 85, 97. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Favaron, F.; Sella, L. First report of Phytophthora acerina, P. pini, and P. plurivora causing root rot and sudden death of olive trees in Italy. Plant Dis. 2020, 104, 996. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Hrabetova, M.; Mrazkova, M.; Cerny, K. First Report of Phytophthora occultans Causing Dieback of Buxus sempervirens in the Czech Republic. Plant Dis. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Gregory, P.H. Some major epidemics caused by Phytophthora. In Phytophthora: Its Biology, Taxonomy, Ecology, and Pathology; Erwin, D.C., Bartnicki-Garcia, S., Eds.; American Phytopathological Society (APS) Press: St. Paul, MN, USA, 1983; pp. 271–278. [Google Scholar]
- Werres, S.; Marwitz, R.; In’T Veld, W.A.M.; De Cock, A.W.A.M.; Bonants, P.J.M.; De Weerdt, M.; Themann, K.; Ilieva, E.; Baayen, R.P. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and Viburnum. Mycol. Res. 2001, 105, 1155–1165. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M.; Davidson, J.M.; Slaughter, G.W.; Koike, S.T. Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 2002, 86, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Huo, X. Pathogen Identification and Fungicides Screening of Bletilla Striata Blight; Guangxi University: Nanning, China, 2019; Volume 1, p. 82. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1019215854.nh&DbName=CMFD2020 (accessed on 15 May 2023).
- Lan, C.Z.; Ruan, H.C.; Yao, J.A. First report of Phytophthora cinnamomi causing root and stem rot of blueberry (Vaccinium corymbosum) in China. Plant Dis. 2016, 100, 2537. [Google Scholar] [CrossRef]
- Lan, C.Z.; Ruan, H.C.; Yao, J.A. First report of Phytophthora cinnamomi causing root rot of Castanea mollissima (Chinese chestnut) in China. Plant Dis. 2016, 100, 1248. [Google Scholar] [CrossRef]
- Zheng, X.B.; Lu, J.Y. Studies on Phytophthora species in Fujian, Zhejiang, Jiangsu Provinces and Shanghai, China. Acta Mycol. Sin. 1989, 8, 161–168. [Google Scholar] [CrossRef]
- Zeng, H.C.; Ho, H.H.; Zheng, F.C. A survey of Phytophthora species on Hainan Island of South China. J. Phytopathol. 2009, 157, 33–39. [Google Scholar] [CrossRef]
- Bi, X.Q.; Hieno, A.; Otsubo, K.; Kageyama, K.; Liu, G.; Li, M.Z. A multiplex PCR assay for three pathogenic Phytophthora species related to kiwifruit diseases in China. J. Gen. Plant Pathol. 2019, 85, 12–22. [Google Scholar] [CrossRef]
- Zhou, X.G.; Zhu, Z.Y.; Lu, C.P.; Wang, S.J.; Ko, W.H. Phytophthora cinnamomi in Shanghai and it possible origin. Mycopathologia 1992, 120, 29–32. [Google Scholar] [CrossRef]
- Dai, T.; Wang, A.; Yang, X.; Yu, X.; Tian, W.; Xu, Y.; Hu, T. PHYCI_587572: An RxLR Effector Gene and New Biomarker in A Recombinase Polymerase Amplification Assay for Rapid Detection of Phytophthora cinnamomi. Forests 2020, 11, 306. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Zheng, X.B. Phytophthora and Methods in Phytophthoa; China Agriculture Press: Beiing, China, 1997. (In Chinese) [Google Scholar]
- Martin, F.N.; Abad, Z.G.; Balci, Y.; Ivors, K. Identification and Detection of Phytophthora: Reviewing Our Progress, Identifying Our Needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.E.; Coffey, M.D.; Park, S.-Y.; Geiser, D.M.; Kang, S. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genet. Biol. 2008, 45, 266–277. [Google Scholar] [CrossRef]
- Kroon, L.P.N.M.; Bakker, F.T.; van den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylodenetic analsis of Phytophthora species based on mitochondrial and nuclear DNA seouens. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Chen, Z.P.; Yang, X.; Xue, J.X.; Jiao, B.B.; Li, Y.X.; Xu, Y.; Dai, T.T. First Report of Phytopythium helicoides Causing Crown and Root Rot on Rhododendron pulchrum in China. Plant Dis. 2021, 105, 713. [Google Scholar] [CrossRef]
- dos Santos, A.F.; Luz, E.D.M.N.; de Souza, J.T. Phytophthora nicotianae: Agente Etiologico da Gomose da Acacia-Negra no Brasil I. Fitopatol. Bras. 2005, 30, 81–83. [Google Scholar] [CrossRef]
- Sun, D.W.; Cao, J.F.; Pei, W.H.; Yin, G.F.; Ma, J.Q.; Pan, K.H.; Wu, K.; Zhao, Q.; Duan, Z.; Yang, M.; et al. ldentification of a new Phytophthora blight disease on konjac in Yunnan. Phytopathol. Res. 2015, 45, 84–87. [Google Scholar] [CrossRef]
- Damm, U.; Mostert, L.; Crous, P.W.; Fourie, P.H. Novel Phaeoacremonium species associated with necrotic wood of Prunus trees. Persoonia 2008, 20, 87–102. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Martin, F.N.; and Tooley, P.W. Phylogenetic Relationships among Phytophthora Species Inferred from Sequence Analysis of Mitochondrially Encoded Cytochrome Oxidase I and II Genes. Mycologia 2003, 95, 269–284. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/ nt. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovli´c, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.L. Study on Scab-anthranoge of Fatsia japonica. J. Zhejiang For. Sci. Technol. 2007, 27, 64–67. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=ZJLK200705016&DbName=CJFQ2007 (accessed on 15 May 2023).
- Shi, N.N.; Du, Y.X.; Chen, F.R.; Ruan, H.C.; Yang, X.J. First report of leaf spot caused by Colletotrichum fructicola on Japanese Fatsia (Fatsia japonica) in Fujian Province in China. Plant Dis. 2017, 101, 1552. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, S.B.; Wang, Y.H.; Lin, Q.K.; Zhou, Z. First Report of Botryosphaeria dothidea Causing a Leaf Wilt on Fatsia japonica in Henan Province, China. Plant Dis. 2018, 102, 450. [Google Scholar] [CrossRef]
- Mehrabi-Koushki, M.; Artand, S.; Ahmadpour, S.A. Botryosphaeria dothidea causes stem canker on Fatsia japonica in Iran. Australas. Plant Dis. Notes 2021, 16, 31. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gilardi, G.; Gullino, M.L. First Report of Alternaria Leaf Blight of Aralia japonica Caused by Alternaria panax in Europe. Plant Dis. 2004, 88, 82. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Park, M.S.; Yu, S.H. Molecular characterization, morphology, and pathogenicity of Alternaria panax from araliaceous plants in Korea. Mycol. Prog. 2013, 12, 383–396. [Google Scholar] [CrossRef]
- Panabieres, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.-L.; Roy, S.G.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognised pathogen. Phytopathol. Mediterr. 2016, 55, 20–40. [Google Scholar] [CrossRef]
- Cline, E.T.; Farr, D.F.; Rossman, A.Y. A synopsis of Phytophthora with accurate scientific names, host range, and geographic distribution. Plant Health Prog. 2008, 9, 32. [Google Scholar] [CrossRef]
- Falcón, A.B.; Cabrera, J.C.; Costales, D.; Ramírez, M.A.; Cabrera, G.; Toledo, V.; Martínez-Téllez, M.A. The effect of size and acetylation degree of chitosan derivatives on tobacco plant protection against Phytophthora parasitica nicotianae. World J. Microbiol. Biotechnol. 2008, 24, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Li, D.L.; Huo, C.; Cao, J.F.; Rajput, N.A.; Zhao, Z.J. First Report of Phytophthora nicotianae Causing Asparagus Spear and Root Rot in China. Plant Dis. 2018, 102, 1042. [Google Scholar] [CrossRef]
- Yu, X.L.; Liu, X.Q.; Wang, P.S.; Wang, Y.Z. First Report of Cherry Stem Rot and Leaf Necrosis Disease Caused by Phytophthora nicotianae in Yantai, China. Plant Dis. 2015, 99, 284. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.V.; Seijo, T.E.; Zuchelli, E.; Peres, N.A. Resistance to Mefenoxam of Phytophthora cactorum and Phytophthora nicotianae Causing Crown and Leather Rot in Florida Strawberry. Plant Dis. 2021, 105, 3490–3495. [Google Scholar] [CrossRef]
- Chaudhary, S.; Laughlin, D.A.; Setamou, M.; da Graça, J.V.; Kunta, M.; Alabi, O.J.; Crosby, K.M.; Ong, K.L.; Ancona, V. Incidence, Severity, and Characterization of Phytophthora Foot Rot of Citrus in Texas and Implications for Disease Management. Plant Dis. 2020, 104, 2455–2461. [Google Scholar] [CrossRef]
- Xu, J.Y.; Yang, X.; Wu, C.P.; Zhou, Z.W.; Chen, Z.P.; Dai, T.T. First Report of Phytophthora nicotianae Causing Dianthus chinensis Root Rot and Foliage Blight in China. Plant Dis. 2022, 106, 2002. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Forster, H.; Belisle, R.J.; Adaskaveg, J. New Preharvest Treatments and Strategies in Managing Phytophthora Brown Rot of Citrus in California. Plant Dis. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- El Meleigi, M.A.; Omar, A.F.; Al Rokibah, A.A.; Alsohim, A.; Al Jamhan, K.A.; Sukar, N.A. Molecular identification andpathogenicity of Phytophthora nicotianae caused bud rot disease of Washingtonia palms in Saudi Arabia and use of Lysobacter enzymogenes, as abioagent in an in vitro study. Egypt. J. Biol. Pest. Control. 2019, 29, 3. [Google Scholar] [CrossRef]
- Farhana, L.; Nangia-Makker, P.; Arbit, E.; Shango, K.; Sarkar, S.; Mahmud, H.; Hadden, T.; Yu, Y.; Majumdar, A.P. Bile acid: A potential inducer of colon cancer stem cells. Stem Cell. Res. Ther. 2016, 7, 181. [Google Scholar] [CrossRef] [Green Version]
- Aloi, F.; Riolo, M.; La Spada, F.; Bentivenga, G.; Moricca, S.; Santilli, E.; Pane, A.; Faedda, R.; Cacciola, S.O. Phytophthora Rootand Collar Rot of Paulownia, a New Disease for Europe. Forests 2021, 12, 1664. [Google Scholar] [CrossRef]
- Aloi, F.; Parlascino, R.; ContiTaguali, S.; Faedda, R.; Pane, A.; Cacciola, S.O. Phytophthora pseudocryptogea, P. nicotianae and P. multivora Associated to Cycas revoluta: First Report World wide. Plants 2023, 12, 1197. [Google Scholar] [CrossRef] [PubMed]




| Locus | Primer | Sequence (5′-3′) | PCR Conditions | Reference |
|---|---|---|---|---|
| The internal transcribed spacer (ITS) | ITS1 | TCCGTAGGTGAACCTGCGG | 94 °C, 3 min; (94 °C, 30 s, 55 °C, 30 s; 72 °C, 45 s) × 35; 72 °C, 10 min | [53] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| Large subunit (LSU) | LROR-O | ACCCGCTGAACTYAAGC | 94 °C, 3 min; (94 °C, 30 s; 52 °C, 30 s; 72 °C, 45 s) × 35; 72 °C, 10 min | [46] |
| LR6-O | CGCCAGACGAGCTTACC | |||
| Mitochondrially encoded cytochrome oxidase II (COXII) | FM82 | TTGGCAATTAGGTTTTCAAGATCC | 94 °C, 3 min; (94 °C, 30 s; 52 °C, 30 s; 72 °C, 45 s) × 35; 72 °C, 10 min | [54] |
| FM80 | AATATCTTTATGATTTGTTGAAA |
| Phytophthora Species | Isolate | GenBank Accession Numbers 1 | ||
|---|---|---|---|---|
| ITS | LSU | COXII | ||
| P. nicotianae | P6303 | JN699566 | EU080603 | GU318304 |
| P. nicotianae | dc3 | MZ557793 | MZ573546 | MZ573545 |
| P. nicotianae | dc7 | MZ519893 | MZ573547 | MZ540768 |
| P. nicotianae | dc8 | MZ557794 | MZ573549 | MZ573548 |
| P. cactorum | P0714 | HQ261514 | EU080282 | GU221951 |
| P. hedraiandra | P11056 | FJ802065 | EU080077 | JF771449 |
| P. idaei | P6767 | HQ261579 | EU080134 | GU222032 |
| P. pseudotsugae | P10339 | HQ261654 | EU080431 | GU222121 |
| P. clandestina | P3942 | HQ261538 | EU079871 | GU221981 |
| P. iranica | P3882 | HQ261598 | EU080116 | GU222048 |
| P. tentaculata | P8497 | HQ261717 | EU079960 | GU222150 |
| P. andina | P13365 | FJ801734 | EU080187 | GU318297 |
| P13660 | FJ801748 | - | GU221934 | |
| P13766 | FJ801753 | - | JQ439407 | |
| P. infestans | P10650 | HQ261589 | EU079630 | GU318302 |
| P11633 | FJ802075 | - | JF771479 | |
| P12021 | GU258555 | - | JF771480 | |
| P. ipomoeae | P10225 | HQ261597 | EU080835 | GU222045 |
| P10226 | HQ261596 | EU080842 | GU222046 | |
| P10227 | HQ261595 | EU080849 | GU222047 | |
| P. mirabilis | P3005 | HQ261622 | EU079780 | GU222077 |
| P. phaseoli | P6609 | HQ261640, | EU079918 | GU222106 |
| P10145 | HQ261642 | EU080753 | GU222104 | |
| P10150 | HQ261641 | EU080766 | GU222105 | |
| P. capsici | P0253 | FJ801244 | EU080856 | GU318299 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Xu, T.; Xu, X.; Dai, T.; Liu, T. The New Report of Root Rot on Fatsia japonica Caused by Phytophthora nicotianae in China. Forests 2023, 14, 1459. https://doi.org/10.3390/f14071459
Zhou J, Xu T, Xu X, Dai T, Liu T. The New Report of Root Rot on Fatsia japonica Caused by Phytophthora nicotianae in China. Forests. 2023; 14(7):1459. https://doi.org/10.3390/f14071459
Chicago/Turabian StyleZhou, Jing, Tingyan Xu, Xiaoqiao Xu, Tingting Dai, and Tingli Liu. 2023. "The New Report of Root Rot on Fatsia japonica Caused by Phytophthora nicotianae in China" Forests 14, no. 7: 1459. https://doi.org/10.3390/f14071459