
Effect of Stand Density on Soil Organic Carbon Storage and Extracellular Enzymes Activity of Larch Plantation in Northeast China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Conditions
2.2. Sampling
2.3. Incubation
2.4. Physical and Chemical Analysis
2.5. Extracellular Enzymes
3. Results
3.1. Soil Properties
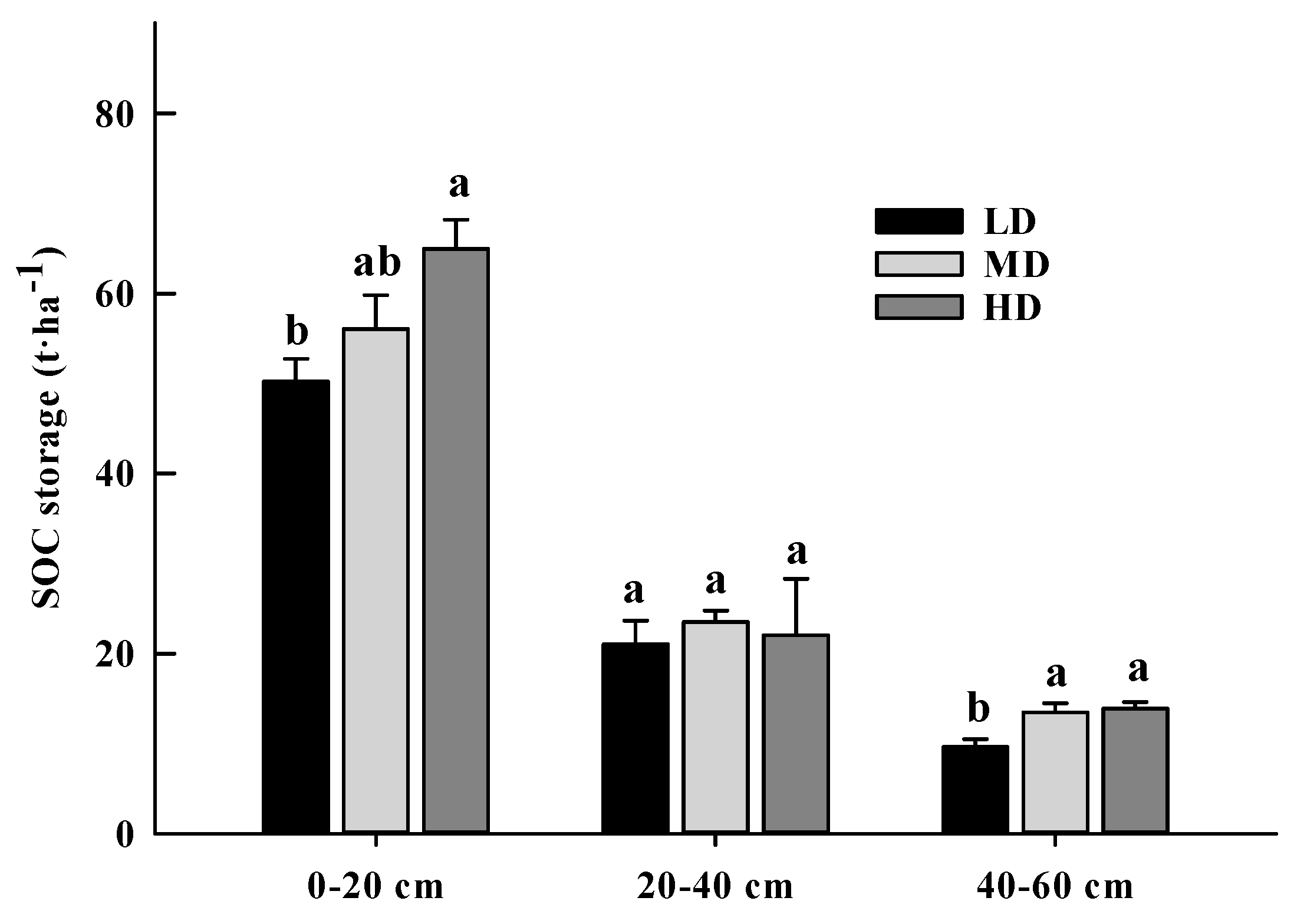
3.2. SOC Storage and Its Allocation in Light Fraction and Heavy Fraction
3.3. Emission of 14C-CO2 and SOC Derived CO2
3.4. Extracellular Enzymes Activity
4. Discussion
4.1. SOC Storage Changes with Stand Density
4.2. The Explanations for SOC Storage Change
4.3. Suppression of Microbes
4.4. Microbial Suppression Attributions to Soil C Storage
4.5. Limitation of the Study
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frouz, J.; Pižl, V.; Cienciala, E.; Kalčík, J. Carbon storage in post-mining forest soil, the role of tree biomass and soil bioturbation. Biogeochemistry 2009, 94, 111–121. [Google Scholar] [CrossRef]
- Karhu, K.; Wall, A.; Vanhala, P.; Liski, J.; Esala, M.; Regina, K. Effects of afforestation and deforestation on boreal soil carbon stocks-Comparison of measured C stocks with Yasso07 model results. Geoderma 2011, 164, 33–34. [Google Scholar]
- Li, D.; Niu, S.; Luo, Y. Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: A meta-analysis. New Phytol. 2012, 195, 172–181. [Google Scholar]
- Litton, C.M.; Ryan, M.G.; Knight, D.H.; Stahl, P.D. Soil-surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem. Glob. Change Biol. 2003, 9, 680–696. [Google Scholar]
- Scott, N.A.; Tate, K.R.; Ross, D.J.; Parshotam, A. Processes influencing soil carbon storage following afforestation of pasture with Pinus radiata at different stocking densities in New Zealand. Aust. J. Soil Res. 2006, 44, 85–96. [Google Scholar]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar]
- Fernández-Núñez, E.; Rigueiro-Rodríguez, A.; Mosquera-Losada, M.R. Carbon allocation dynamics one decade after afforestation with Pinus radiata D. Don and Betula alba L. under two stand densities in NW Spain. Ecol. Eng. 2010, 36, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Noh, N.J.; Son, Y.; Lee, S.K.; Yoon, T.K.; Seo, K.W.; Kim, C.; Lee, W.-K.; Bae, S.W.; Hwang, J. Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. J. Plant Res. 2010, 123, 411–419. [Google Scholar] [CrossRef]
- González, I.G.; Corbí, J.M.G.; Cancio, A.F.; Ballesta, R.J.; Cascón, M.R.G. Soil carbon stocks and soil solution chemistry in Quercus ilex stands in Mainland Spain. Eur. J. For. Res. 2012, 131, 1653–1667. [Google Scholar]
- Sitters, J.; Edwards, P.J.; Venterink, H.O. Increases of Soil C, N, and P Pools Along an Acacia Tree Density Gradient and Their Effects on Trees and Grasses. Ecosystems 2013, 16, 347–357. [Google Scholar]
- Sun, X.; Wang, W.; Razaq, M.; Sun, H. Effects of stand density on soil organic carbon storage in the top and deep soil layers of Fraxinus mandshurica plantations. Austrian J. For. Sci. 2019, 136, 27–44. [Google Scholar]
- Gao, Y.; Dang, P.; Zhao, Z. Effects of afforestation on soil carbon and its fractions: A case study from the Loess Plateau, China. J. For. Res. 2018, 29, 1291–1297. [Google Scholar] [CrossRef]
- Ahmad, B.; Wang, Y.; Hao, J.; Liu, Y.; Bohnett, E.; Zhang, K. Variation of carbon density components with overstory structure of larch plantations in northwest China and its implication for optimal forest management. For. Ecol. Manag. 2021, 496, 119399. [Google Scholar]
- Kirkby, C.A.; Kirkegaard, J.A.; Richardson, A.E.; Wade, L.J.; Blanchard, C.; Batten, G. Stable soil organic matter: A comparison of C:N:P:S ratios in Australian and other world soils. Geoderma 2011, 163, 197–208. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting Density and Site Effects on Stem Dimensions, Stand Productivity, Biomass Partitioning, Carbon Stocks and Soil Nutrient Supply in Hybrid Poplar Plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef] [Green Version]
- Litton, C.; Ryan, M.; Knight, D. Effects of tree density and stand age on carbon allocation patterns in postfire lodgepole pine. Ecol. Appl. 2004, 14, 460–475. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Xue, J.; Tang, L. Biomass production and carbon sequestration potential in poplar plantations with different management patterns. J. Environ. Manage. 2007, 85, 672–679. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2004, 7, 314–320. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S. Size and functional diversity of microbe populations control plant persistence and long-term soil carbon accumulation. Ecol. Lett. 2005, 8, 1075–1087. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Noh, N.J.; Kim, C.; Bae, S.W.; Lee, W.K.; Yoon, T.K.; Muraoka, H.; Son, Y. Carbon and nitrogen dynamics in a Pinus densiflora forest with low and high stand densities. J. Plant. Ecol. 2013, 6, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, S.; Bardoux, G.; Benest, D.; Verdier, B.; Mariotti, A.; Abbadie, L. Mechanisms of the priming effect in a savannah soil amended with cellulose. Soil Sci. Soc. Am. J. 2004, 68, 125–131. [Google Scholar]
- Averill, C.; Waring, B. Nitrogen limitation of decomposition and decay: How can it occur? Glob. Change Biol. 2018, 24, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowden, R.D.; Davidsonb, E.; Savageb, K.; Arabiaa, C.; Steudlerc, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.; Gu, J.; Xu, S.; Yu, L.; Wang, Z. Effects of continuous nitrogen addition on microbial properties and soil organic matter in a Larix gmelinii plantation in China. J. For. Res. 2018, 29, 85–92. [Google Scholar] [CrossRef]
- Kögel-Knabner, I. The macromolecular organic composition of Plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Fontaine, S.; Henault, C.; Aamor, A.; Bdioui, N.; Bloor, J.M.G.; Maire, V.; Mary, B.; Revaillot, S.; Maron, P.A. Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect. Soil Biol. Biochem. 2011, 43, 86–96. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.; Manter, D.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar]
- Luo, Y.; Zhao, X.; Andrén, O.; Zhu, Y.; Huang, W. Artificial root exudates and soil organic carbon mineralization in a degraded sandy grassland in northern China. J. Arid Land 2014, 6, 423–431. [Google Scholar] [CrossRef]
- Badri, D.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Salahuddin, L.J.; Zhang, J.; You, L.; He, C.; Yang, L. Larch (Larix gmelinii) bulk soil phenolic acids promote manchurian walnut (Juglans manshurica) growth and soil microorganism biomass. Pak. J. Bot. 2016, 48, 2549–2556. [Google Scholar]
- Zhu, X.; Liu, W.; Chen, H.; Deng, Y.; Chunfeng, C.; Zeng, H. Effects of forest transition on litterfall, standing litter and related nutrient returns: Implications for forest management in tropical China. Geoderma 2019, 333, 123–134. [Google Scholar] [CrossRef]
- Anyanwu, I.; Semple, K. Assessment of the effects of phenanthrene and its nitrogen heterocyclic analogues on microbial activity in soil. SpringerPlus 2016, 5, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushio, M.; Miki, T.; Kitayama, K. Phenolic Control of Plant Nitrogen Acquisition through the Inhibition of Soil Microbial Decomposition Processes: A Plant-Microbe Competition Model. Microbes Environ. 2009, 24, 180–187. [Google Scholar] [CrossRef]
- Bagchi, S.; Roy, S.; Maitra, A.; Rubanpreet, S.S. Herbivores suppress soil microbes to influence carbon sequestration in the grazing ecosystem of the Trans-Himalaya. Agric. Ecosyst. Environ. 2017, 239, 199–206. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Deep soil organic matter-a key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barre, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef]
- Iversen, C.; Keller, J.K.; Garten, C.T.; Norby, R. Soil carbon and nitrogen cycling and storage throughout the soil profile in a sweetgum plantation after 11 years of CO2-enrichment. Glob. Change Biol. 2012, 18, 1684–1697. [Google Scholar] [CrossRef]
- Salomé, C.; Nunan, N.; Pouteau, V.; Lerch, T.Z.; Chenu, C. Carbon dynamics in topsoil and in subsoil may be controlled by different regulatory mechanisms. Glob. Change Biol. 2010, 16, 416–426. [Google Scholar]
- Sinsabaugh, R.S. Enzymic analysis of microbial pattern and process. Biol. Fertil. Soils 1994, 17, 69–74. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar]
- Allison, S.D.; Weintraub, M.N.; Gartner, T.B.; Waldrop, M.P. Evolutionary-Economic Principles as Regulators of Soil Enzyme Production and Ecosystem Function. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 229–243. [Google Scholar]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar]
- Arunrat, N.; Sereenonchai, S.; Sansupa, C.; Kongsurakan, P.; Hatano, R. Effect of Rice Straw and Stubble Burning on Soil Physicochemical Properties and Bacterial Communities in Central Thailand. Biology 2023, 12, 501. [Google Scholar]
- Hernández, D.L.; Hobbie, S.E. The effects of substrate composition, quantity, and diversity on microbial activity. Plant Soil 2010, 335, 397–411. [Google Scholar] [CrossRef]
- Brzostek, E.; Finzi, A.C. Substrate supply, fine roots, and temperature control proteolytic enzyme activity in temperate forest soils. Ecology 2011, 92, 892–902. [Google Scholar] [CrossRef]
- Brackin, R.; Robinson, N.; Lakshmanan, P.; Schmidt, S. Soil microbial responses to labile carbon input differ in adjacent sugarcane and forest soils. Soil Res. 2014, 52, 307. [Google Scholar] [CrossRef]
- Heitkötter, J.; Niebuhr, J.; Heinze, S.; Marschner, B. Patterns of nitrogen and citric acid induced changes in C-turnover and enzyme activities are different in topsoil and subsoils of a sandy Cambisol. Geoderma 2017, 292, 111–117. [Google Scholar]
- Keeler, B.; Hobbie, S.; Kellogg, L. Effects of Long-Term Nitrogen Addition on Microbial Enzyme Activity in Eight Forested and Grassland Sites: Implications for Litter and Soil Organic Matter Decomposition. Ecosystems 2008, 12, 1–15. [Google Scholar] [CrossRef]
- Riggs, C.; Hobbie, S. Mechanisms driving the soil organic matter decomposition response to nitrogen enrichment in grassland soils. Soil Biol. Biochem. 2016, 99, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar]
- Heinze, S.; Ludwig, B.; Piepho, H.-P.; Mikutta, R.; Don, A.; Wordell-Dietrich, P.; Helfrich, M.; Hertel, D.; Leuschner, C.; Kirfel, K.; et al. Factors controlling the variability of organic matter in the top- and subsoil of a sandy Dystric Cambisol under beech forest. Geoderma 2018, 311, 37–44. [Google Scholar] [CrossRef]
- Vance, E.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Marx, M.C.; Wood, M.; Jarvis, S.C. A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem. 2001, 33, 1633–1640. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; Passioura, J.B.; Batten, G.D.; Blanchard, C.; Kirkegaard, J.A. Nutrient availability limits carbon sequestration in arable soils. Soil Biol. Biochem. 2014, 68, 402–409. [Google Scholar] [CrossRef]
- DeMarco, J.; Mack, M.C.; Bret-Harte, M.S. Effects of arctic shrub expansion on biophysical vs. biogeochemical drivers of litter decomposition. Ecology 2014, 95, 1861–1875. [Google Scholar] [CrossRef] [PubMed]
- de Graaff, M.A.; Jastrow, J.D.; Gillette, S.; Johns, A.; Wullschleger, S.D. Differential priming of soil carbon driven by soil depth and root impacts on carbon availability. Soil Biol. Biochem. 2014, 69, 147–156. [Google Scholar] [CrossRef]
- Czarnota, M.A.; Rimando, A.M.; Weston, L.A. Evaluation of Root Exudates of Seven Sorghum Accessions. J. Chem. Ecol. 2003, 29, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Klumpp, K.; Fontaine, S.; Attard, E.; Le Roux, X.; Gleixner, G.; Soussana, J.-F. Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community. J. Ecol. 2009, 97, 876–885. [Google Scholar] [CrossRef]
- Shahzad, T.; Chenu, C.; Repincay, C.; Mougin, C.; Ollier, J.-L.; Fontaine, S. Plant clipping decelerates the mineralization of recalcitrant soil organic matter under multiple grassland species. Soil Biol. Biochem. 2012, 51, 73–80. [Google Scholar] [CrossRef]
- Dawson-Glass, E.; Hewins, C.R.; Burke, D.J.; Stuble, K.L. Experimental increases in pH and P availability exert long-term impacts on decomposition in forests. Appl. Soil Ecol. 2023, 181, 104654. [Google Scholar] [CrossRef]
- Stott, D.E.; Andrews, S.S.; Liebig, M.A.; Wienhold, B.J.; Karlen, D.L. Evaluation of β-Glucosidase Activity as a Soil Quality Indicator for the Soil Management Assessment Framework. Soil Sci. Soc. Am. J. 2010, 74, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wu, Z.; Zong, Y.; Li, W.; Chen, F.; Wang, G.G.; Li, J.; Fang, X. Mixing with coniferous tree species alleviates rhizosphere soil phosphorus limitation of broad-leaved trees in subtropical plantations. Soil Biol. Biochem. 2022, 175, 108853. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.; Simpson, R.; Moore, J. Carbon-degrading enzyme activities stimulated by increased nutrient availability in arctic tundra soils. PLoS ONE 2013, 8, e77212. [Google Scholar] [CrossRef] [Green Version]
- Stark, S.; Männistö, M.; Eskelinen, A. Nutrient availability and pH jointly constrain microbial extracellular enzyme activities in nutrient-poor tundra soils. Plant Soil 2014, 383, 373–385. [Google Scholar] [CrossRef]
- Creamer, C.A.; Jones, D.L.; Baldock, J.A.; Rui, Y.; Murphy, D.V.; Hoyle, F.C.; Farrell, M. Is the fate of glucose-derived carbon more strongly driven by nutrient availability, soil texture, or microbial biomass size? Soil Biol. Biochem. 2016, 103, 201–212. [Google Scholar] [CrossRef]
- Hüblová, L.; Frouz, J. Contrasting effect of coniferous and broadleaf trees on soil carbon storage during reforestation of forest soils and afforestation of agricultural and post-mining soils. J. Environ. Manage. 2021, 290, 112567. [Google Scholar] [CrossRef]
- Wäldchen, J.; Schöning, I.; Mund, M.; Schrumpf, M.; Bock, S.; Herold, N.; Totsche, K.; Ernst Detlef, S. Estimation of clay content from easily measurable water content of air-dried soil. J. Plant. Nutr. Soil Sci. 2012, 175, 367–376. [Google Scholar] [CrossRef]
- Dilustro, J.J.; Collins, B.; Duncan, L.; Crawford, C. Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests. For. Ecol. Manag. 2005, 204, 87–97. [Google Scholar] [CrossRef]
- Müller, T.; Höper, H. Soil organic matter turnover as a function of the soil clay content: Consequences for model applications. Soil Biol. Biochem. 2004, 36, 877–888. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, Z.; Yang, K.; Zhu, J. Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities. Appl. Soil Ecol. 2017, 111, 73–83. [Google Scholar] [CrossRef]
- Berger, T.W.; Neubauer, C.; Glatzel, G. Factors controlling soil carbon and nitrogen stores in pure stands of Norway spruce (Picea abies) and mixed species stands in Austria. For. Ecol. Manag. 2002, 159, 3–14. [Google Scholar] [CrossRef]
- Iversen, C.; Ledford, J.; Norby, R. CO2 enrichment increases carbon and nitrogen input from fine roots in a deciduous forest. New Phytol. 2008, 179, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zeng, D.H.; Ma, X.Q.; Chang, S. Root rather than leaf litter input drives soil carbon sequestration after afforestation on a marginal cropland. For. Ecol. Manag. 2016, 362, 38–45. [Google Scholar] [CrossRef]
- Ding, Y.; Leppälammi-Kujansuu, J.; Helmisaari, H.S. Fine root longevity and below- and aboveground litter production in a boreal Betula pendula forest. For. Ecol. Manag. 2019, 431, 17–25. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer | Density | Fine Root Biomass (g m−2) | C (mg g−1) | N (mg g−1) | C/N | Cmic (μg g−1) | Nmin (μg g−1) | Nmin/Cmic (10−2) | DOC (μg g−1) | pH |
|---|---|---|---|---|---|---|---|---|---|---|
| 0–20 cm | LD | 128.98 b (28.03) | 31.21 a (6.48) | 3.04 a (0.59) | 10.20 a (0.51) | 230.48 a (10.77) | 18.56 a (1.98) | 8.05 b (1.32) | 51.41 c (3.14) | 5.01 a (0.06) |
| MD | 165.53 ab (23.36) | 32.07 a (5.88) | 3.04 a (0.53) | 10.55 a (0.41) | 189.50 b (25.83) | 19.56 a (2.14) | 10.84 b (2.19) | 59.36 b (2.91) | 5.29 a (0.14) | |
| HD | 187.16 a (26.18) | 30.44 a (5.70) | 2.88 a (0.48) | 10.50 a 0.24) | 128.32 c (15.92) | 19.40 a (2.70) | 15.12 a (3.06) | 73.66 a (4.31) | 5.20 a (0.06) | |
| 20–40 cm | LD | 43.96 b (9.74) | 10.62 a (1.84) | 1.11 a (0.17) | 9.51 a (0.44) | 39.04 a (3.51) | 17.14 a (1.42) | 43.90 c (4.12) | 44.53 b (3.37) | 4.79 a (0.13) |
| MD | 49.96 ab (11.43) | 11.33 a (1.92) | 1.15 a (0.10) | 9.74 a (0.82) | 30.32 b (2.18) | 15.84 a (2.07) | 52.24 b (6.18) | 50.59 b (2.41) | 5.00 a (0.13) | |
| HD | 69.98 a (16.57) | 7.81 a (2.53) | 0.83 a (0.27) | 9.43 a (0.21) | 26.27 b (1.83) | 16.94 a (0.95) | 64.48 a (5.60) | 57.38 a (1.86) | 5.01 a (0.03) | |
| 40–60 cm | LD | 12.03 b (5.84) | 3.63 a (0.57) | 0.42 a (0.05) | 8.69 b (0.26) | 9.22 a (0.70) | 3.21 a (0.35) | 34.82 c (3.53) | 33.60 b (4.19) | 4.76 a (0.16) |
| MD | 14.37 b (4.26) | 5.99 a (1.81) | 0.68 a (0.20) | 8.73 b (0.67) | 8.95 a (0.71) | 4.70 a (0.39) | 52.51 b (5.81) | 34.13 b (3.79) | 5.06 a (0.10) | |
| HD | 22.11 a (5.22) | 5.86 a (0.87) | 0.61 a (0.13) | 9.87 a (0.80) | 6.38 b (0.57) | 4.99 a (0.53) | 78.21 a (8.38) | 60.43 a (9.71) | 5.07 a (0.03) |
| Layer | Treatment | Density | β-glu | N-acet | β-xyl | β-cello | pho | tyr | arg |
|---|---|---|---|---|---|---|---|---|---|
| 0–20 cm | Control | LD | 500.01 a (29.67) | 240.78 a (20.45) | 116.95 a (10.49) | 85.76 a (16.29) | 1983.20 a (167.84) | 185.00 b (27.82) | 258.69 b (28.46) |
| MD | 859.12 a (103.54) | 216.54 a (36.29) | 128.66 a (11.13) | 105.40 a (15.35) | 1570.11 a (421.56) | 350.67 a (13.8) | 595.59 a (5.04) | ||
| HD | 622.26 a (30.63) | 304.82 a (21.31) | 134.91 a (6.25) | 95.47 a (14.03) | 1861.63 a (150.35) | 211.06 b (35.93) | 340.14 b (43.30) | ||
| Cellulose | LD | 3710.13 b (170.85) * | 2759.40 a (218.98) * | 791.96 a (36.43) * | 715.70 a (166.82) * | 18,159.09 a (521.17) * | 243.48 b (12.38) | 427.36 b (39.77) * | |
| MD | 4543.38 a (486.78) * | 1389.09 c (153.62) * | 724.97 a (58.73) * | 446.70 b (68.65) * | 10,902.76 b (885.14) * | 390.96 a (43.05) | 730.36 a (32.98) | ||
| HD | 3373.42 b (193.90) * | 2192.40 b (123.31) * | 680.28 a (57.45) * | 443.14 b (65.57) * | 11,712.99 b (1748.08) * | 306.82 b (27.64) * | 516.48 b (24.72) | ||
| 20–40 cm | Control | LD | 1807.84 b (191.85) | 1123.05 a (143.49) | 672.21 a (32.80) | 327.02 a (19.60) | 12,301.62 a (368.78) | 146.48 a (13.97) | 242.27 a (46.96) |
| MD | 2792.59 a (427.59) | 1362.95 a (60.06) | 824.95 a (68.19) | 387.71 a (44.86) | 12,557.36 a (273.12) | 203.45 a (20.54) | 295.42 a (32.47) | ||
| HD | 1650.74 b (197.79) | 1193.27 a (132.94) | 663.68 a (121.98) | 402.52 a (93.86) | 13,397.28 a (648.90) | 142.85 a (60.85) | 183.01 a (71.83) | ||
| Cellulose | LD | 2075.58 a (343.62) | 1475.58 b (87.90) * | 636.04 b (15.64) | 158.18 c (45.61) * | 14,279.66 b (541.38) * | 167.46 a (42.64) | 410.46 b (21.73) | |
| MD | 2027.89 a (116.52) * | 1719.33 b (134.22) * | 674.19 b (22.14) | 387.48 b (16.55) | 14,303.86 b (298.03) | 153.76 a (18.98) | 256.73 c (61.66) | ||
| HD | 2229.84 a (116.12) * | 2085.64 a (160.47) * | 2049.15 a (64.08) * | 939.41 a (47.07) * | 25,002.57 a (773.87) * | 176.89 a (5.07) | 1460.92 a (85.52) * | ||
| 40–60 cm | Control | LD | 1639.56 a (130.29) | 1955.34 a (114.28) | 687.46 b (163.97) | 560.03 a (63.29) | 13,030.01 a (1537.30) | 45.10 b (24.8) | 900.74 a (58.51) |
| MD | 1490.02 a (225.11) | 1649.26 a (55.68) | 467.42 b (18.13) | 367.09 b (33.36) | 11,220.70 a (138.31) | 98.86 a (12.12) | 941.19 a (62.18) | ||
| HD | 2298.75 a (154.64) | 1017.12 b (313.84) | 1051.88 a (95.55) | 690.48 a (51.53) | 13,987.43 a (926.54) | 118.59 a (8.06) | 948.92 a (53.64) | ||
| Cellulose | LD | 1014.58 b (308.63) | 693.71 b (269.16) * | 730.09 a (365.97) * | 270.27 b (70.27) * | 7295.34 c (3647.68) * | 67.78 a (11.42) | 576.35 a (182.81) * | |
| MD | 1653.36 a (107.32) | 1528.03 a (62.80) | 817.26 a (90.73) * | 269.87 b (48.03) | 16,227.29 a (1004.07) * | 124.14 a (10.82) | 227.89 b (38.91) * | ||
| HD | 1826.48 a (101.26) | 1376.07 a (43.9) | 717.67 a (19.00) * | 514.15 a (72.46) | 11,286.67 b (579.34) * | 106.03 a (23.08) | 495.64 a (51.44) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Sun, H.; Chen, J.; Gao, G.; Li, R.; Li, J.; Li, Y.; Sun, X.; Zhang, Y. Effect of Stand Density on Soil Organic Carbon Storage and Extracellular Enzymes Activity of Larch Plantation in Northeast China. Forests 2023, 14, 1412. https://doi.org/10.3390/f14071412
Sun X, Sun H, Chen J, Gao G, Li R, Li J, Li Y, Sun X, Zhang Y. Effect of Stand Density on Soil Organic Carbon Storage and Extracellular Enzymes Activity of Larch Plantation in Northeast China. Forests. 2023; 14(7):1412. https://doi.org/10.3390/f14071412
Chicago/Turabian StyleSun, Xudong, Hailong Sun, Juan Chen, Guoqiang Gao, Rui Li, Jinfang Li, Yang Li, Xiaoyang Sun, and Yandong Zhang. 2023. "Effect of Stand Density on Soil Organic Carbon Storage and Extracellular Enzymes Activity of Larch Plantation in Northeast China" Forests 14, no. 7: 1412. https://doi.org/10.3390/f14071412