
Impacts of Forest Management on the Biodiversity and Sustainability of Carya dabieshanensis Forests
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
2.3. Data Analysis
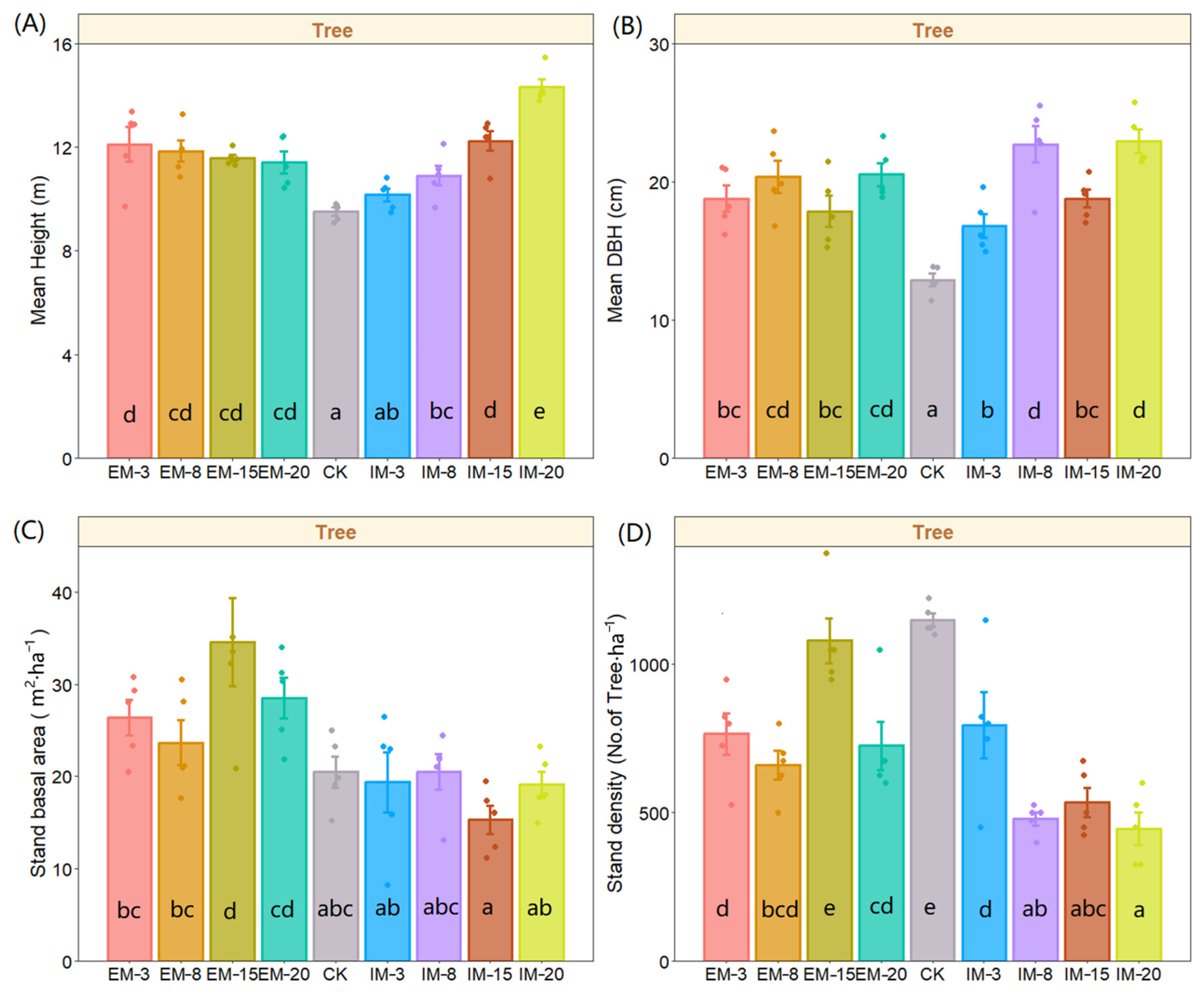
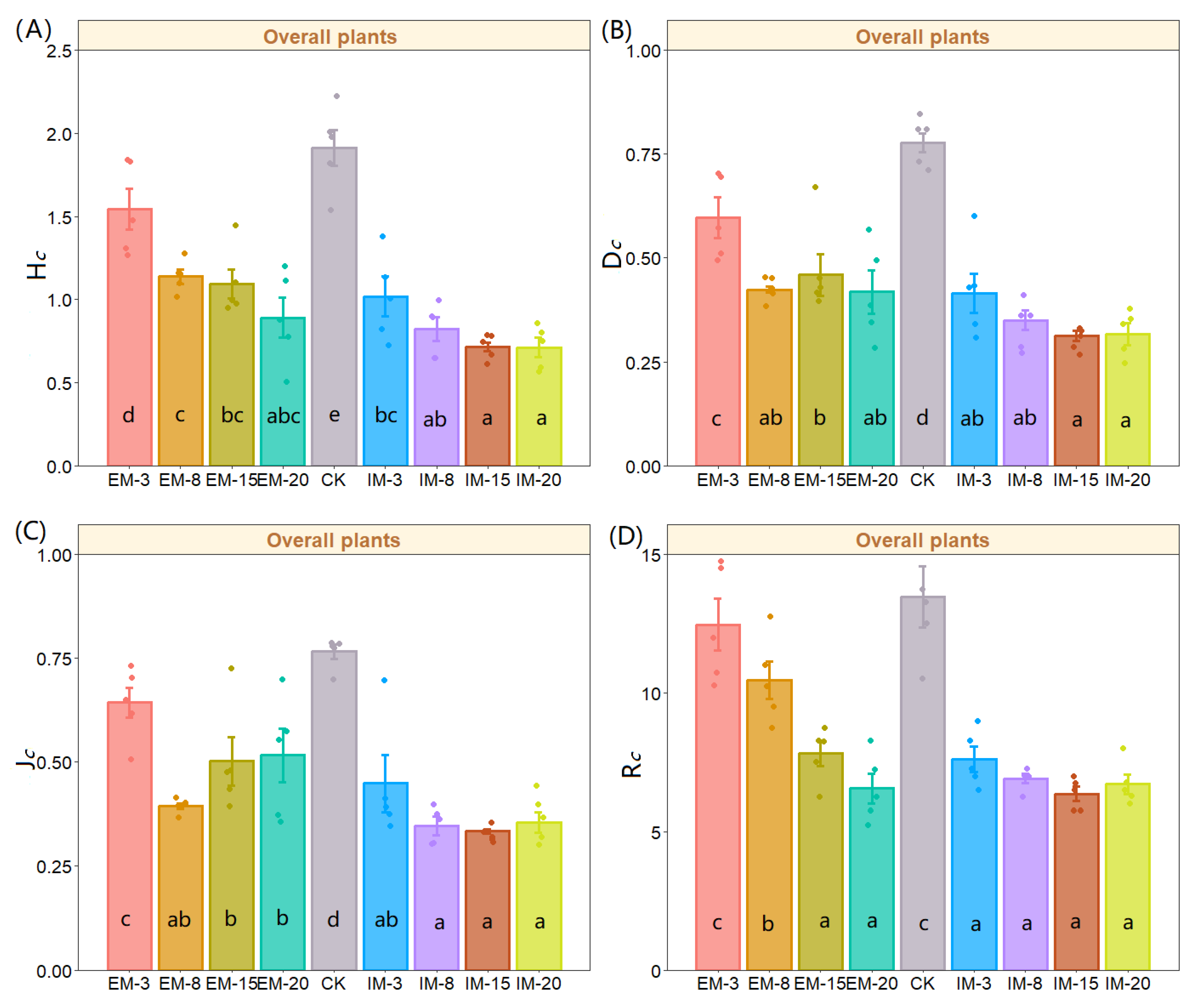
3. Results
3.1. Effects on Stand Characteristics
3.2. Effects on the α Diversity Index of Species for Each Forest Layer
3.3. Effects on Species β Diversity at All Levels
3.4. Effects on Community Stability
4. Discussion
4.1. Vegetation Community Species Diversity
4.2. Vegetation Community Stability
5. Conclusions
- Long-term forest management led to a continuous decline in species diversity in the plant communities of natural secondary forests in Dashan Mountain, which was expedited compared with intensive management. Community species diversity at both management intensities generally stabilized after 15 years, which did not align with our first hypothesis.
- Since there was no evidence that community species diversity translated to community stability, and intensive management did not lead to lower community stability under the same management durations, we rejected Hypotheses 2 and 3.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vellend, M.; Baeten, L.; Becker-Scarpitta, A.; Boucher-Lalonde, V.; McCune, J.L.; Messier, J.; Myers-Smith, I.H.; Sax, D.F. Plant Biodiversity Change Across Scales During the Anthropocene. Annu. Rev. Plant Biol. 2017, 68, 563–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.L.; Zhou, C.J.; Zhou, X.N.; Hu, X.S.; Gan, J.B. Variability after 15 Years of Vegetation Recovery in Natural Secondary Forest with Timber Harvesting at Different Intensities in Southeastern China: Community Diversity and Stability. Forests 2018, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Taylor, A.R.; Reich, P.B.; Hisano, M.; Chen, H.Y.H.; Chang, S.X. Tree diversity increases decadal forest soil carbon and nitrogen accrual. Nature 2023, 618, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Kermavnar, J.; Eler, K.; Marinsek, A.; Kutnar, L. Post-harvest forest herb layer demography: General patterns are driven by pre-disturbance conditions. For. Ecol. Manag. 2021, 491, 13. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.H.; Chen, X.; Huang, Z. Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration. Nat. Commun. 2019, 10, 1332. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, R.L.; Chen, H.Y.H.; Bartels, S.F.; Palik, B.J.; Prepas, E.E. Compositional stability of boreal understorey vegetation after overstorey harvesting across a riparian ecotone. J. Veg. Sci. 2015, 26, 733–741. [Google Scholar] [CrossRef]
- Vakili, M.; Shakeri, Z.; Motahari, S.; Farahani, M.; Robbins, Z.J.; Scheller, R.M. Resistance and Resilience of Hyrcanian Mixed Forests Under Natural and Anthropogenic Disturbances. Front. For. Glob. Change 2021, 4, 14. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Gou, M.; Chen, L.; Lei, P.; Xiao, W.; Deng, X.; Zeng, L.; Li, J.; Zhang, T.; et al. Stability in subtropical forests: The role of tree species diversity, stand structure, environmental and socio-economic conditions. Glob. Ecol. Biogeogr. 2020, 30, 500–513. [Google Scholar] [CrossRef]
- Forrester, D.I.; Rodenfels, P.; Haase, J.; Hardtle, W.; Leppert, K.N.; Niklaus, P.A.; von Oheimb, G.; Scherer-Lorenzen, M.; Bauhus, J. Tree-species interactions increase light absorption and growth in Chinese subtropical mixed-species plantations. Oecologia 2019, 191, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Mazzochini, G.G.; Fonseca, C.R.; Costa, G.C.; Santos, R.M.; Oliveira, A.T.; Ganade, G. Plant phylogenetic diversity stabilizes large-scale ecosystem productivity. Glob. Ecol. Biogeogr. 2019, 28, 1430–1439. [Google Scholar] [CrossRef]
- Esquivel-Muelbert, A.; Baker, T.R.; Dexter, K.G.; Lewis, S.L.; Brienen, R.J.W.; Feldpausch, T.R.; Lloyd, J.; Monteagudo-Mendoza, A.; Arroyo, L.; Alvarez-Davila, E.; et al. Compositional response of Amazon forests to climate change. Glob. Change Biol. 2019, 25, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, J.; Fibich, P.; Altman, J.; Leps, J.; Uemura, S.; Takahashi, K.; Hara, T. Determinants of ecosystem stability in a diverse temperate forest. Oikos 2020, 129, 1692–1703. [Google Scholar] [CrossRef]
- Jiang, L.M.; Sattar, K.; Lu, G.H.; Hu, D.; Zhang, J.; Yang, X.D. Different contributions of plant diversity and soil properties to the community stability in the arid desert ecosystem. Front. Plant Sci. 2022, 13, 969852. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Gao, Y.; Mei, Y.; Liu, J.; Kalkhajeh, Y.K.; Hu, H.; Huang, J. Effects of continuous straw returning on bacterial community structure and enzyme activities in rape-rice soil aggregates. Sci. Rep. 2023, 13, 2357. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Yan, E.R. The mediation roles of intraspecific and interspecific functional trait diversity for linking the response of aboveground biomass to species richness across forest strata in a subtropical forest. Ecol. Indic. 2018, 85, 493–501. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 2016, 91, 760–781. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zheng, G.; Chen, H.Y.H. Understory diversity are driven by resource availability rather than resource heterogeneity in subtropical forests. For. Ecol. Manag. 2022, 503, 9. [Google Scholar] [CrossRef]
- Tsai, H.-C.; Chiang, J.-M.; McEwan, R.W.; Lin, T.-C. Decadal effects of thinning on understory light environments and plant community structure in a subtropical forest. Ecosphere 2018, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.E.; Jose, S. The role of herbicide in savanna restoration: Effects of shrub reduction treatments on the understory and overstory of a longleaf pine flatwoods. For. Ecol. Manag. 2009, 257, 978–986. [Google Scholar] [CrossRef]
- Nobilly, F.; Atikah, S.N.; Yahya, M.S.; Jusoh, S.; Cun, G.S.; Norhisham, A.R.; Tohiran, K.A.; Zulkifli, R.; Azhar, B. Rotational cattle grazing improves understory vegetation biodiversity and structural complexity in oil palm plantations. Weed Biol. Manage. 2022, 22, 13–26. [Google Scholar] [CrossRef]
- Hou, X.Y.; Liu, S.L.; Cheng, F.Y.; Su, X.K.; Dong, S.K.; Zhao, S.; Liu, G.H. Variability of environmental factors and the effects on vegetation diversity with different restoration years in a large open-pit phosphorite mine. Ecol. Eng. 2019, 127, 245–253. [Google Scholar] [CrossRef]
- Koncz, P.; Vadasz-Besnyoi, V.; Csatho, A.I.; Nagy, J.; Szerdahelyi, T.; Toth, Z.; Pinter, K.; Foti, S.; Papp, M.; Balogh, J.; et al. Carbon uptake changed but vegetation composition remained stable during transition from grazing to mowing grassland management. Agric. Ecosyst. Environ. 2020, 304, 11. [Google Scholar] [CrossRef]
- Braun, A.C.; Koch, B. Estimating impacts of plantation forestry on plant biodiversity in southern Chile-a spatially explicit modelling approach. Environ. Monit. Assess. 2016, 188, 564. [Google Scholar] [CrossRef]
- Medina, O.; Manian, V.; Chinea, J.D. Biodiversity assessment using hierarchical agglomerative clustering and spectral unmixing over hyperspectral images. Sensors 2013, 13, 13949–13959. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, H.R.; Torezan, J.M. Evaluating the ecological integrity of Atlantic forest remnants by using rapid ecological assessment. Environ. Monit. Assess. 2013, 185, 4373–4382. [Google Scholar] [CrossRef]
- Ngo Bieng, M.A.; Delgado-Rodriguez, D.; Vilchez-Mendoza, S.; Lopez-Sampson, A.; Garcia, E.; Sepulveda, N.; Somarriba, E. Tree diversity in a tropical agricultural-forest mosaic landscape in Honduras. Sci. Rep. 2022, 12, 18544. [Google Scholar] [CrossRef] [PubMed]
- Godron, M. Some aspects of heterogeneity in grasslands of Cantal. Stat. Ecol. 1972, 3, 397–415. [Google Scholar]
- Dovciak, M.; Halpern, C.B. Positive diversity-stability relationships in forest herb populations during four decades of community assembly. Ecol. Lett. 2010, 13, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, Y.; Shen, D.; Xia, G.; Huang, Y.; Huang, Y.; Chang, J.; Huang, J.; Wang, Z. Development of SSR Markers in Hickory (Carya cathayensis Sarg.) and Their Transferability to Other Species of Carya. Curr. Genom. 2014, 15, 357–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Fu, S.; Ma, X.; Ma, X.; Ren, X.; Tian, X.; Tong, Y.; Yuan, F.; Liu, H. Long-term intensive management reduced the soil quality of a Carya dabieshanensis forest. Sci. Rep. 2023, 13, 5058. [Google Scholar] [CrossRef]
- Ni, H.J.; Su, W.H.; Fan, S.H.; Chu, H.Y. Effects of intensive management practices on rhizosphere soil properties, root growth, and nutrient uptake in Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2021, 493, 11. [Google Scholar] [CrossRef]
- Li, Y.F.; Zhang, J.J.; Chang, S.X.; Jiang, P.K.; Zhou, G.M.; Shen, Z.M.; Wu, J.S.; Lin, L.; Wang, Z.S.; Shen, M.C. Converting native shrub forests to Chinese chestnut plantations and subsequent intensive management affected soil C and N pools. For. Ecol. Manag. 2014, 312, 161–169. [Google Scholar] [CrossRef]
- Das, D.; Dwivedi, B.S.; Datta, S.P.; Datta, S.C.; Meena, M.C.; Dwivedi, A.K.; Singh, M.; Chakraborty, D.; Jaggi, S. Long-term differences in nutrient management under intensive cultivation alter potassium supplying ability of soils. Geoderma 2021, 393, 16. [Google Scholar] [CrossRef]
- Tie-Liang, S.; Feng, Z.; Jin-Tun, Z. Plant diversity of forest community in zhuweigou of lishan mountain nature reserve. Chin. J. Plant Ecol. 2002, 26, 46–51. (In Chinese) [Google Scholar]
- Wu, Z.-l.; Zhou, X.-n.; Zheng, L.-f.; Hu, X.-s.; Zhou, C.-j. Species diversity and stability of natural secondary communities with different cutting intensities after ten years. J. For. Res. 2011, 22, 205–208. [Google Scholar] [CrossRef]
- Norden, N.; Chazdon, R.L.; Chao, A.; Jiang, Y.H.; Vilchez-Alvarado, B. Resilience of tropical rain forests: Tree community reassembly in secondary forests. Ecol. Lett. 2009, 12, 385–394. [Google Scholar] [CrossRef]
- Thorn, S.; Bassler, C.; Svoboda, M.; Muller, J. Effects of natural disturbances and salvage logging on biodiversity–Lessons from the Bohemian Forest. For. Ecol. Manag. 2017, 388, 113–119. [Google Scholar] [CrossRef]
- Walter, C.A.; Adams, M.B.; Gilliam, F.S.; Peterjohn, W.T. Non-random species loss in a forest herbaceous layer following nitrogen addition. Ecology 2017, 98, 2322–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, E.; Newton, M.; Bailey, J.D. Understory vegetation dynamics 15 years post-thinning in 50-year-old Douglas-fir and Douglas-fir/western hemlock stands in western Oregon, USA. For. Ecol. Manag. 2017, 384, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Fan, S.; Guan, F.; Xia, W.; Wang, S.; Xiao, X. Strip Clearcutting Drives Vegetation Diversity and Composition in the Moso Bamboo Forests. For. Sci. 2022, 68, 27–36. [Google Scholar] [CrossRef]
- Li, B.; Zeng, T.; Ran, J.H.; Yue, B.S.; Zhang, M.; Shang, T.; Zhu, D.H. Characteristics of the early secondary succession after landslides in a broad-leaved deciduous forest in the south Minshan Mountains. For. Ecol. Manag. 2017, 405, 238–245. [Google Scholar] [CrossRef]
- Hrivnak, R.; Bosela, M.; Slezak, M.; Lukac, M.; Svitkova, I.; Gizela, J.; Hegedusova, K.; Hrivnak, M.; Kliment, J.; Knopp, V.; et al. Competition for soil resources forces a trade-off between enhancing tree productivity and understorey species richness in managed beech forests. Sci. Total Environ. 2022, 849, 157825. [Google Scholar] [CrossRef]
- Williams, L.J.; Butler, E.E.; Cavender-Bares, J.; Stefanski, A.; Rice, K.E.; Messier, C.; Paquette, A.; Reich, P.B. Enhanced light interception and light use efficiency explain overyielding in young tree communities. Ecol. Lett. 2021, 24, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; du Toit, B.; Seifert, T. Diversity-biomass relationship across forest layers: Implications for niche complementarity and selection effects. Oecologia 2018, 187, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, R.; Ueno, M.; Masaki, T. Initial effects of thinning and concomitant disturbance on the understory woody community in Japanese cedar plantation. J. For. Res. 2018, 23, 120–128. [Google Scholar] [CrossRef]
- Monarrez-Gonzalez, J.C.; Gonzalez-Elizondo, M.S.; Marquez-Linares, M.A.; Gutierrez-Yurrita, P.J.; Perez-Verdin, G. Effect of forest management on tree diversity in temperate ecosystem forests in northern Mexico. PLoS ONE 2020, 15, e0233292. [Google Scholar] [CrossRef]
- Zhang, C.; Xie, G.; Fan, S.; Zhen, L. Variation in vegetation structure and soil properties, and the relation between understory plants and environmental variables under different Phyllostachys pubescens forests in southeastern China. Environ. Manag. 2010, 45, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Sebald, J.; Thrippleton, T.; Rammer, W.; Bugmann, H.; Seidl, R. Mixing tree species at different spatial scales: The effect of alpha, beta and gamma diversity on disturbance impacts under climate change. J. Appl. Ecol. 2021, 58, 1749–1763. [Google Scholar] [CrossRef]
- Gong, C.; Tan, Q.; Liu, G.; Xu, M. Impacts of tree mixtures on understory plant diversity in China. For. Ecol. Manag. 2021, 498, 7. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; de Mazancourt, C.; Scherer-Lorenzen, M.; Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 2014, 17, 1526–1535. [Google Scholar] [CrossRef]
- Schnabel, F.; Schwarz, J.A.; Danescu, A.; Fichtner, A.; Nock, C.A.; Bauhus, J.; Potvin, C. Drivers of productivity and its temporal stability in a tropical tree diversity experiment. Glob. Change Biol. 2019, 25, 4257–4272. [Google Scholar] [CrossRef] [PubMed]
- May, R.M. Stability and Complexity in Model Ecosystems. IEEE Trans. Syst. Man Cybern. 1978, 8, 779. [Google Scholar] [CrossRef]
- Jansen, V.A.A.; Kokkoris, G.D. Complexity and stability revisited. Ecol. Lett. 2003, 6, 498–502. [Google Scholar] [CrossRef]
- Sasaki, T.; Lauenroth, W.K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia 2011, 166, 761–768. [Google Scholar] [CrossRef]
- Pillar, V.D.; Blanco, C.C.; Muller, S.C.; Sosinski, E.E.; Joner, F.; Duarte, L.D.S. Functional redundancy and stability in plant communities. J. Veg. Sci. 2013, 24, 963–974. [Google Scholar] [CrossRef]
- Kang, S.; Ma, W.; Li, F.Y.; Zhang, Q.; Niu, J.; Ding, Y.; Han, F.; Sun, X. Functional Redundancy Instead of Species Redundancy Determines Community Stability in a Typical Steppe of Inner Mongolia. PLoS ONE 2015, 10, e0145605. [Google Scholar] [CrossRef]
- Yu, Q.S.; Rao, X.Q.; Chu, C.J.; Liu, S.P.; Lin, Y.B.; Sun, D.; Tan, X.P.; Hanif, A.; Shen, W.J. Species dominance rather than species asynchrony determines the temporal stability of productivity in four subtropical forests along 30 years of restoration. For. Ecol. Manag. 2020, 457, 11. [Google Scholar] [CrossRef]
- Allesina, S.; Tang, S. Stability criteria for complex ecosystems. Nature 2012, 483, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.W.; Wang, X.P.; Fan, D.Y. Ecosystem functioning and stability are mainly driven by stand structural attributes and biodiversity, respectively, in a tropical forest in Southwestern China. For. Ecol. Manag. 2021, 481, 10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| df | Sum of Squares | Mean Squares | F | p-Value | ||
|---|---|---|---|---|---|---|
| Mean tree height | Between Groups | 8 | 75.262 | 9.408 | 13.287 | *** |
| (m) | Within Groups | 36 | 25.489 | 0.708 | ||
| Mean tree DBH | Between Groups | 8 | 384.053 | 48.007 | 10.596 | *** |
| (cm) | Within Groups | 36 | 163.098 | 4.531 | ||
| Stand basal area | Between Groups | 8 | 1380.824 | 172.603 | 5.211 | *** |
| (m2 · ha−1) | Within Groups | 36 | 1192.493 | 33.125 | ||
| Stand density | Between Groups | 8 | 2,452,777.800 | 306,597.200 | 14.256 | *** |
| (trees · ha−1) | Within Groups | 36 | 774,250.000 | 21,506.940 |
| df | Sum of Squares | Mean Squares | F | p-Value | |||
|---|---|---|---|---|---|---|---|
| Overall plants | HC | Between Groups | 8 | 6.408 | 0.801 | 19.104 | *** |
| Within Groups | 36 | 1.509 | 0.042 | ||||
| DC | Between Groups | 8 | 0.888 | 0.111 | 16.850 | *** | |
| Within Groups | 36 | 0.237 | 0.007 | ||||
| JC | Between Groups | 8 | 0.861 | 0.108 | 12.945 | *** | |
| Within Groups | 36 | 0.299 | 0.008 | ||||
| RC | Between Groups | 8 | 295.461 | 36.933 | 19.262 | *** | |
| Within Groups | 36 | 69.025 | 1.917 |
| df | Sum of Squares | Mean Squares | F | p-Value | |||
|---|---|---|---|---|---|---|---|
| Tree | H | Between Groups | 8 | 12.643 | 1.580 | 24.906 | *** |
| Within Groups | 36 | 2.284 | 0.063 | ||||
| D | Between Groups | 8 | 2.395 | 0.299 | 19.85 | *** | |
| Within Groups | 36 | 0.543 | 0.015 | ||||
| J | Between Groups | 8 | 2.900 | 0.363 | 16.576 | *** | |
| Within Groups | 36 | 0.787 | 0.022 | ||||
| R | Between Groups | 8 | 300.178 | 37.522 | 21.647 | *** | |
| Within Groups | 36 | 62.400 | 1.733 | ||||
| Shrub | H | Between Groups | 8 | 8.301 | 1.038 | 8.563 | *** |
| Within Groups | 36 | 4.362 | 0.121 | ||||
| D | Between Groups | 8 | 0.379 | 0.047 | 3.991 | ** | |
| Within Groups | 36 | 0.427 | 0.012 | ||||
| J | Between Groups | 8 | 0.144 | 0.018 | 2.722 | * | |
| Within Groups | 36 | 0.238 | 0.007 | ||||
| R | Between Groups | 8 | 792.400 | 99.05 | 15.477 | *** | |
| Within Groups | 36 | 230.400 | 6.400 | ||||
| Herb | H | Between Groups | 8 | 4.291 | 0.536 | 4.488 | ** |
| Within Groups | 36 | 4.302 | 0.120 | ||||
| D | Between Groups | 8 | 0.343 | 0.043 | 3.405 | ** | |
| Within Groups | 36 | 0.453 | 0.013 | ||||
| J | Between Groups | 8 | 0.277 | 0.035 | 2.782 | * | |
| Within Groups | 36 | 0.448 | 0.012 | ||||
| R | Between Groups | 8 | 470.800 | 58.850 | 9.543 | *** | |
| Within Groups | 36 | 222.000 | 6.167 |
| df | Sum of Squares | R2 | F | p-Value | ||
|---|---|---|---|---|---|---|
| Tree | Between Groups | 8 | 1.121 | 0.514 | 4.756 | *** |
| Within Groups | 36 | 1.060 | 0.486 | |||
| Shrub | Between Groups | 8 | 7.182 | 0.481 | 4.178 | *** |
| Within Groups | 36 | 7.735 | 0.519 | |||
| Herb | Between Groups | 8 | 6.990 | 0.589 | 6.460 | *** |
| Within Groups | 36 | 11.860 | 1.000 |
| Management Intensity and Duration | Equations | R2 | (x, y) | d |
|---|---|---|---|---|
| CK | y = −0.0144 × 2 + 2.17x + 20 | 0.961 | (29.08, 70.93) | 12.841 |
| EM-3 | y = −0.0129 × 2 + 2.07x + 16.4 | 0.985 | (31.36, 68.64) | 16.065 |
| EM-8 | y = −0.0117 × 2 + 2x + 14 | 0.992 | (32.88, 67.12) | 18.215 |
| EM-15 | y = −0.0127 × 2 + 2.08x + 14.3 | 0.988 | (32.06, 67.94) | 17.055 |
| EM-20 | y = −0.0148 × 2 + 2.25x + 15.7 | 0.986 | (29.29, 70.71) | 13.138 |
| IM-3 | y = −0.00992 × 2 + 1.75x + 21.4 | 0.99 | (32.36, 67.64) | 17.480 |
| IM-8 | y = −0.012 × 2 + 1.91x + 22.9 | 0.971 | (30.27, 69.73) | 14.524 |
| IM-15 | y = −0.0116 × 2 + 2.93x + 18.7 | 0.99 | (31.73, 68.27) | 16.589 |
| IM-20 | y = −0.0106 × 2 + 1.73x + 29 | 0.978 | (29.35, 70.65) | 13.223 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Fu, S.; Tong, Y.; Ma, X.; Yuan, F.; Ma, Y.; Feng, C.; Liu, H. Impacts of Forest Management on the Biodiversity and Sustainability of Carya dabieshanensis Forests. Forests 2023, 14, 1331. https://doi.org/10.3390/f14071331
Huang C, Fu S, Tong Y, Ma X, Yuan F, Ma Y, Feng C, Liu H. Impacts of Forest Management on the Biodiversity and Sustainability of Carya dabieshanensis Forests. Forests. 2023; 14(7):1331. https://doi.org/10.3390/f14071331
Chicago/Turabian StyleHuang, Cheng, Songling Fu, Yinhao Tong, Xiaomin Ma, Feiyang Yuan, Yuhua Ma, Chun Feng, and Hua Liu. 2023. "Impacts of Forest Management on the Biodiversity and Sustainability of Carya dabieshanensis Forests" Forests 14, no. 7: 1331. https://doi.org/10.3390/f14071331