1. Introduction
A cooperative genetic improvement program for coastal Douglas-fir (
Pseudotsuga menziesii var.
menziesii [Mirb.] Franco) in the U.S. Pacific Northwest (PNW) started in 1954, with progeny tests now in a third cycle of testing and selection [
1,
2,
3,
4]. Most progeny tests are designed with the primary objective of identifying families that exhibit superior volume growth without sacrificing fitness to their targeted biophysical environment. Traits used for selecting superior trees are, therefore, dimensions that determine tree stem volume growth, specifically total height (HT) and diameter at breast height (DBH). A typical progeny test contains trees that are completely interspersed by family origin. In contrast, operational plantations containing genetically improved stock originate from seed lots containing a set of only families with better growth performance in progeny tests [
5,
6,
7]. Selections from progeny tests are often performed at a young age (one-fourth to one-half of the rotation age) using narrow spacing, so crown closure and inter-tree competition are typically prevalent in most stands. It has been repeatedly demonstrated that selection in forest tree improvement programs at around 1/4 to 1/3 of rotation age is economically optimal in terms of net present value added [
8]. Most selections are, therefore, indirect and possibly subject to some revision because (1) progeny tests were established to measure the relative growth performance of individual, relatively open-grown trees; (2) the relative performance of young open-grown trees may change as they approach crown closure and the onset of increasingly intense intertree competition; and (3) genetic gain can be overestimated if the measurement takes place after the severe inter-tree competition has taken place [
9,
10]. However, realized genetic gain trials established for several conifer species confirm that breeding and testing programs deliver significant gains in large-plot trials where elite genotypes are grown together in competition [
5,
6,
7].
Donald [
11] introduced the crop ideotype concept and extended the concept to plant breeding and conceptual models of the ideal plant for maximizing yield per unit area [
12,
13,
14]. The crop ideotype concept focuses on yield enhancement attributable to multiple phenotypic attributes of a plant that are expected to produce higher biomass and economic yield in a fixed area and under a fixed biophysical environment. Dickmann [
15] and Cannell [
16] further expanded the ideotype concept to forests by adopting it as a strategy for the genetic improvement of trees [
17,
18,
19]. The concept of crop ideotype distinguishes three different strategies. The isolation ideotype grows well in an open space without competition and tends to have a wide and long crown but does not grow well under intensive competition. The competition ideotype tends to exploit site resources rapidly and grow large at the expense of neighbors, in part by developing relatively wide crowns. Resource-use efficiency manifest as production per unit occupied area is therefore low for competition ideotypes. In contrast, the crop ideotype exploits resources more efficiently by occupying less growing space and using lesser amounts of most or all resources to achieve the same level of growth as the competition ideotype. The ideal improved stands will theoretically include only families conforming most closely to the crop ideotype by having long but narrow live crowns, more trees per unit area, more leaf area per unit area, higher tree-level and stand-level leaf area index, and consequently greater yield per unit area, especially under intensive silviculture [
13,
16].
In progeny tests, family selection typically occurs before crown closure; however, some degree of intertree competition has typically commenced as crown closure approaches. The crop ideotype concept might supplement selection criteria based on growth rate because (1) even mild to moderate intertree competition may cause underestimation of tree- and stand-level growth potential of isolation ideotype families; (2) individual trees from competition ideotype families may exhibit superior growth of individual trees at the expense of stand-level growth; (3) competition ideotype families may grow better before crown closure, so earlier selection may cause overestimation of their stand-level growth potential; and (4) the relatively slow growth of crop ideotypes before crown closure may cause underestimation of stand-level performance in crop ideotype plantations due to better resource use efficiency and reduced intertree competition (e.g., progeny tests) [
9,
20]. Because many of the traits for identifying crop ideotypes are more difficult to measure than height and diameter growth, implementing these concepts in operational tree improvement programs could add substantially to testing costs.
Because wood production in forestry involves a 40–70-year rotation in species like Douglas-fir, forecasts are needed to estimate the yield gains from genetic improvement. Forest growth and yield models have accounted for genetic improvements by adjusting site index or applying genetic-gain multipliers in height and diameter growth rate equations [
21,
22,
23]. Identifying physiological or morphological crown attributes consistent with the crown ideotype concept could not only help explain differences in the tree and stand growth performance among families, but their incorporation into mechanistic growth and yield models offers the potential to facilitate simulation of family combinations that have not or cannot be field tested.
Past work on progeny tests in Douglas-fir provides insight into attributes justifying close focus during tree breeding. St. Clair [
24,
25] felled sample trees from an 18-year-old progeny test in the Coast Range of Oregon, measured numerous crown and stem attributes in detail on these felled trees, and then examined the genetic and phenotypic correlations between crown structure and stem growth performance. Although these univariate analyses of whole-tree crown attributes were an important first step in identifying traits potentially controlling differential family performance, ecophysiological mechanisms functionally integrate many morphological attributes in complex interactions. Likewise, because crown attributes are intercorrelated, and height and diameter growth are intercorrelated, multivariate analyses offer an alternative and complementary way to identify, explore, and establish general relationships between stem growth and phenotypic tree and crown structure. A multivariate approach acknowledges the complexity among measurable variables without requiring the much deeper level of understanding required to represent functional relationships explicitly in a mechanistic model. In short, morphological crown ideotypes of practical appeal must be identifiable from key sets of multiple attributes that are measurable and relevant to known ecophysiological processes.
The analyses presented here represented continued research on the progeny tests first explored by St. Clair [
24,
25] to unravel interactions between morphological crown attributes and growth performance, to tentatively infer key mechanisms explaining the differential growth performance of families, and ultimately to incorporate these mechanisms into growth models. Unlike the univariate analyses conducted by St. Clair [
24,
25], our study advances this research using a multivariate approach to analyze the intercorrelations between these crown attributes from each crown third. The specific objectives of the multivariate analyses below were (1) to test whether Douglas-fir families differ concerning morphological crown attributes in one or more thirds of live crown length; (2) to identify crown attributes that are relevant to the crop ideotype concepts; (3) to quantify the strength of relationships between crop ideotype attributes and growth performance of Douglas-fir families; and (4) identify several heritable morphological crown attributes that could be incorporated into individual-tree growth models to represent mechanisms driving genetic improvement and resulting differences in stand-level growth performance of alternative combinations of families.
4. Discussion
Crop ideotype studies in various tree species have most commonly been based on univariate analyses determining how crown attributes affect the attainable stem volume of trees and/or stands, i.e., specifically by the correlation between a potential predictor variable and response variable. Because crown attributes are intercorrelated, and all crown attributes function simultaneously to affect growth performance, characterizing the combined patterns of crown attributes and growth rates among families in a progeny test should provide insight into heritable attributes to target during family selections. Growth performance has been represented by total stem volume at a given age in many studies, but partitioning stem volume growth into height and diameter growth components sharpens insight into biophysical and ecophysiological mechanisms conferring growth advantages. In this study, we had the opportunity to investigate family differences not only in whole-tree crown attributes but also within-tree variation in crown structure. Crown attributes for incorporation into growth models can be identified by determining the variability in crown attributes, their heritability, and the extent to which heritable attributes relate to family differences in growth performance can help identify crown attributes and model structures that can facilitate realistic simulation of growth differences among families identified as superior in progeny tests.
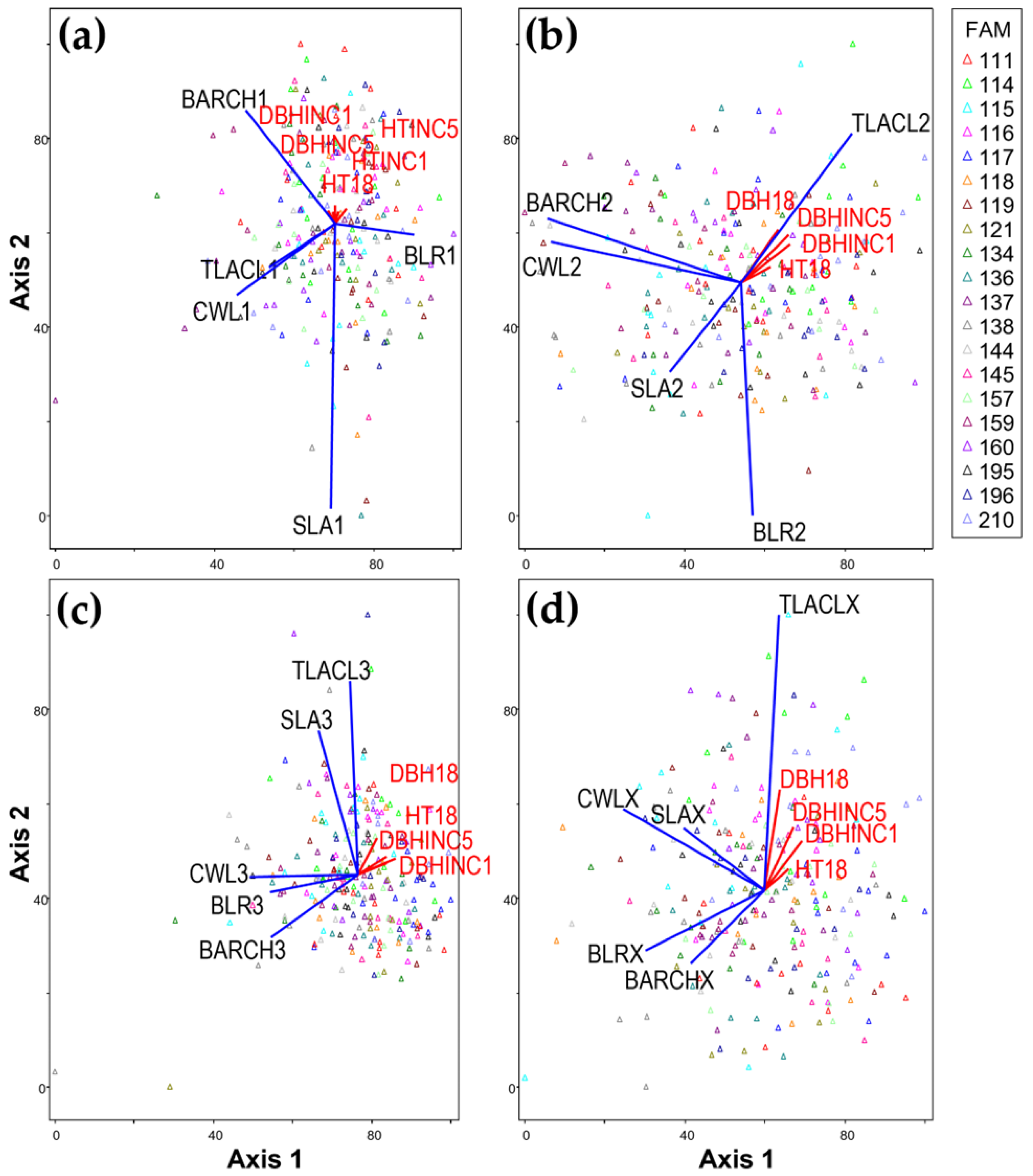
Structural attributes in each third of the live crown length and the entire crown showed significant differences among families, with whole-tree crown variables showing the biggest crown structural differences (see
Table 3). Similar differences in crown structure have been reported at the family level in numerous tree species; however, few studies have tested whether detected differences were attributable to a finer scale difference in specific crown segments. King et al. [
39] reported a significant genetic difference in branch length relative to total height, and Vargas–Hernandez et al. [
40] reported significant differences in the number of branches, branch length, and branch angle among open-pollinated and full-sib families of Douglas-fir seedlings. Total leaf area per crown length (TLACL) appears to be the most important morphological crown attribute differentiating families due to its high heritability (see
Table 4). Gould et al. [
41] detected significant differences among families in crown width relative to DBH, branch length relative to adjacent stem cross-sectional area, and leaf area relative to branch length in high-density Douglas-fir plantations representing pure-family deployment trials. Likewise, a significant amount of genetic variation in crown length, maximum crown diameter, crown volume, foliage biomass, vertical foliage distribution, crown shape, branch longevity, and specific leaf area was found among families of outplanted loblolly pine [
42,
43,
44,
45]. McGarvey et al. [
46] identified potential functional links between crown attributes and growth rates at the family level by confirming that fast-growing families of loblolly and slash pine had higher leaf area and light-saturated net photosynthesis than slow-growing families.
Among crown thirds, the bottom third showed the biggest structural differences and genetic influences, followed by the middle and top crown thirds (see
Table 3 and
Table 5). Greater variation and genetic influence of family origin in the bottom third might indicate the ability of some families to better adapt to cumulative effects of light interception in the upper and middle crown thirds. Kuuluvainen [
47] reported that trees with relatively narrow crowns allow a greater amount of light penetration into the lower crown section. Ishii and Wilson [
48] reported that variations in branch length and branch diameter were relatively small in the upper crown but increased in the lower crown in old-growth Douglas-fir. In the progeny test examined in this analysis, trees with a relatively steep branch angle (low angle from vertical, i.e., closer to vertical) and shorter branch lengths in the middle crown section tended to have higher total leaf area per crown length. However, in the top third of the crown, trees with relatively wider crown width tended to have greater total leaf area per crown length (
Figure 1a,b). The more umbrella shape implied by this latter pattern would be expected for trees and families with relatively slow height growth, short relative height in the stand, and therefore a poorer light environment [
49]. In an analysis of Douglas-fir trees from southwestern Oregon, Hann [
50] verified that dominant trees had a more conic profile, and understory trees were more parabolic in shape.
Branch angles tend to increase from the top to the crown base in Norway spruce [
47] and Douglas-fir [
51]. King et al. [
39] reported that branch angle differs among Douglas-fir families. In this study, trees with relatively steep branch angles (more vertical) tended to have greater total leaf area per crown length, probably due to branch angle effects on effective leaf area [
52] and a resulting increase in average light interception per unit leaf area [
53]. However, the heritabilities for branch angle at a given relative crown height were low in all crown thirds and for the entire crown (
= 0.01 − 0.08;
= 0.01 − 0.53). In contrast, branch angle had moderate heritabilities in Scots pine (
= 0.22) [
54], in Norway spruce (
= 0.44;
= 0.65) [
55], and in slash pine
= 0.33) [
56].
Site index (e.g., [
57]) remains a widely applied measure of site productivity in forest management. This measure relies on the pattern of past height growth rates in dominant and codominant trees on a given site, and height growth equations in growth models are typically constructed to provide height growth rates consistent with site index estimates (e.g., [
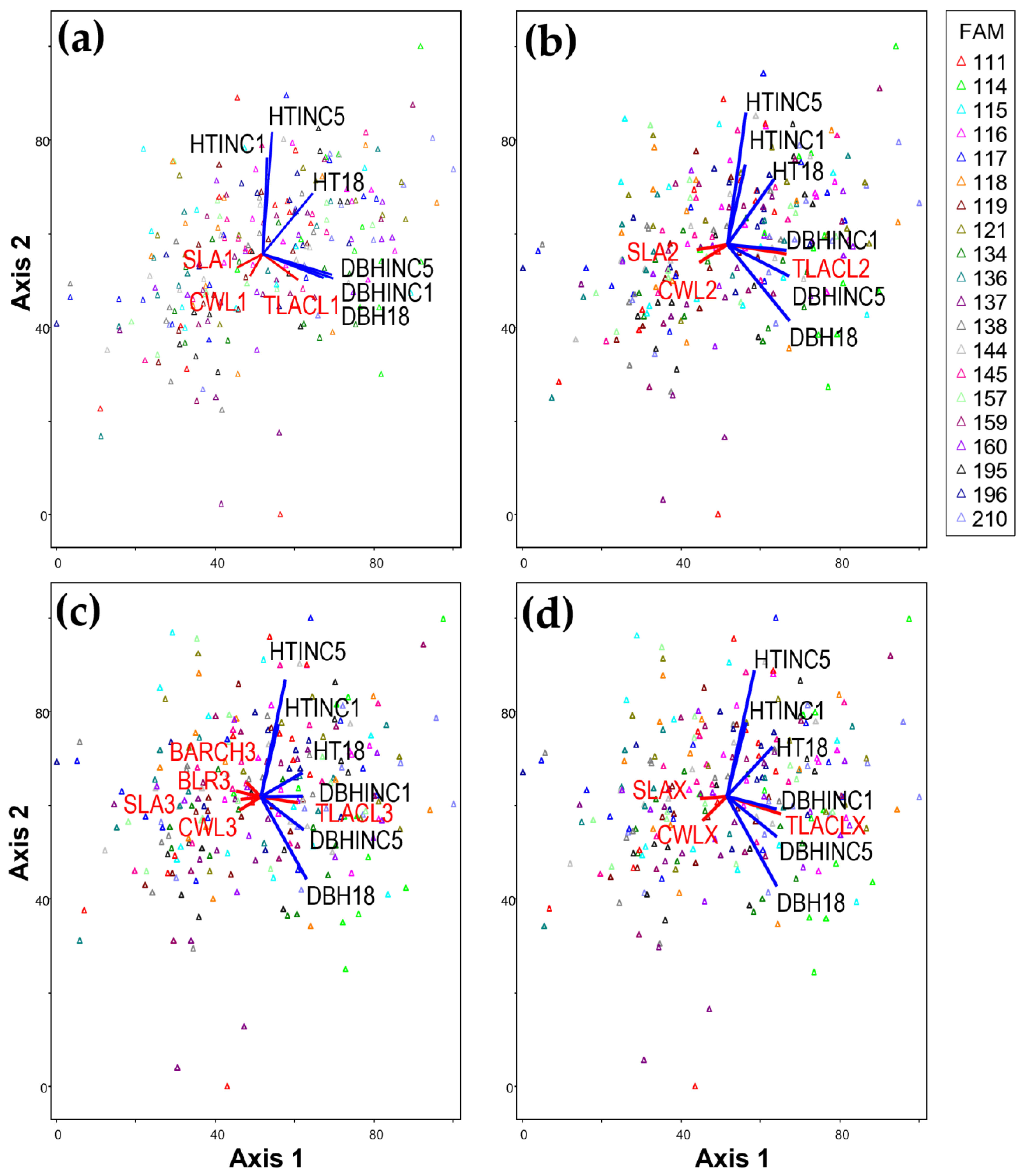
58]). In the top crown section, Axis 2 explained 5.0% of the variation in 1-year and 5-year height increments (HTINC1 and HTINC5, respectively), while in the middle and bottom crown thirds, Axis 2 explained only 2.2% and 2.9%, respectively (
Table 7). In these progeny tests, some variability in height growth was probably initiated and eventually accentuated by family differences in the measured crown attributes. Still, many potentially relevant variables like light availability, apical dominance, relative vigor of primary branches near the top of trees (e.g., [
59]), and many others relevant to ecophysiological processes were not. Unfortunately, the latter class of variables cannot easily be applied in large-scale progeny testing and simulation of stand dynamics and stand growth.
In contrast to height growth, the total leaf area per crown length (TLACL) in all crown sections showed a strong positive correlation with diameter growth (DBHINC1, DBHINC5, and DBH18). Many studies have reported that whole-tree total leaf area or foliage biomass is correlated with sapwood cross-sectional area at breast height [
60,
61,
62] and with DBH [
63,
64]. The former relationship is consistent with the pipe model theory [
65] and the physiological balance between transpiring leaf area and the cross-sectional area of conducting stem tissue. The latter allometric relationship has long been recognized [
66,
67,
68] as the functional relationship between total foliage amount and stem diameter growth [
69] and even cross-sectional stem increment at a given height on the stem (Pressler [
70] as cited in [
69]). Relationships among total leaf area, crown width, crown shape, and stem growth have previously been analyzed in Norway spruce [
47] and Scots pine [
71]. Stemwood production per unit crown projection area was found higher in narrow-crowned trees, presumably partly because light could penetrate deeper into the inner and lower crown. In intensively managed plantations with uniform spacing, trees with a given height and diameter but lower CWL might intercept more solar radiation than those with a higher CWL [
72].
Specific leaf area (SLA; cm
2/g) in conifer species generally has increased from top to bottom of the crown and decreased with foliage age [
63,
73,
74,
75,
76]. In this study, trees with larger DBH18, DBHINC1, DBHINC5, and higher social positions were negatively correlated with SLA in 2-dimensional RDA ordination. Other studies similarly have shown that increasing light exposure to leaves has led to lower SLA (i.e., greater specific leaf weight (g/cm
2)) relative to shaded leaves [
77,
78]. The change in leaf structure causing these patterns in SLA has been related to needle thickness [
79] and the number of palisade layers that can be sustained by differing intensities of incident photosynthetically active radiation (PAR) [
80]. Although some studies have found that tree size in Douglas-fir was only weakly related to tree-average SLA [
66,
76], the influence of relative social position on tree-average SLA is much weaker than the within-crown trend with increasing depth into the crown. Fast-growing families in loblolly pine showed greater SLA than slow-growing families in the fourth growing season [
45]; however, as would be expected, tree-average SLA was lower in families that also had greater foliage mass. The apparent difference from the Douglas-fir progeny test is probably due to the greater shade tolerance of Douglas-fir and perhaps lower genetic variability in the upper limit of SLA under increasing within-crown shade.
The weak negative correlation between DBH-related variables and both branch angle at a given relative crown height (BACRH) and branch length relative to stem diameter below whorl (BLR) was consistent with Campbell’s [
81] findings that branch angle and length were not significantly correlated with stem volume. In contrast to results from the Douglas-fir progeny tests (
Table 4), Birot and Christophe [
82] reported that the branch angle in Douglas-fir had a high heritability (
h2 = 0.49).
In this study, we found that fast-growing trees had higher TLACL. Highly heritable TLACL was strongly correlated with DBH growth and moderately correlated with height growth (
Figure 2 and
Table 4). This relationship may indicate a functional relationship; specifically, higher total leaf area per unit crown length in genetically superior families probably implies great light capture. TLACLX could profitably be incorporated into a diameter growth model to simulate family growth differences. In contrast, for a height growth model, TLACLX may not be sufficient to represent family differences. It is also possible that TLACLX is not appropriate for predicted height growth because the two are confounded; i.e., trees may exhibit lower TLACLX if faster growth causes greater distances between branches. TLACLX, in fact, had a weakly negative correlation with axes representing height increments (
Table 8). Crown attributes indicating less total leaf area, wider branch angles, and longer branch length might be more important for predicting height increment, but they also may result from rather than cause more rapid growth (
Table 8 and
Figure 2). For modeling height growth, therefore, including other crown attributes such as CWLX in addition to TLACLX might better represent family differences, particularly if superior families have larger total leaf area for a given crown length and a lower ratio of crown width to crown length.
5. Conclusions
The multivariate analyses suggested that trees with relatively short branches and steeper branch angles tend to have higher total leaf area per unit crown length. This result may help confirm and quantify the crop ideotype concept by confirming that trees with narrow crown width, longer crown length, relatively shorter branch length, steeper branch angle, and higher total leaf area relative to crown length seem capable of greater volume growth. Theoretically, a given area may be able to carry more of these crop ideotype trees. To the extent that these crown attributes are heritable, the diameter growth rate on the trees from this progeny test conforms to the crop ideotype concept. In contrast, crop ideotype attributes had a less obvious link to the height growth rate. All crown thirds showed significant differences in crown structure among Douglas-fir families. Still, whole-tree crown attributes showed the biggest differences among families, followed successively by bottom, middle, and top crown thirds. Total leaf area per unit crown length (TLACL) could represent at least one of the mechanisms driving genetic gain in Douglas-fir families based on its high heritability and strong positive correlation with growth performance. If families could be indexed by either an empirical construct for TLACL and/or an associated process (e.g., light absorption and/or carbon allocation), they could represent potential gains from genetic improvement in diameter growth models. TLACL is less strongly correlated with height growth rate, so incorporating crown width relative to crown length may give better predictions for genetic effects on height growth.
For further research, a detailed comparison of total leaf area or foliage biomass between fast-growing and slow-growing families is needed. In particular, a comparison of shoot length, number of buds, needle size by age distribution, and needle density and retention would give a better understanding of differences in total leaf area or foliage biomass between fast-growing and slow-growing families. Progeny tests include a large number of genotypes, so field trials for all possible combinations of potential crop ideotypes would be prohibitive. However, incorporating crown structural attributes into our growth models may enhance our ability to simulate the growth implications of novel combinations of heritable traits that cannot be field tested.


 ,
, 


{kind=link}
{kind=link}