
Assessing the Global Pest Risk of Aeolesthes sarta with Regards to the Host Specie Populus alba under Climate Change Scenarios


Abstract
:1. Introduction
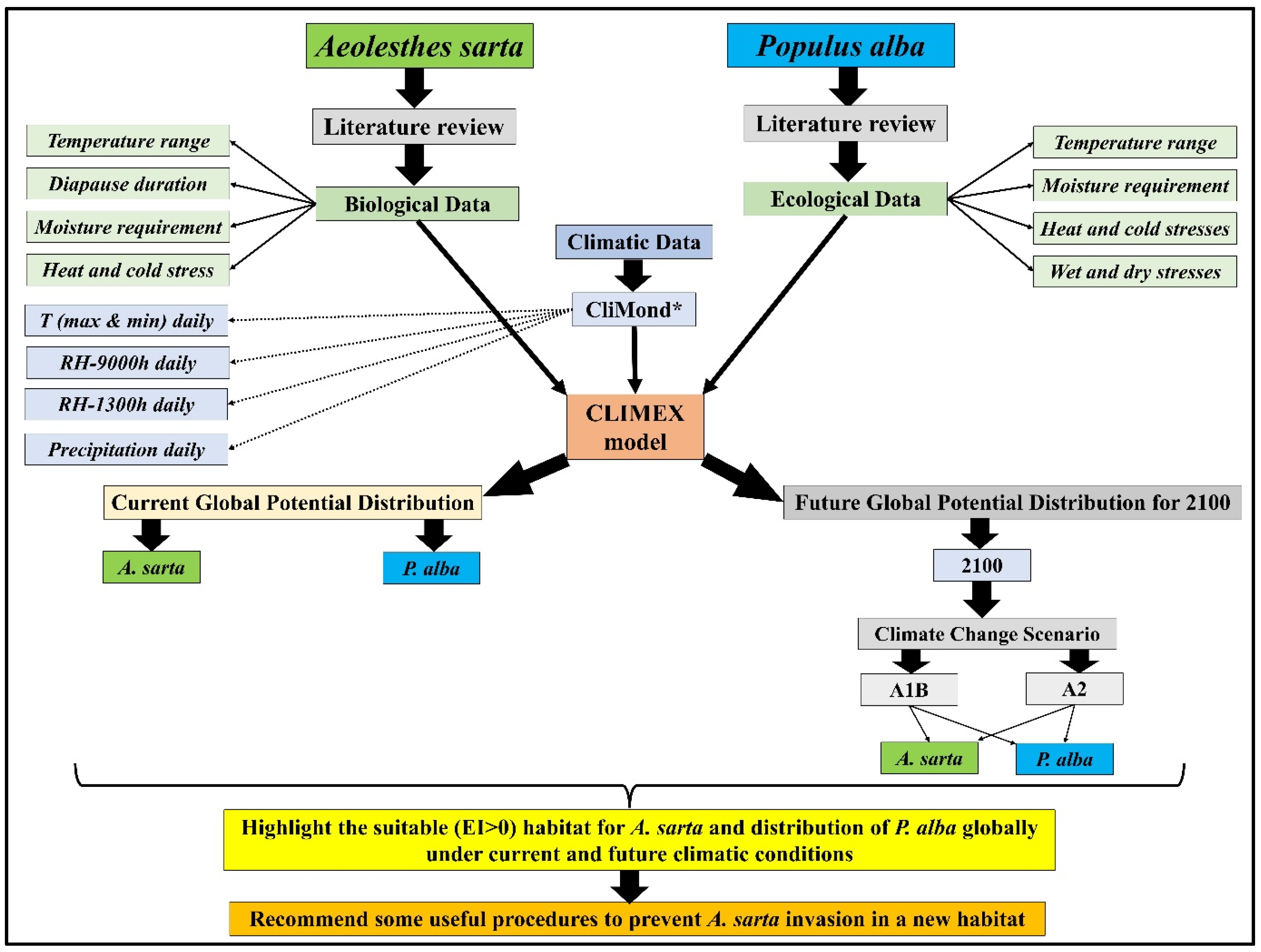
2. Materials and Methods
2.1. CLIMEX Model
2.2. Climatic Data
2.3. Aeolesthes sarta and Populus alba Global Distribution Data
2.4. Pest (Aelosthes sarta) Biological Data
2.4.1. Temperature Index
2.4.2. Diapause Index
2.4.3. Moisture Index
2.4.4. Stress Index
2.5. Host (Populus alba) Ecological Data
2.5.1. Temperature Index
2.5.2. Moisture Index
2.5.3. Stress Index
2.6. Model Verification
2.7. Map Generation
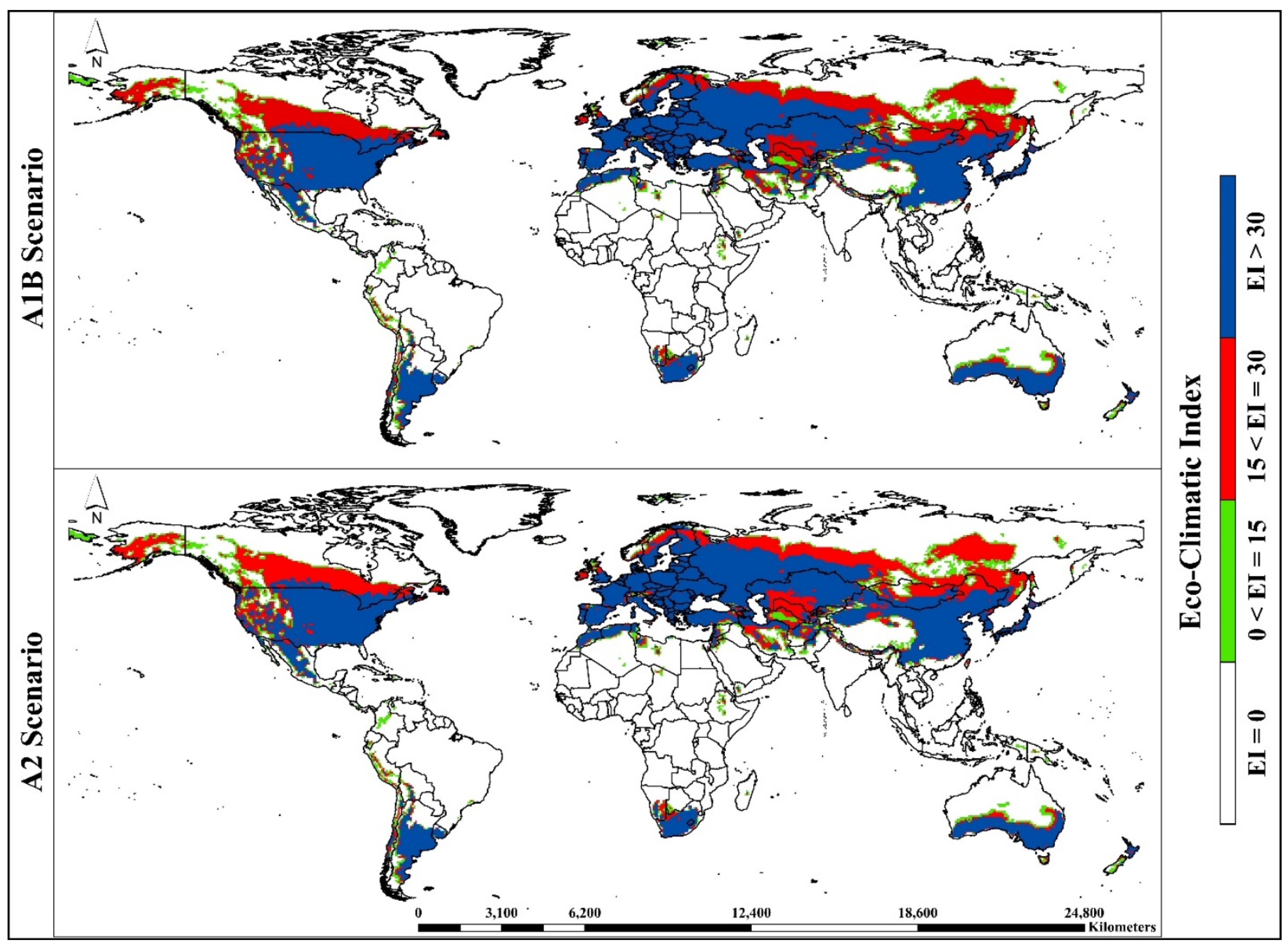
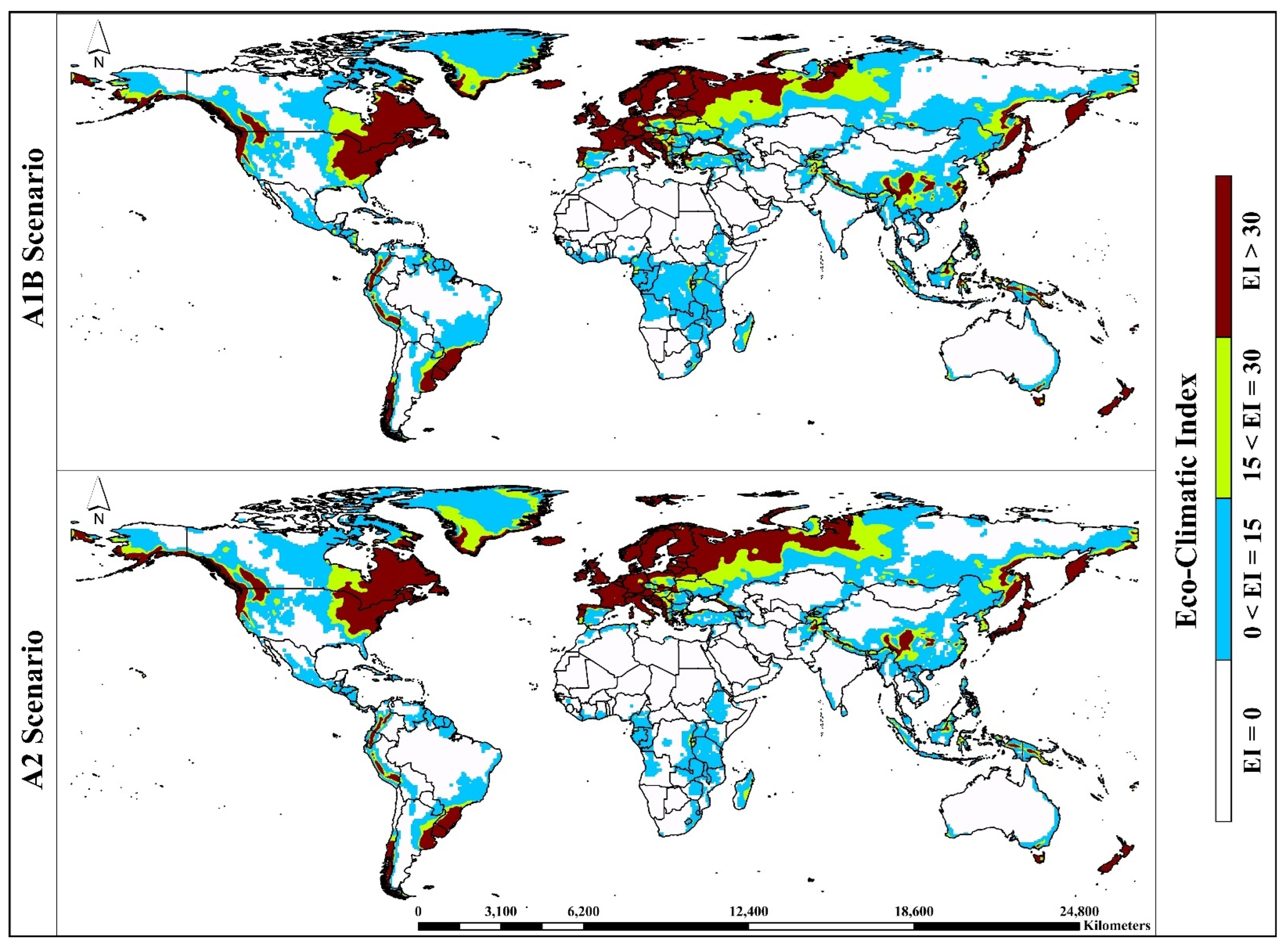
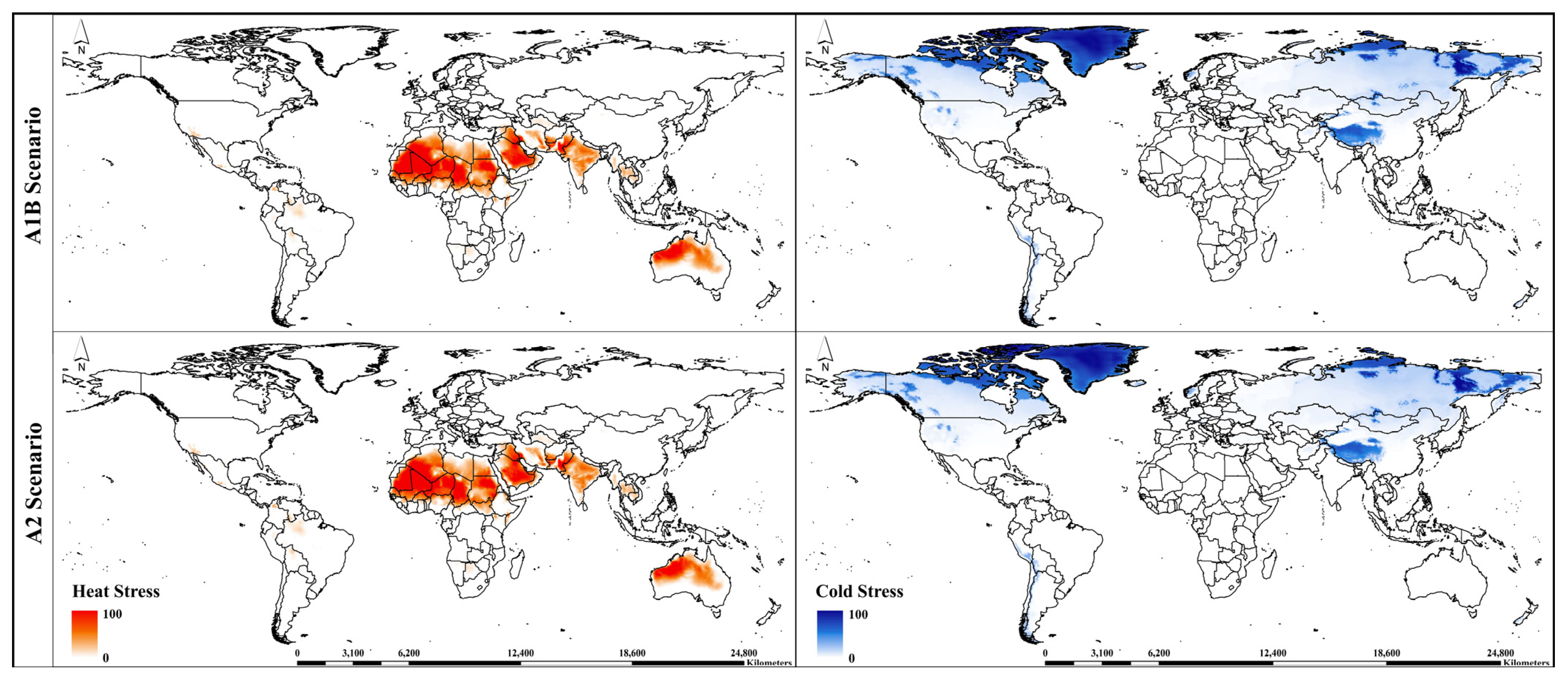
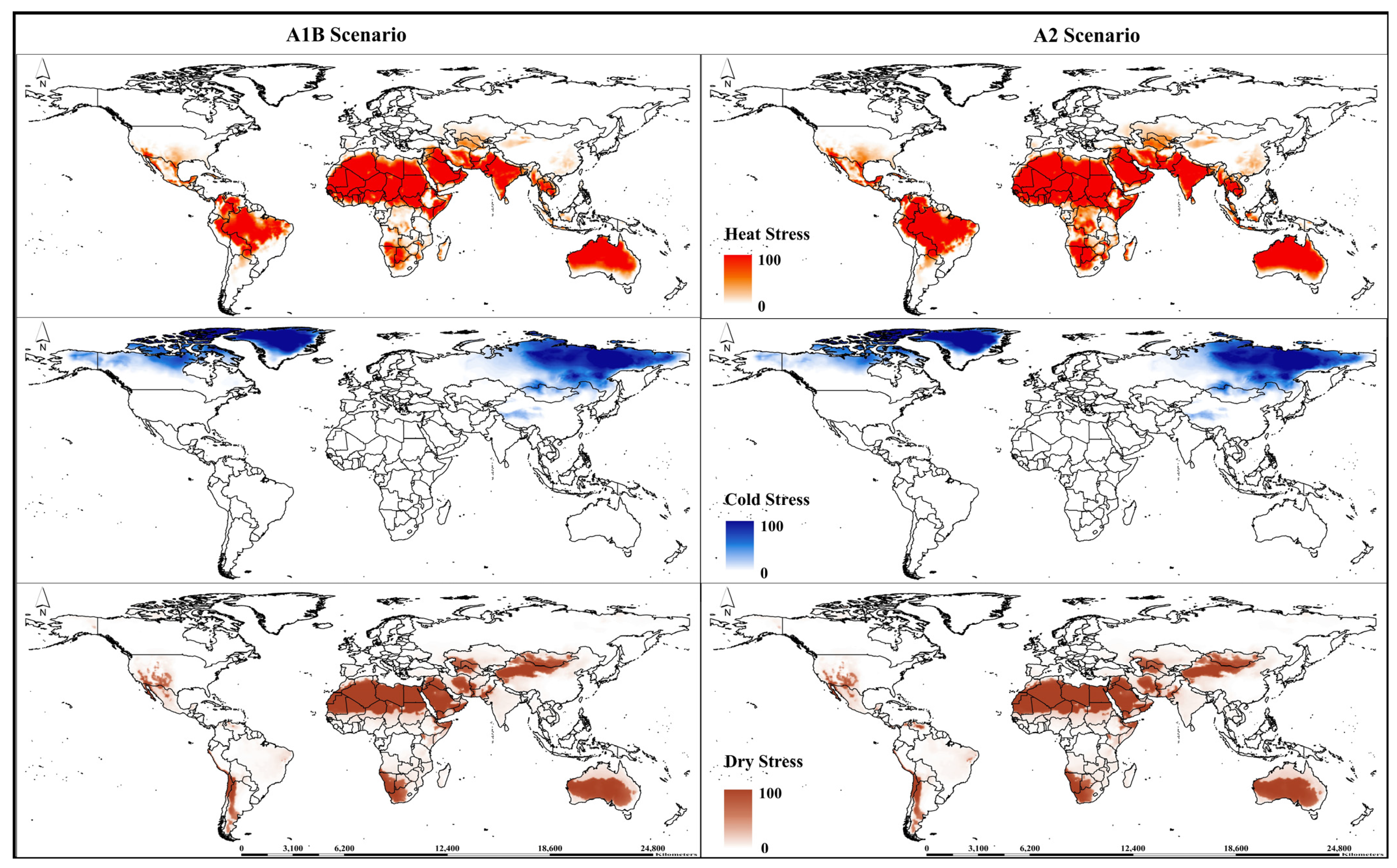
3. Results
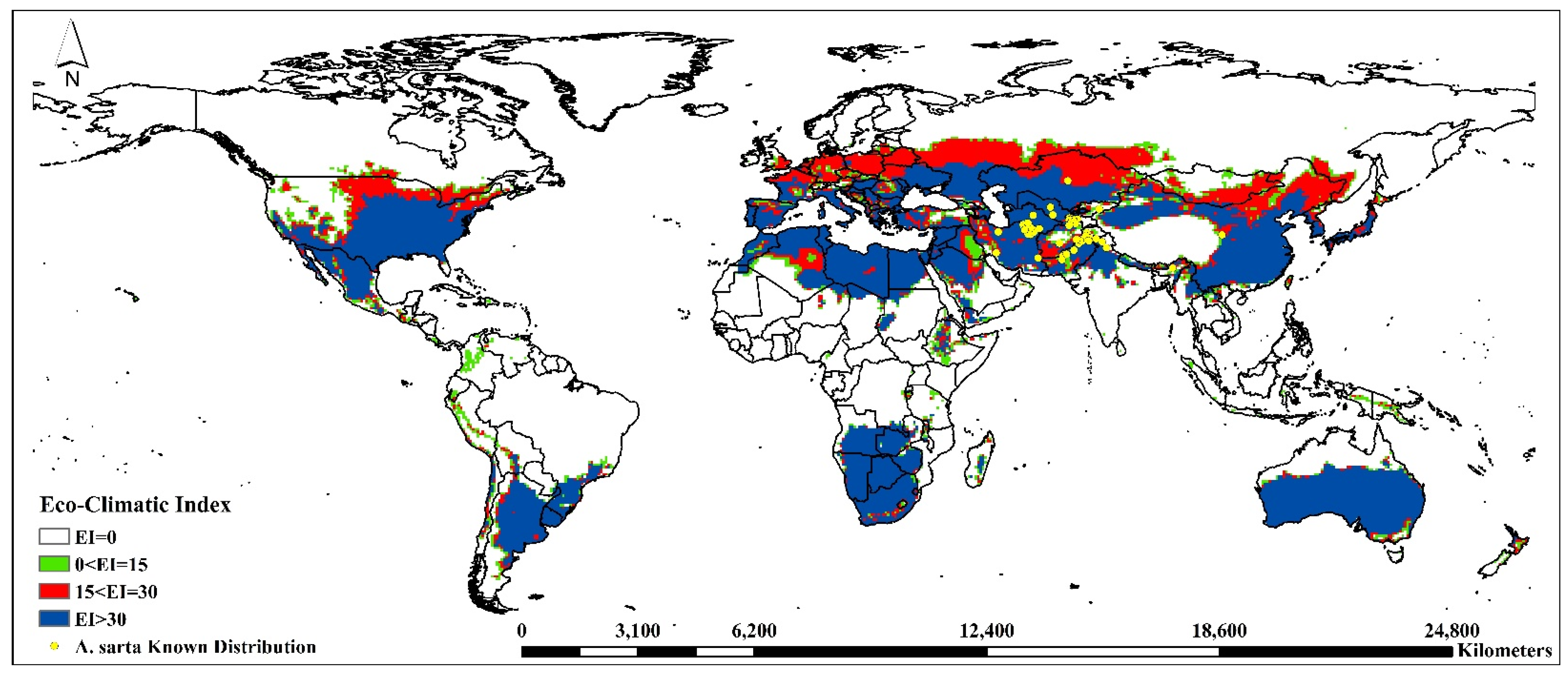
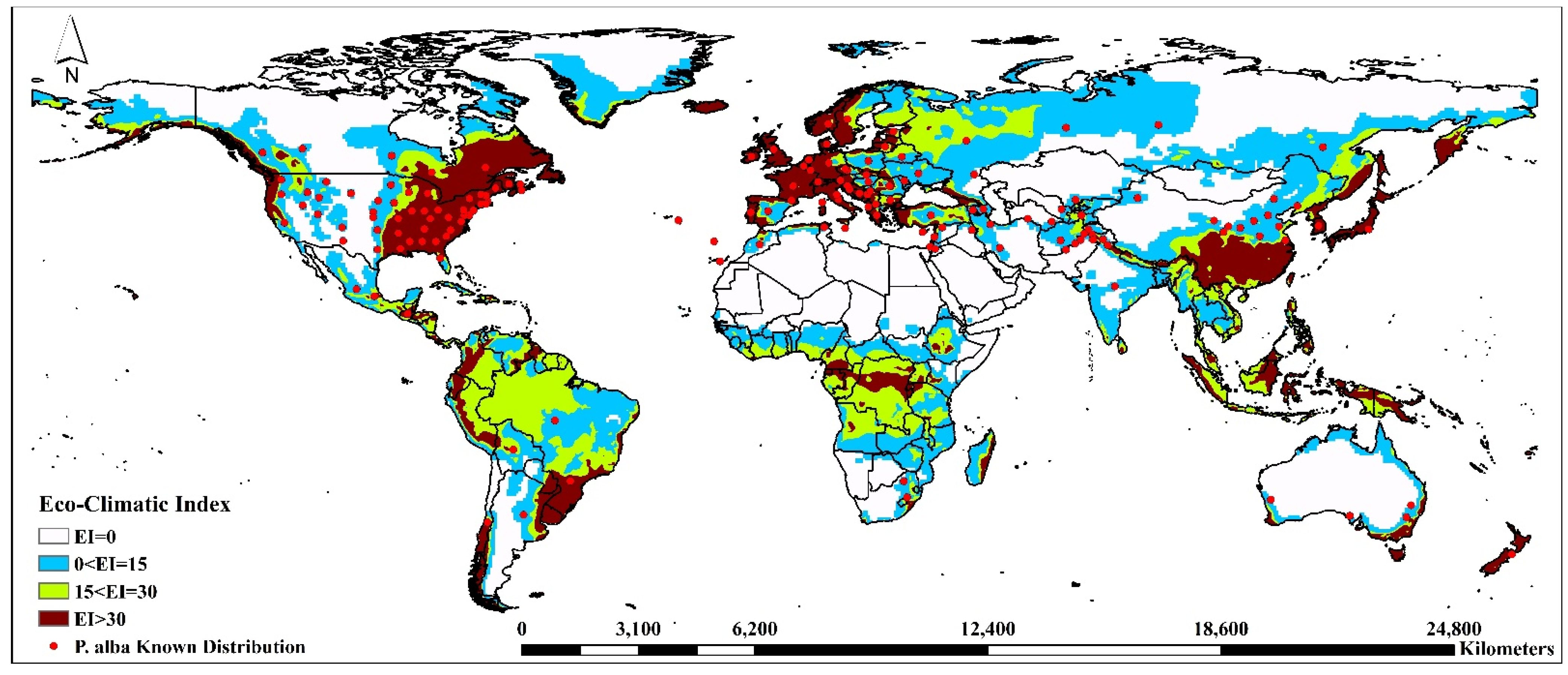
3.1. Model Fitting/Verification and Current Global Distribution
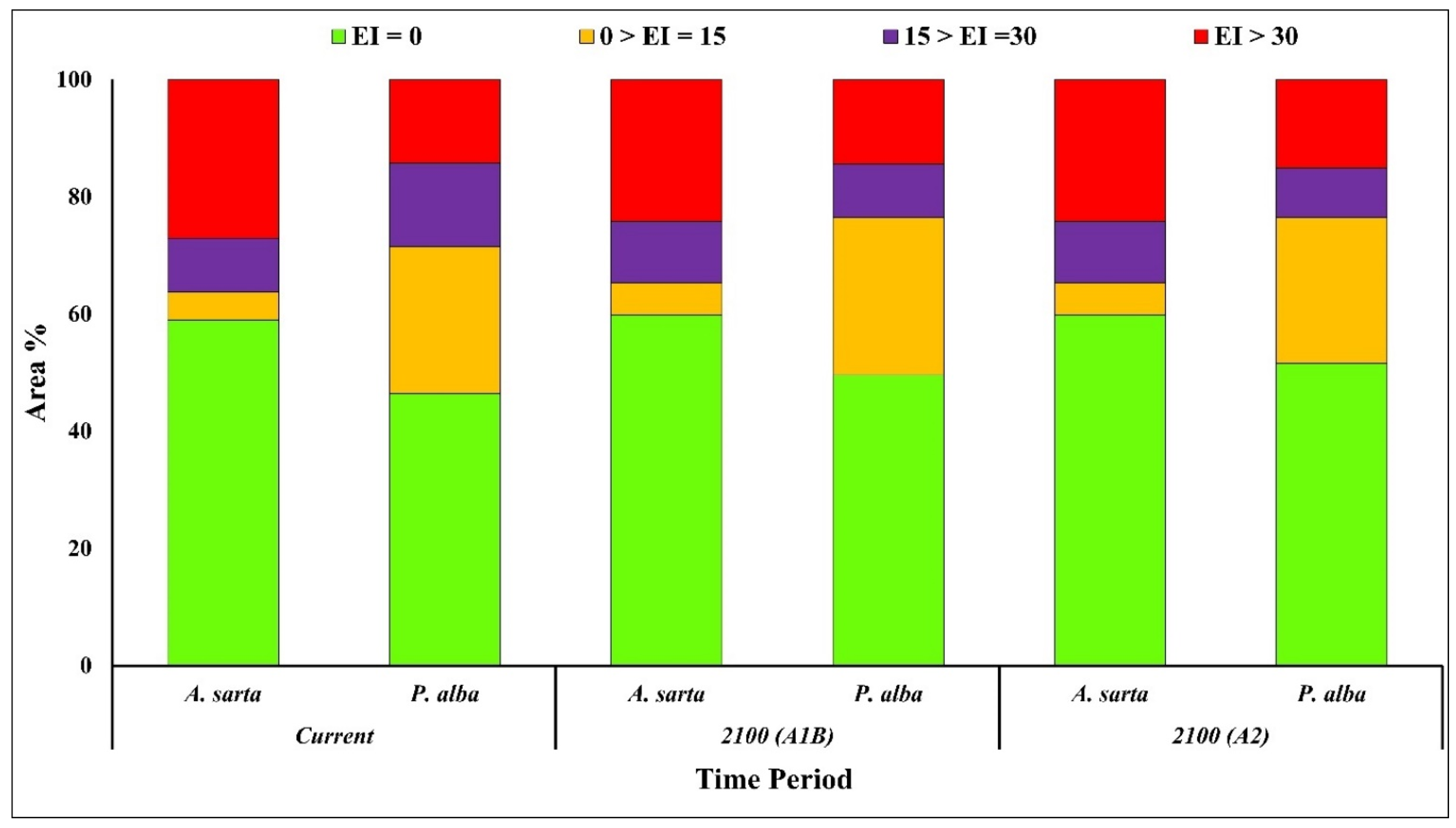
3.2. Potential Global Distribution under Future Climate Conditions
3.3. Continent-Wise Distribution under Future Climate Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| CCS | Climate Change Scenario |
| EI | Ecoclimatic index |
References
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of IPCC the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhao, Q.; Wei, J.; Zhang, H. Study on the Potential Distribution of Leptinotarsa decemlineata and Its Natural Enemy Picromerusbidens under Climate Change. Front. Ecol. Evol. 2022, 9, 786436. [Google Scholar] [CrossRef]
- Wei, J.; Peng, L.; He, Z.; Lu, Y.; Wang, F. Potential distribution of two invasive pineapple pests under climate change. Pest Manag. Sci. 2020, 76, 1652–1663. [Google Scholar] [CrossRef] [PubMed]
- Hayat, U. City longhorn beetle (Aeolesthes sarta): A review of the species, its morphology, distribution, da-mage, prevention and control. J. For. Sci. 2022, 68, 199–212. [Google Scholar] [CrossRef]
- Ślipiński, A.; Escalona, H. Australian Longhorn Beetles (Coleoptera: Cerambycidae). In Subfamily Cerambycinae; CSIRO Publishing: Clayton, Australia, 2016; Volume 2, p. 640. [Google Scholar] [CrossRef]
- Khan, S.A.; Bhatia, S.; Tripathi, N. Entomological investigation on Aeolesthes sarta (Solsky), a major pest on walnut trees (Juglans regia L.) in Kashmir valley. J. Acad. Ind. Res. 2013, 2, 325. [Google Scholar]
- Mazaheri, A.; Khajehali, J.; Marzieh, K.; Hatami, B. Laboratory and field evaluation of insecticides for the control of Aeolesthes sarta Solsky (Col.: Cerambycidae). J. Crop Prot. 2015, 4, 257–266. [Google Scholar]
- Morewood, W.D.; Hoover, K.; Neiner, P.R.; McNeil, J.R.; Sellmer, J.C. Host tree resistance against the polyphagous wood-boring beetle Anoplophora glabripennis. Entomol. Exp. Appl. 2004, 110, 79–86. [Google Scholar] [CrossRef]
- Poland, T.M.; Haack, R.A.; Petrice, T.R.; Miller, D.L.; Bauer, L.S.; Gao, R. Field evaluations of systemic insecticides for control of Anoplophora glabripennis (Coleoptera: Cerambycidae) in China. J. Econ. Entomol. 2006, 99, 383–392. [Google Scholar] [CrossRef]
- Farashiani, M.E.; Shamohammadi, D.; Sadeghi, S.E. Biological study of Sart long horn beetle, Aeolesthes sarta Solsky (Coleoptera: Cerambycidae) in the laboratory. J. Entomol. Soc. Iran 2000, 20, 77–90. [Google Scholar]
- Orlinskii, A.D. Outcomes of the EPPO project on quarantine pests for forestry 1. EPPO Bull. 2006, 36, 497–511. [Google Scholar] [CrossRef]
- Vanhanen, H.; Veteli, T.O.; Niemelä, P. Potential distribution ranges in Europe for Aeolesthes sarta, Tetropium gracilicorne and Xylotrechus altaicus, a CLIMEX analysis. EPPO Bull. 2008, 38, 239–248. [Google Scholar] [CrossRef]
- Hayat, U.; Qin, H.; Zhao, J.; Akram, M.; Shi, J.; Ya, Z. Variation in the potential distribution of Agrotis ipsilon (Hufnagel) globally and in Pakistan under current and future climatic conditions. Plant Prot. Sci. 2021, 57, 148–158. [Google Scholar] [CrossRef]
- CABI (2022): Trirachys Sartus—Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/3430 (accessed on 8 February 2023).
- Ištok, I.; Šefc, B.; Hasan, M.; Popović, G.; Sedlar, T. Fiber characteristics of white poplar (Populus alba L.) juvenile wood along the Drava river. Drv. Ind. 2017, 68, 241–247. [Google Scholar] [CrossRef]
- Ostry, M.E. A Guide to Insect, Disease, and Animal Pests of Poplars (No. 677); US Department of Agriculture, Forest Service: Washington, DC, USA, 1989; pp. 1–120. [Google Scholar]
- Ball, J.; Carle, J.; Del Lungo, A. Contribution of poplars and willows to sustainable forestry and rural development. UNASYLVA-FAO. 2005, 56, pp. 3–9. Available online: https://www.fao.org/3/a0026e/a0026e03.pdf (accessed on 15 March 2023).
- Farashiani, M.E.; Sadeghi, S.E.; Abaii, M. Geographic distribution and hosts of Sart longhorn beetle, Aeolesthes sarta Solsky (Col.: Cerambycidae) in Iran. J. Entomol. Soc. Iran 2001, 20, 81–96. [Google Scholar]
- Ahmad, M.I.; Hafiz, I.A.; Chaudhry, M.I. Biological studies on Aeolesthes sartus Solsky attacking poplars in Pakistan. Pak. J. For. 1977, 27, 122–129. [Google Scholar]
- Gul, H.; Chaudhry, M.I. Some observations on natural enemies of poplar borers in Pakistan. Pak. J. For. 1992, 42, 214–222. [Google Scholar]
- Arshad, M.; Hafiz, I.A. Microbial trials of a pathogenic fungus, Beauveria bassiana (Bals.) Vuill. against the adults of Aeolesthes sartus Solsky (Cerambycidae: Coleoptera). Pak. J. Zool. 1983, 15, 213–215. [Google Scholar]
- Isebrands, J.G.; Richardson, J. Poplars and Willows: Trees for Society and the Environment; CABI International: Wallingford, UK; FAO: Roma, Italy, 2014; pp. 1–7. [Google Scholar] [CrossRef]
- Cocquempot, C.; Lindelöw, Å. Longhorn beetles (Coleoptera, Cerambycidae). In Alien Terrestrial Arthropods of Europe; Roques, A., Ed.; BioRisk: Singapore, 2010; Chapter 8.1; Volume 4, pp. 193–218. [Google Scholar] [CrossRef]
- Cocquempot, C. Alien longhorned beetles (Coleoptera Gerambycidae): Original interceptions and introductions in Europe, mainly in France, and notes about recently imported species. Redia 2006, 89, 35–50. [Google Scholar]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/AR5_SYR_FINAL_Front_matters.pdf (accessed on 10 February 2023).
- Tamura, Y.; Osawa, T.; Tabuchi, K.; Yamasaki, K.; Niiyama, T.; Sudo, S.; Ishigooka, Y.; Yoshioka, A.; Takada, M.B. Estimating plant–insect interactions under climate change with limited data. Sci. Rep. 2022, 12, 10554. [Google Scholar] [CrossRef] [PubMed]
- Damos, P.; Savopoulou-Soultani, M. Temperature-driven models for insect development and vital thermal requirements. Psyche 2012, 2012, 123405. [Google Scholar] [CrossRef] [Green Version]
- Sutherst, R.W.; Maywald, G.F.; Russell, B.L. Estimating vulnerability under global change: Modular modelling of pests. Agric. Ecosyst. Environ. 2000, 82, 303–319. [Google Scholar] [CrossRef]
- Sutherst, R.W.; Maywald, G.F.; Kriticos, D.J. CLIMEX Version 3: User’s Guide; Hearne Scientific Software Pty Ltd.: Victoria, Australia, 2007. [Google Scholar]
- Sutherst, R.W.; Maywald, G.F.; Bourne, A.S. Including species interactions in risk assessments for global change. Glob. Chang. Biol. 2007, 13, 1843–1859. [Google Scholar] [CrossRef]
- Hayat, U.; Akram, M.; Kour, S.; Arif, T.; Shi, J. Pest Risk Assessment of Aeolesthes sarta (Coleoptera: Cerambycidae) in Pakistan under Climate Change Scenario. Forests 2023, 14, 253. [Google Scholar] [CrossRef]
- Nevidomov, A.M. Ecophytocoenotic patterns in the distribution of poplar forests in the floodplains of southeastern European Russia. Bot. Zhurnal 1994, 79, 47–58. [Google Scholar]
- Modir-Rahmati, A. Cultivation of poplar in Iran. Holzzucht 1997, 51, 41–43. [Google Scholar]
- Pasiecznik, N.M.; Smith, I.M.; Watson, G.W.; Brunt, A.A.; Ritchie, B.; Charles, L.M.F. CABI/EPPO distribution maps of plant pests and plant diseases and their important role in plant quarantine. EPPO Bull. 2005, 35, 1–7. [Google Scholar] [CrossRef]
- Afshan, N.S.; Iqbal, S.H.; Khalid, A.N.; Niazi, A.R. Some additions to the uredinales of Azad Jammu and Kashmir (AJ & K), Pakistan. Pak. J. Bot. 2011, 43, pp. 1373–1379. Available online: http://www.pakbs.org/pjbot/PDFs/43(2)/PJB43(2)1373.pdf (accessed on 28 March 2023).
- Pasiecznik, N. ‘Populus alba (Silver-Leaf Poplar)’, CABI Compendium; CABI International: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Kour, S.; Khurma, U.; Brodie, G.; Singh, S. Modeling the potential global distribution of suitable habitat for the biological control agent Heterorhabditis indica. Ecol. Evol. 2022, 12, e8997. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Reisinger, A., Eds.; IPCC: Geneva, Switzerland, 2007; p. 104. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/ar4_syr_full_report.pdf (accessed on 25 February 2022).
- Sekawin, M. Genetics of Populus alba. Ann. For. 1975, 6, 159–189. [Google Scholar]
- Gucker, C.L. Populus alba and hybrids. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Missoula, MT, USA, 2010. Available online: https://www.fs.usda.gov/database/feis/plants/tree/popspp/all.html (accessed on 15 March 2023).
- Domingo, I.L.; Gordon, J.C. Physiological responses of an aspen-poplar hybrid to air temperature and soil moisture. Bot. Gaz. 1974, 135, pp. 184–192. Available online: https://www.journals.uchicago.edu/doi/abs/10.1086/336750 (accessed on 16 March 2023).
- Rédei, K. Early evaluation of promising white poplar (Populus alba L.) clones in sandy ridges between the rivers Danube and Tisza in Hungary. Silva Lusit. 1998, 6, 63–71. [Google Scholar]
- Aljaryian, R.; Kumar, L.; Taylor, S. Modelling the current and potential future distributions of the sunn pest Eurygaster integriceps (Hemiptera: Scutelleridae) using CLIMEX. Pest Manag. Sci. 2016, 72, 1989–2000. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zhao, Q.; Zhao, W.; Zhang, H. Predicting the potential distributions of the invasive cycad scale Aulacaspis yasumatsui (Hemiptera: Diaspididae) under different climate change scenarios and the implications for management. PeerJ 2018, 6, e4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, W.J.; Haack, R.A. The role of drought in outbreaks of plant-eating insects. Bioscience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- McIntyre, N.E. Ecology of urban arthropods: A review and a call to action. Ann. Entomol. Soc. Am. 2000, 93, 825–835. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2022, 416, 389–395. [Google Scholar] [CrossRef]
- Battisti, A. Forests and climate change—Lessons from insects. Forest 2004, 1, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Niemelä, P.; Mattson, W.J. Invasion of North American forests by European phytophagous insects. BioScience 1996, 46, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Roques, A.; Auger-Rozenberg, M.A.; Boivin, S. A lack of native congeners may limit colonization of introduced conifers by indigenous insects in Europe. Can. J. For. Res. 2006, 36, 299–313. [Google Scholar] [CrossRef]
- Allen, E.; Noseworthy, M.; Ormsby, M. Phytosanitary measures to reduce the movement of forest pests with the international trade of wood products. Biol. Invasions 2017, 19, 3365–3376. [Google Scholar] [CrossRef]
- Dhiman, R.C. Availability of poplar wood-a boon for plywood industry. Plywood Gaz. 2004, 64–72. [Google Scholar]
- EPPO. Trirachys sartus. EPPO Datasheets on Pests Recommended for Regulation. 2023. Available online: https://gd.eppo.int (accessed on 8 June 2023).
- Liebhold, A.M. Alien species as agents of global change ecology and management of the gypsy moth in north america as a case history. In Proceedings: International Symposium of the Kanazawa University 21st Century COE Program; Kamata, N., Ed.; Kanazawa University: Kanazawa, Japan, 2003; Volume 1, pp. 71–75. [Google Scholar]
- Akram, M.; Hayat, U.; Shi, J.; Anees, S.A. Association of the Female Flight Ability of Asian Spongy Moths (Lymantria dispar asiatica) with Locality, Age and Mating: A Case Study from China. Forests 2022, 13, 1158. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D. Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-2007-0. [Google Scholar]
- Webber, B.L.; Yates, C.J.; Le Maitre, D.C.; Scott, J.K.; Kriticos, D.J.; Ota, N.; McNeill, A.; Le Roux, J.J.; Midgley, G.F. Modelling horses for novel climate courses: Insights from projecting potential distributions of native and alien Australian acacias with correlative and mechanistic models. Divers. Distrib. 2011, 17, 978–1000. [Google Scholar] [CrossRef]
- Khan, A.A.; Kundoo, A.A. Pests of walnut. In Pests and Their Management; Springer: Singapore, 2018; pp. 605–647. [Google Scholar] [CrossRef]
- Hussain, A.; Muhammad, A.; Hayat, U.; Ahmad, B.; Murtaza, A.M.; Khalid, K.M.B.; Ullah, S. Response of rice cultivars and insecticides against Rice stem borer (Scirpophaga incertulus) in Pakistan (Swat). J. Biodivers. Environ. Sci. 2019, 15, pp. 88–94. Available online: https://www.innspub.net/wp-content/uploads/2022/04/JBES-V15-No2-p88-94.pdf (accessed on 28 March 2023).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eco-Climatic Index | Representation |
|---|---|
| EI = 0 | Unsuitable habitat—The climate and environment are not conducive to the growth and establishment of A. sarta and P. alba. |
| 0 < EI = 15 | Marginally suitable habitat—The climate and environment are marginally conducive to the growth and establishment of A. sarta and P. alba. |
| 15 < EI = 30 | Suitable habitat—The climate and environment are conducive to the growth and establishment of A. sarta and P. alba. |
| EI > 30 | Highly suitable habitat—The climate and environment are highly conducive to the growth and establishment of A. sarta and P. alba. |
| Climate Change Scenarios | Period | Data Set | Source |
|---|---|---|---|
| Current | CM10_1975H_CX_MM_V1.2 | Climond | |
| A1B * | 2100 | CM10_2100_A1B_CS_Cx_MM_V1.2 | Climond |
| A2 ** | 2100 | CM10_2100_A2_CS_Cx_MM_V1.2 | Climond |
| Parameters | Code | Values | |
|---|---|---|---|
| Aelosthes sarta | Populus alba | ||
| Temperature | |||
| Limiting low temperature (°C) | DV0 | 10 | −5 |
| Lower optimal temperature (°C) | DV1 | 15 | 4 |
| Upper optimal temperature (°C) | DV2 | 37 | 20 |
| Limiting high temperature (°C) | DV3 | 40 | 35 |
| Moisture Index | |||
| Limiting low soil moisture | SM0 | 0 | 0.5 |
| Lower optimal soil moisture | SM1 | 0.001 | 1 |
| Upper optimal soil moisture | SM2 | 1.5 | 1.5 |
| Limiting high soil moisture | SM3 | 2.5 | 2 |
| Diapause Index | |||
| Diapause induction day length | DPD0 | 12 | - |
| Diapause induction temperature (°C) | DPT0 | 13 | - |
| Diapause termination temperature (°C) | DPT1 | 10 | - |
| Diapause development days | DPD | 90 | - |
| Summer or winter Diapause | DPSW | 0 | - |
| Cold Stress | |||
| CS temperature threshold (°C) | TTCS | 9 | −15 |
| CS temperature rate | THCS | –0.00001 | −0.0001 |
| Heat Stress | |||
| HS temperature threshold (°C) | TTHS | 41 | 35 |
| HS temperature rate | THHS | 0.005 | 0.005 |
| Wet Stress | |||
| Wet stress threshold | SMWS | - | 2.5 |
| Wet stress accumulation rate | HWS | - | 0.002 |
| Dry Stress | |||
| Dry stress threshold | SMDS | - | 0.2 |
| Dry stress accumulation rate | HDS | - | −0.005 |
| Population degree day | PDD | 700 | - |
| Eco-Climatic Index | Asia | Europe | Africa | North America | South America | Oceania | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | Mi. km2 | % | Mi. km2 | % | Mi. km2 | % | Mi. km2 | % | Mi. km2 | % | Mi. km2 | |||
| Current | EI = 0 | A. sarta | 53.81 | 23.46 | 37.59 | 3.60 | 63.70 | 18.84 | 65.06 | 15.30 | 77.62 | 13.53 | 29.60 | 2.32 |
| P. alba | 53.21 | 26.41 | 5.44 | 0.72 | 56.58 | 13.32 | 53.10 | 18.17 | 15.19 | 2.15 | 74.10 | 5.04 | ||
| 0 < EI = 15 | A. sarta | 5.96 | 2.60 | 5.61 | 0.54 | 3.74 | 1.11 | 4.36 | 1.03 | 5.05 | 0.88 | 3.53 | 0.28 | |
| P. alba | 27.39 | 13.59 | 39.70 | 5.25 | 20.12 | 4.74 | 20.21 | 6.91 | 28.21 | 3.99 | 12.66 | 0.86 | ||
| 15 < EI = 30 | A. sarta | 13.44 | 5.86 | 28.35 | 2.72 | 3.28 | 0.97 | 8.27 | 1.94 | 2.78 | 0.48 | 2.92 | 0.23 | |
| P. alba | 9.25 | 4.59 | 24.26 | 3.21 | 17.73 | 4.17 | 7.85 | 2.69 | 38.53 | 5.45 | 2.23 | 0.15 | ||
| EI > 30 | A. sarta | 26.79 | 11.68 | 28.44 | 2.73 | 29.28 | 8.66 | 22.31 | 5.25 | 14.55 | 2.54 | 63.95 | 5.01 | |
| P. alba | 10.16 | 5.04 | 30.60 | 4.05 | 5.57 | 1.31 | 18.84 | 6.44 | 18.07 | 2.56 | 11.00 | 0.75 | ||
| 2100 (A1B) | EI = 0 | A. sarta | 49.09 | 21.40 | 7.29 | 0.70 | 91.06 | 26.93 | 43.38 | 10.20 | 82.99 | 14.46 | 60.34 | 4.73 |
| P. alba | 52.60 | 26.11 | 3.48 | 0.46 | 72.90 | 17.16 | 36.04 | 12.33 | 59.43 | 8.41 | 84.93 | 5.78 | ||
| 0 < EI = 15 | A. sarta | 7.35 | 3.20 | 3.38 | 0.32 | 2.18 | 0.65 | 7.07 | 1.66 | 4.96 | 0.87 | 7.21 | 0.56 | |
| P. alba | 29.68 | 14.73 | 13.84 | 1.83 | 25.23 | 5.94 | 34.18 | 11.69 | 24.24 | 3.43 | 7.03 | 0.48 | ||
| 15 < EI = 30 | A. sarta | 16.45 | 7.17 | 10.39 | 1.00 | 1.64 | 0.48 | 18.22 | 4.28 | 2.52 | 0.44 | 6.03 | 0.47 | |
| P. alba | 7.19 | 3.57 | 23.23 | 3.07 | 1.63 | 0.38 | 9.68 | 3.31 | 4.92 | 0.70 | 2.01 | 0.14 | ||
| EI > 30 | A. sarta | 27.11 | 11.82 | 78.94 | 7.57 | 5.12 | 1.51 | 31.34 | 7.37 | 9.52 | 1.66 | 26.41 | 2.07 | |
| P. alba | 10.53 | 5.23 | 59.45 | 7.87 | 0.24 | 0.06 | 20.10 | 6.88 | 11.41 | 1.61 | 6.03 | 0.41 | ||
| 2100 (A2) | EI = 0 | A. sarta | 49.09 | 21.40 | 7.29 | 0.70 | 91.06 | 26.93 | 43.38 | 10.20 | 82.99 | 14.46 | 60.34 | 4.73 |
| P. alba | 53.23 | 26.42 | 3.77 | 0.50 | 82.61 | 19.44 | 33.48 | 11.45 | 66.34 | 9.39 | 85.85 | 5.84 | ||
| 0 < EI = 15 | A. sarta | 7.35 | 3.20 | 3.38 | 0.32 | 2.18 | 0.65 | 7.07 | 1.66 | 4.96 | 0.87 | 7.21 | 0.56 | |
| P. alba | 29.52 | 14.65 | 12.40 | 1.64 | 16.42 | 3.86 | 34.72 | 11.88 | 19.03 | 2.69 | 7.34 | 0.50 | ||
| 15 < EI = 30 | A. sarta | 16.45 | 7.17 | 10.39 | 1.00 | 1.64 | 0.48 | 18.22 | 4.28 | 2.52 | 0.44 | 6.03 | 0.47 | |
| P. alba | 9.61 | 4.77 | 19.30 | 2.55 | 0.90 | 0.21 | 10.85 | 3.71 | 4.16 | 0.59 | 1.44 | 0.10 | ||
| EI > 30 | A. sarta | 27.11 | 11.82 | 78.94 | 7.57 | 5.12 | 1.51 | 31.34 | 7.37 | 9.52 | 1.66 | 26.41 | 2.07 | |
| P. alba | 7.64 | 3.79 | 64.52 | 8.54 | 0.08 | 0.02 | 20.96 | 7.17 | 10.46 | 1.48 | 5.37 | 0.37 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayat, U.; Kour, S.; Akram, M.; Shi, J.; Wiarta, R. Assessing the Global Pest Risk of Aeolesthes sarta with Regards to the Host Specie Populus alba under Climate Change Scenarios. Forests 2023, 14, 1260. https://doi.org/10.3390/f14061260
Hayat U, Kour S, Akram M, Shi J, Wiarta R. Assessing the Global Pest Risk of Aeolesthes sarta with Regards to the Host Specie Populus alba under Climate Change Scenarios. Forests. 2023; 14(6):1260. https://doi.org/10.3390/f14061260
Chicago/Turabian StyleHayat, Umer, Sumeet Kour, Muhammad Akram, Juan Shi, and Rinto Wiarta. 2023. "Assessing the Global Pest Risk of Aeolesthes sarta with Regards to the Host Specie Populus alba under Climate Change Scenarios" Forests 14, no. 6: 1260. https://doi.org/10.3390/f14061260