


Phenotypic Diversity Analysis in Elaeagnus angustifolia Populations in Gansu Province, China

Abstract
:1. Introduction
2. Materials and Methods
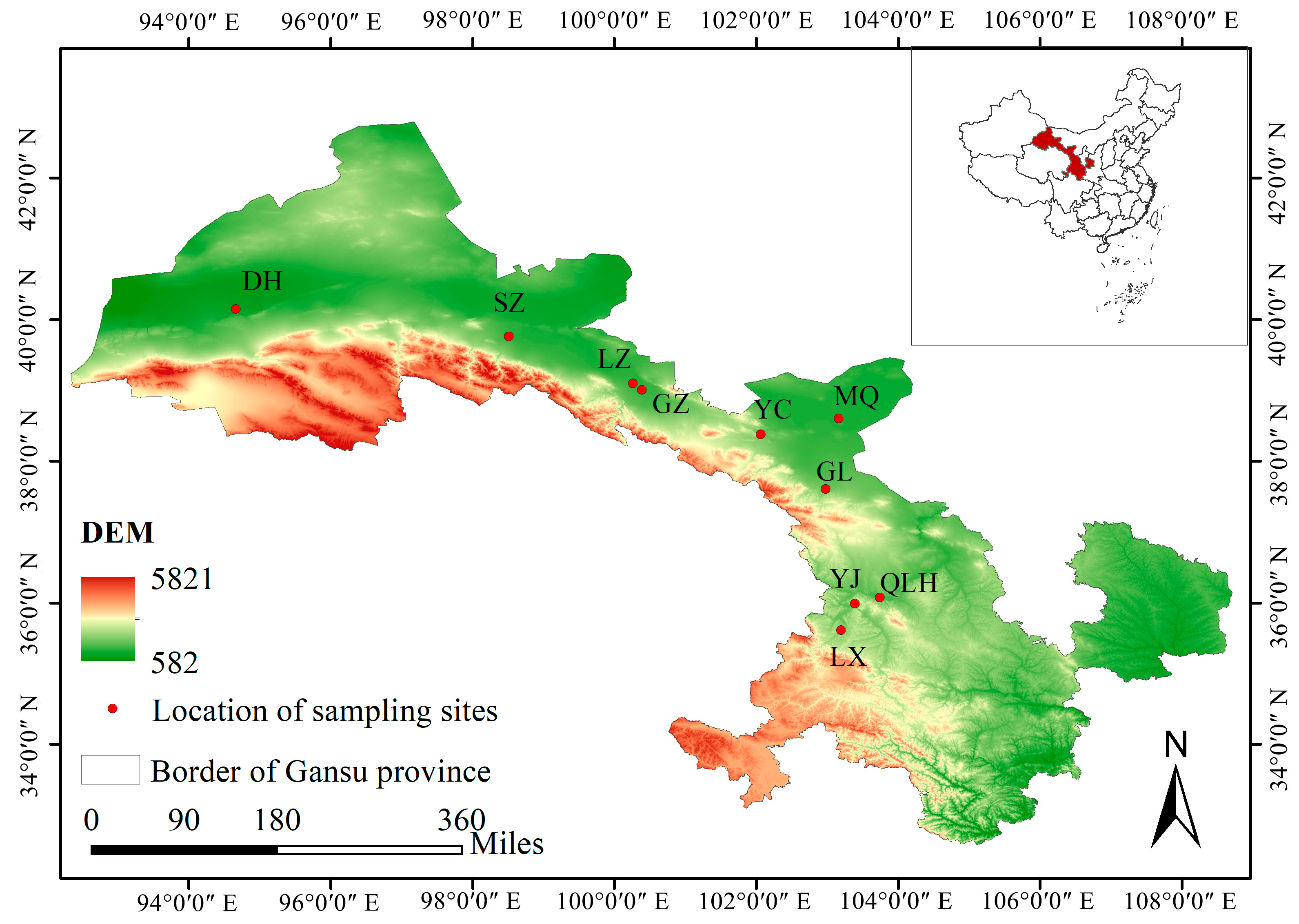
2.1. Plant Material
2.2. Morphometric Analysis
2.3. Statistical Analysis
2.3.1. Analysis of Variance
2.3.2. Phenotypic Differentiation Coefficient
2.3.3. Descriptive Statistics
2.3.4. Diversity Index
2.3.5. Multivariate Analysis
3. Results
3.1. Phenotypic Differences among and within Populations
3.2. Variation Degree of Phenotypic Traits
3.3. Diversity Index of Phenotypic Traits
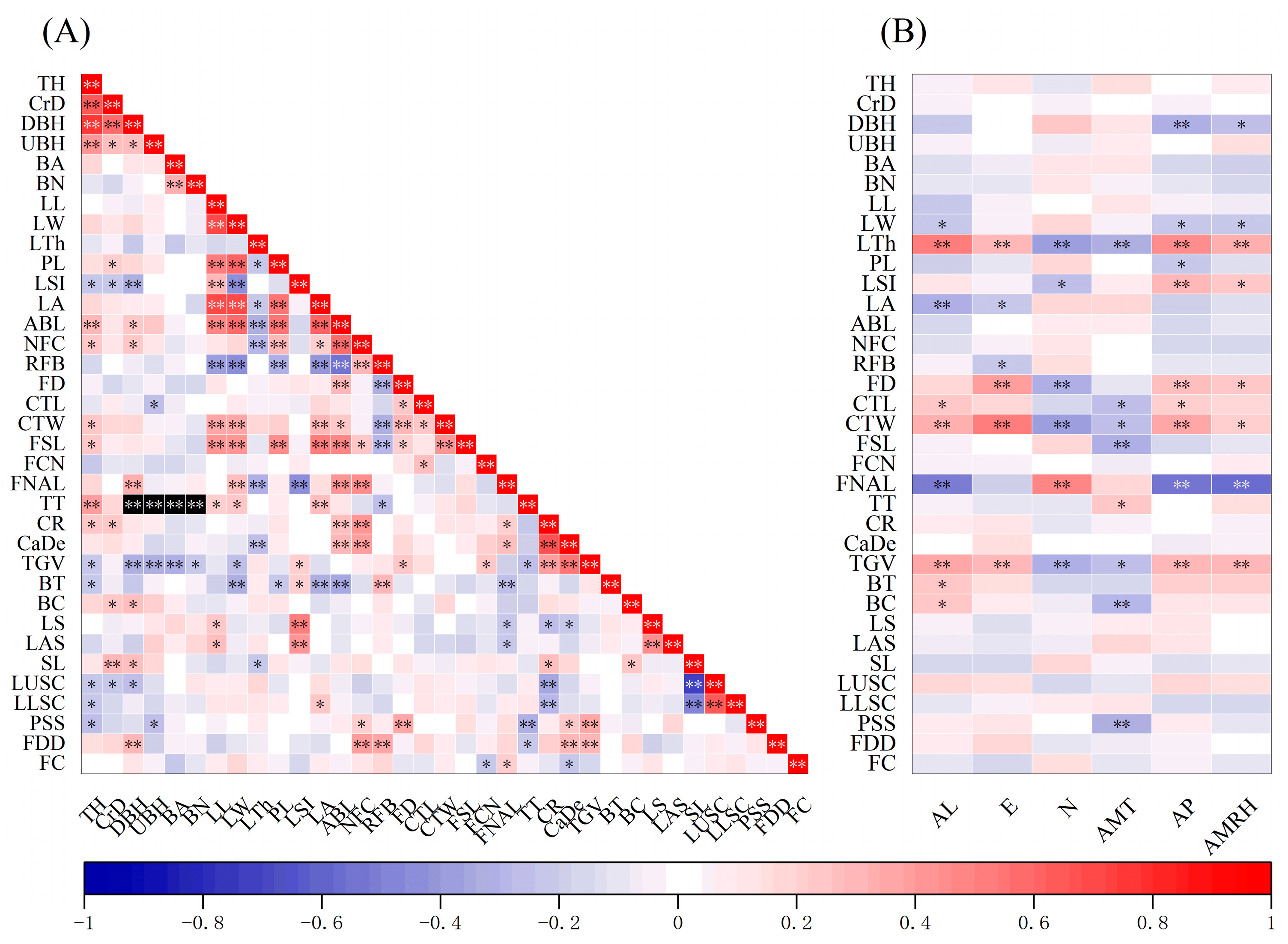
3.4. Correlations among Phenotypic Traits
3.5. Correlation between Phenotypic Traits and Geo-Climatic Factors
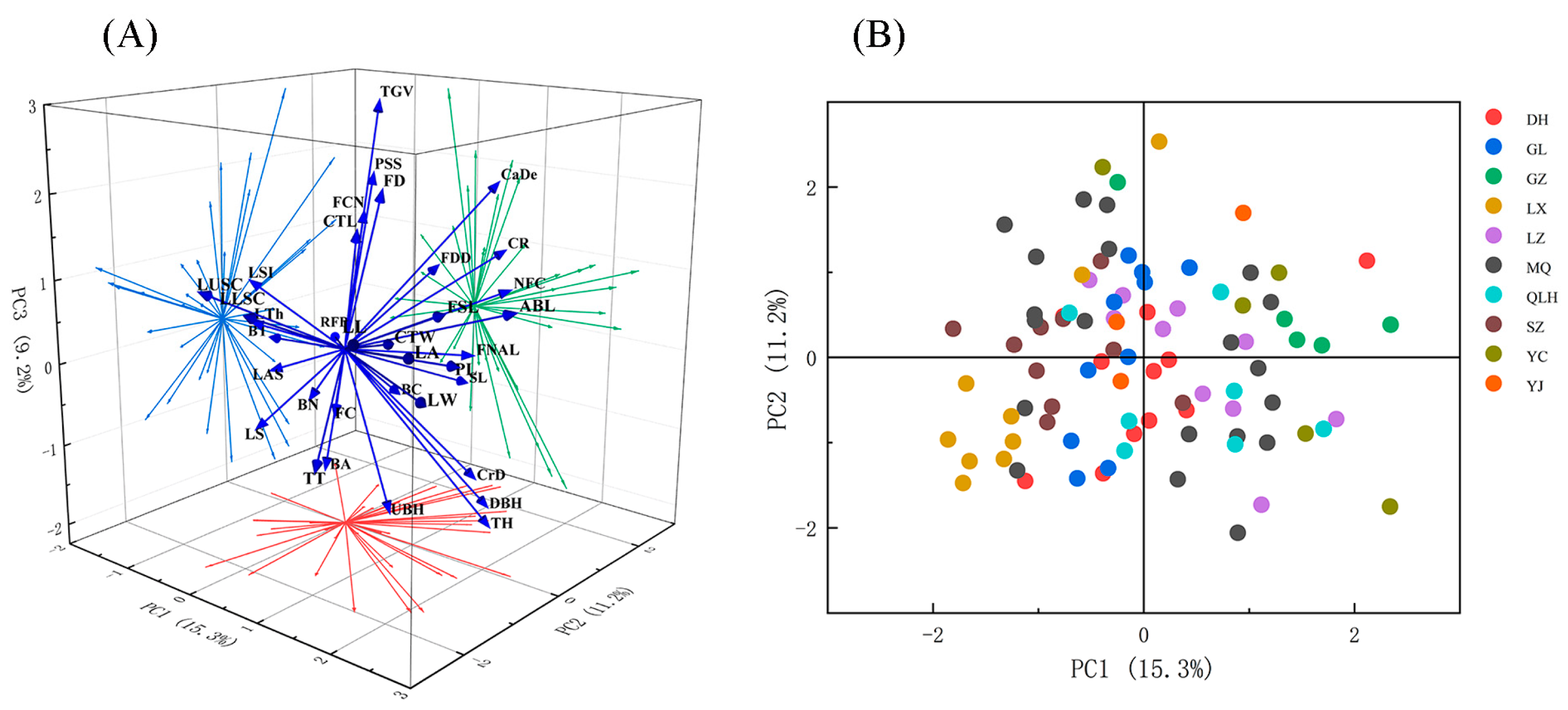
3.6. Principal Component Analysis of Phenotypic Traits
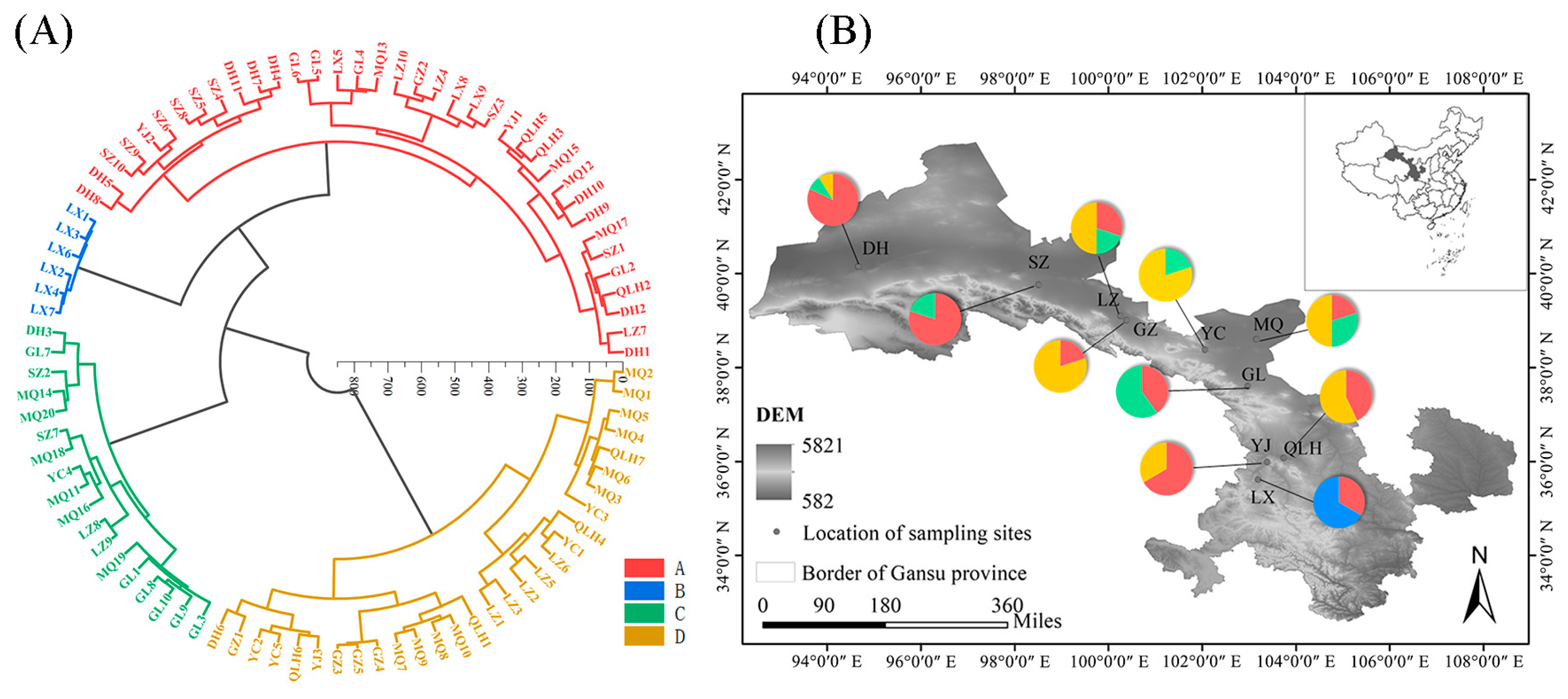
3.7. Cluster Analysis of E. angustifolia Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raj, D.; Govindaraju, D.; Orians, C. Genetic variation among pitch pine Pinus rigida families from Walden Woods: Heritability and path analysis of developmental variation of phenotypic traits. Rhodora 2009, 108, 356–369. [Google Scholar] [CrossRef]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef]
- Donohue, K.; Pyle, E.H.; Messiqua, D.; Heschel, M.S.; Schmitt, J. Adaptive divergence in plasticity in natural populations of Impatiens capensis and its consequences for performance in novel habitats. Evolution 2001, 55, 692–702. [Google Scholar] [CrossRef]
- Shirley, L.; Vesk, P. Eucalyptus phylogenetics: How traits, evolution, and the environment influence the distribution of a diverse genus. In Proceedings of the 94th ESA Annual Convention 2009, Albuquerque, NM, USA, 2–7 August 2009. [Google Scholar]
- Etterson, J.R. Evolutionary potential of Chamaecrista asciculata in relation to climate change. 1. Clinal patterns of selection along an environmental gradient in the Great Plains. Evolution 2004, 58, 1446–1456. [Google Scholar] [CrossRef]
- Han, H.; Li, S.; Gan, X.; Zhang, X. Phenotypic diversity in natural populations of an endangered plant Tetracentron Sinense. Bot. Sci. 2017, 95, 283–294. [Google Scholar] [CrossRef]
- Iezzoni, A.F.; Pritts, M.P. Applications of principal component analysis to horticultural research. HortScience 1991, 26, 334–338. [Google Scholar] [CrossRef]
- Omire, A.; Budambula, N.L.M.; Neondo, J.; Gituru, R.; Mweu, C. Phenotypic diversity of doum palm (Hyphaene compressa), a semi-domesticated palm in the arid and semi-arid regions of Kenya. Scientifica 2020, 2020, 4920830. [Google Scholar] [CrossRef]
- Asheghi, M.S. Phenotypic diversity of Crataegus orientalis subsp. szovitsii (Pojark.) K.I.Chr. population in Markazi Province, Iran. Erwerbs-Obstbau 2022, 64, 69–74. [Google Scholar] [CrossRef]
- Xin, Y.-H.; Wu, Y.-X.; Qiao, B.; Su, L.; Xie, S.-Q.; Ling, P. Evaluation on the phenotypic diversity of Calamansi (Citrus microcarpa) germplasm in Hainan island. Sci. Rep. 2022, 12, 371. [Google Scholar] [CrossRef]
- Yue, D.X.; Li, Z.Z.; Hui, C. Development trend of Gansu’s ecological footprint and ecological capacity. Acta Bot. Bor-Occid. Sin. 2004, 24, 454–463. [Google Scholar] [CrossRef]
- Li, Y.; Li, H. Gansu biodiversity and ecological protection. For. Gansu 2004, 04, 18–20. [Google Scholar]
- Li, X.H. Discussion on biodiversity protection in Gansu. For. China 2012, 759, 31. [Google Scholar]
- Tian, Y. Study on Fruit Phenotypic Diversity of Natural Population in Calligonum mongolicum in Gansu Province. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2013. [Google Scholar]
- Luo, H.J.; Li, J.H.; Zhao, S.C.; Wang, L.G. Phenotypic characteristics and diversity of walnut nuts in Gansu Province. China Fruits 2019, 199, 87–92. [Google Scholar] [CrossRef]
- Stannard, M.; Ogle, D.; Holzworth, L.; Scianna, J.; Sunleaf, E. History, Biology, Ecology, Suppression and Revegetation of Russian-olive Sites (Elaeagnus angustifolia L.); Plant Materials Technical Note; USDA-Natural Resources Conservation Servic: Boise, ID, USA, 2002. [Google Scholar]
- Yu, W.W.; Yan, G.R. Resources and research situation of Elaeagnus spp. J. Tianjin Agric. Univ. 2009, 16, 46–50. [Google Scholar]
- Kiseleva, T.I.; Chindyaeva, L.N. Biology of oleaster (Elaeagnus angustifolia L.) at the northeastern limit of its range. Contemp. Probl. Ecol. 2011, 4, 218–222. [Google Scholar] [CrossRef]
- Yaghoubi, M.; Alirezalu, K.; Hesari, J.; Peighambardoust, S.H.; Marcinkowska-Lesiak, M.; Barzegar, Y.; Hoseinian-Khosrowshahi, S.R.; Marszałek, K.; Mousavi Khaneghah, A. Application of oleaster leaves (Elaeagnus angustifolia L.) essential oil and natural nanoparticle preservatives in frankfurter-type sausages: An assessment of quality attributes and stability during refrigerated storage. Meat Sci. 2023, 198, 109097. [Google Scholar] [CrossRef]
- Çakmakçı, S.; Topdaş, E.F.; Kalın, P.; Han, H.; Şekerci, P.; Köse, L.P.; Gülçin, İ. Antioxidant capacity and functionality of oleaster (Elaeagnus angustifolia L.) flour and crust in a new kind of fruity ice cream. Int. J. Food Sci. Technol. 2015, 50, 472–481. [Google Scholar] [CrossRef]
- Ahmadiani, A.; Hosseiny, J.; Semnanian, S.; Javan, M.; Saeedi, F.; Kamalinejad, M.; Saremi, S. Antinociceptive and anti-inflammatory effects of Elaeagnus angustifolia fruit extract. J. Ethnopharmacol. 2000, 72, 287–292. [Google Scholar] [CrossRef]
- Chen, C.X.; Xie, X.H.; Wang, Y.P.; Li, J.P.; Xin, J.; Zhu, T.T.; Liu, J.; Chen, M. Effects of salt and drought on the physiological characteristics of Elaeagnus angustifolia L. seedlings. Acta Ecol. Sin. 2019, 39, 4540–4550. [Google Scholar]
- Safdari, L.; Khadivi, A. Identification of the promising oleaster (Elaeagnus angustifolia L.) genotypes based on fruit quality-related characters. Food Sci. Nutr. 2021, 9, 5712–5721. [Google Scholar] [CrossRef]
- Zeng, Z.H.; Li, Q.W.; Ti, Y.J. Analysis on phenotypic diversity and ornamental of wild Elaeagnus angustifolia from Zhangye City of Gansu Province. J. Plant Resour. Environ. 2021, 30, 12–21. [Google Scholar] [CrossRef]
- Xu, J.; Yu, W.Z.; Ni, W.; Mao, Y.F.; Liu, Q.; Shen, X. Diversity of phenotypic traits and fruit quality of Elaeagnus germplasm resources. Northern Hortic. 2016, 373, 20–24. [Google Scholar] [CrossRef]
- Fan, Y.C.; Chai, S.S.; Zhang, M.M.; Zhao, X.H.; Shen, X. Phenotypic genetic diversity of Elaeagnus angustifolia resources from Ningxia. Northern Hortic. 2018, 422, 37–43. [Google Scholar]
- Wang, C.; Gong, H.; Feng, M.; Tian, C. Phenotypic variation in leaf, fruit and seed traits in natural populations of Eucommia Ulmoides, a relict Chinese endemic tree. Forests 2023, 14, 462. [Google Scholar] [CrossRef]
- Brouwers, N.C. Biostatistical design and analysis using R: A practical guide. Austral Ecol. 2012, 37, e19. [Google Scholar] [CrossRef]
- Ge, S.; Wang, M.X.; Chen, Y.W. An analysis of population genetic structure of Masson pine by isozyme technique. Sci. Silv. Sin. 1988, 04, 399–409. [Google Scholar]
- Sokal, R.; Rohlf, F. Biometry: The Principles and Practice of Statistics in Biological Research. Available online: https://www.researchgate.net/publication/44554870_Biometry_the_principles_and_practice_of_statistics_in_biological_research_Robert_R_Sokal_and_F_James_Rohlf (accessed on 12 August 2022). [CrossRef]
- Maughan, P.J.; Saghi Maroof, M.A.; Buss, G.R. Microsatellite and amplified sequence length polymorphisms in cultivated and wild soybean. Genome 1995, 38, 715–723. [Google Scholar] [CrossRef]
- Jain, S.; Qualset, C.; Bhatt, G.; Wu, K. Geographical patterns of phenotypic diversity in a world collection of durum wheats. Crop Sci. 1975, 15, 700–704. [Google Scholar] [CrossRef]
- Zhao, Y.; Bi, Q.X.; Ju, J.; Yu, D.; Fan, S.Q.; Chen, M.Y.; Cui, Y.F.; Fu, G.H.; Wang, L.B. Geographic variation of seed and seedling growing traits in Xanthoceras sorbifolium. For. Res. 2019, 32, 160–168. [Google Scholar] [CrossRef]
- Tu, Z.D.; Chang, Z.F. Investigation of Elaeagnus angustifolia variety resources in Gansu Province. J. Gansu For. Sci. Tech. 1993, 04, 20–23. [Google Scholar]
- Qin, Z.K.; Liu, N.; Zhou, X.; Li, Z.H.; Cao, F.L.; Li, H. Phenotypic diversity of Quercus gilva natural populations inmiddle subtropical China. Guihaia 1–20.
- Ramírez-Valiente, J.A.; Solé-Medina, A.; Pyhäjärvi, T.; Savolainen, O.; Cervantes, S.; Kesälahti, R.; Kujala, S.T.; Kumpula, T.; Heer, K.; Opgenoorth, L.; et al. Selection patterns on early-life phenotypic traits in Pinus Sylvestris are associated with precipitation and temperature along a climatic gradient in Europe. New Phytol. 2021, 229, 3009–3025. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, P.; Qi, Q.; Zhang, Q.; Gao, X.; Lin, M.; Cui, Y. Phenotypic diversity and variation of Loniceracaerulea populations in the Changbai Mountain alongside the elevation gradient. Pol. J. Environ. Stud. 2020, 30, 705–716. [Google Scholar] [CrossRef]
- Liang, S.; Rong, X.; Sai, L.; Chen, J.; Changqing, X.; Caixiang, X.; Tongning, L. Phenotypic variation of seed Traits of Haloxylon ammodendron and its affecting factors. Biochem. Syst. Ecol. 2015, 60, 81–87. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, C.; Li, J.; Miao, Y.; Cui, K. Phenotypic diversity analysis and superior family selection of industrial raw material forest species-Pinus Yunnanensis Franch. Forests 2022, 13, 618. [Google Scholar] [CrossRef]
- Diao, S.F.; Shao, W.H.; Jiang, J.M.; Dong, R.X.; Sun, H.G. Phenotypic diversity in natural populations of Sapindus mukorossi based on fruit and seed traits. Acta Ecol. Sin. 2014, 34, 1451–1460. [Google Scholar] [CrossRef]
- Kidner, C.; Umbreen, S. Why Is Leaf Shape so Variable? Int. J. Plant Dev. Biol. 2010, 4, 64–75. [Google Scholar]
- Vidaković, A.; Liber, Z.; Šatović, Z.; Idžojtić, M.; Volenec, I.; Zegnal, I.; Pintar, V.; Radunić, M.; Poljak, I. Phenotypic diversity of almond-leaved pear (Pyrus Spinosa Forssk.) along Eastern Adriatic Coast. Forests 2021, 12, 1630. [Google Scholar] [CrossRef]
- Wu, S.H. Elaeagnus angustifolia, an excellent tree species for windbreak and sand fixation in Northwest China. Bull. Biol. 1964, 6, 10–12. [Google Scholar]
- Gu, X.J.; Wang, Y.K. Problems and prospects in the development of Elaeagnus angustifolia resources in Minqin County. Jiangxi Agric. 2017, 111, 66. [Google Scholar] [CrossRef]
- Zhang, Z.Y. Analysis on the performance and sustainable developmentof Elaeagnus angustifolia in Gaotai County. Agric. Technol. Equip. 2022, 393, 79–81. [Google Scholar]
- Ma, L.Q.; Yang, F.Q.; Xu, L. Fractal feature of Elaeagnus angustifolia tree of in Minqin desert. J. Gansu Forestry Sci. Tech. 2016, 41, 27–35. [Google Scholar] [CrossRef]
- Petruccelli, R.; Ganino, T.; Ciaccheri, L.; Maselli, F.; Mariotti, P. Phenotypic diversity of traditional cherry accessions present in the Tuscan region. Sci. Hortic. 2013, 150, 334–347. [Google Scholar] [CrossRef]
- Wu, G.S.; Sun, L.D.; Hao, R.J.; Shi, W.F.; Zhang, J.; Chen, J.X. Study on the phenotypic diversity of P. mume Sieb.et Zucc. germ plasm resources. J. Anhui Agric. Sci. 2011, 39, 12008–12009+12012. [Google Scholar] [CrossRef]
- Yu, W.W.; Wang, X.L.; Li, H.; Yan, G.R. Comparative studies on botanical characters of Elaeagnus moorcroftii Wall. ex Schlecht. and Elaeagnus oxycarpa Schlecht. J. Tianjin Agric. Univ. 2012, 19, 36–38+50. [Google Scholar] [CrossRef]
- Wang, P.P. The Variation Pattern of Elaeagnus L. in Xinjiang Based on Morphological Characters. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2015. [Google Scholar]
- Li, W.; Wang, P.; Qiqige; Zhang, Q.C.; Huang, B.J.; Xiao, Z.J. Phenotypic diversity analysis of blueberry germplasm resources. J. Beijing For. Univ. 2020, 42, 124–134. [Google Scholar] [CrossRef]
- Szamosi, C.; Solmaz, I.; Sari, N.; Bársony, C. Morphological evaluation and comparison of Hungarian and Turkish melon (Cucumis melo L.) Germplasm. Sci. Hortic. 2010, 124, 170–182. [Google Scholar] [CrossRef]
- He, T.; Fan, X.L.; Lu, L.; Huang, T.F. Phenotypic trait diversity of Dendrobium nobile from different provenances and their correlations with geographical factors. J. Plant Resour. Environ. 2021, 30, 1–11. [Google Scholar] [CrossRef]
- Fu, Q.J.; Li, C.N.; Chen, Y.; Zhao, F.K.; Sun, Y. Analysis of Phenotypic diversity of major Salvia splendens germplasm resources in China. J. Plant Genet. Resour. 2015, 16, 294–299. [Google Scholar] [CrossRef]
- Rui, W.J.; Wang, X.M.; Zhang, Q.N.; Hu, X.Y.; Hu, X.H.; Fu, J.J.; Gao, Y.M.; Li, J.S. Genetic diversity analysis of 353 tomato germplasm resources by phenotypic traits. Acta Hortic. Sin. 2018, 45, 561–570. [Google Scholar] [CrossRef]
- Khadivi, A. Phenotypic characterization of Elaeagnus angustifolia using multivariate analysis. Ind. Crops Prod. 2018, 120, 155–161. [Google Scholar] [CrossRef]
- Norman, P.; Tongoona, P.; Shanahan, P. Determination of tnterrelationships among agr-morphological traits of yams (Discorea spp.) using correrlation and factor analyses. J. Appl. Biosci. 2011, 45, 3059–3070. [Google Scholar]
- Zeng, Y.-F.; Zhang, J.-G.; Abuduhamiti, B.; Wang, W.-T.; Jia, Z.-Q. Phylogeographic patterns of the desert poplar in Northwest China shaped by both geology and climatic oscillations. BMC Evol. Biol. 2018, 18, 75. [Google Scholar] [CrossRef]
- Miller, R.B. Hawkmoths and the geographic patterns of floral variation in Aquilegia caerulea. Evolution 1981, 35, 763–774. [Google Scholar] [CrossRef]
- Steiner, K.E.; Whitehead, V.B.; Johnson, S.D. Floral and pollinator divergence in two sexually deceptive south African orchids. Am. J. Bot. 1994, 81, 185–194. [Google Scholar] [CrossRef]
- Johnson, S.D.; Steiner, K.E. Long-tongued fly pollination and evolution of floral spur length in the Disa draconis complex (Orchidaceae). Evolution 1997, 51, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Galen, C. Why do flowers vary? BioScience 1999, 49, 631–640. [Google Scholar] [CrossRef]
- Galen, C.; Ritland, A.E.K. High and Dry: High and dry: Drought stress, sex-allocation trade-offs, and selection on flower size in the alpine wildflower Polemonium viscosum (Polemoniaceae). Am. Nat. 2000, 156, 72–83. [Google Scholar] [CrossRef]
- Carroll, A.B.; Pallardy, S.G.; Galen, C. Drought stress, plant water status, and floral trait expression in fireweed, Epilobium Angustifolium (Onagraceae). Am. J. Bot. 2001, 88, 438–446. [Google Scholar] [CrossRef]
- Herrera, J. Flower size variation in Rosmarinus officinalis: Individuals, populations and habitats. Ann. Bot. 2004, 95, 431–437. [Google Scholar] [CrossRef]
- Cruz-Maldonado, N.; Weemstra, M.; Jiménez, L.; Roumet, C.; Angeles, G.; Barois, I.; de los Santos, M.; Morales-Martinez, M.A.; Palestina, R.A.; Rey, H.; et al. Aboveground-trait variations in 11 (sub)alpine plants along a 1000-m elevation gradient in tropical Mexico. Alp. Bot. 2021, 131, 187–200. [Google Scholar] [CrossRef]
- Huang, W.; Zhong, Y.; Zhang, C.; Ren, M.; Du, Y.; Song, X. Leaf traits and water-use characteristics of Impatiens hainanensis, a limestone-endemic plant under different altitudes in dry and foggy seasons. Water 2022, 14, 139. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, L.; Qi, D. Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes. Ecol. Evol. 2020, 10, 8166–8175. [Google Scholar] [CrossRef]
- Karadeniz, T. Selection of native “Cornelian” cherries grown in Turkey. J. Am. Pomol. Soc. 2002, 56, 164. [Google Scholar] [CrossRef]
- Du, Q.; Xu, B.; Gong, C.; Yang, X.; Pan, W.; Tian, J.; Li, B.; Zhang, D. Variation in growth, leaf, and wood property traits of Chinese white poplar (Populus tomentosa), a major industrial tree species in Northern China. Can. J. For. Res. 2014, 44, 326–339. [Google Scholar] [CrossRef]
- Cao, D.M.; Zhang, Y.H.; Cheng, X.Q.; Xiang, X.D.; Zhang, L.; Hu, J.J. Genetic variation of leaf phenotypic traits in different populations of Populus cathayana. Sci. Silvae Sin. 2021, 57, 56–67. [Google Scholar] [CrossRef]
- Fu, G.; Dai, X.; Symanzik, J.; Bushman, S. Quantitative gene–gene and gene–environment mapping for leaf shape variation using tree-based models. New Phytol. 2017, 213, 455–469. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Sample Size | Altitude (m, AL) | Longitude (°E, E) | Latitude (°N, N) | Annual Mean Temperature (°C, AMT) | Annual Precipitation (mm, AP) | Annual Mean Relative Humidity (%, AMRH) |
|---|---|---|---|---|---|---|---|
| Dunhuang (DH) | 11 | 1168.2 | 94.66 | 40.14 | 9.9 | 42.2 | 40 |
| Suzhou (SZ) | 10 | 1492.0 | 98.51 | 39.75 | 7.8 | 88.4 | 48 |
| Linze (LZ) | 10 | 1453.1 | 100.26 | 39.09 | 8.3 | 113.4 | 49 |
| Ganzhou (GZ)) | 5 | 1515.3 | 100.38 | 39.00 | 7.8 | 132.6 | 52 |
| Yongchang (YC) | 5 | 1728.7 | 102.06 | 38.37 | 5.4 | 211.8 | 52 |
| Minqin (MQ) | 20 | 1481.7 | 103.15 | 38.59 | 8.8 | 113.2 | 44 |
| Gulang (GL) | 10 | 1792.4 | 102.97 | 37.60 | 5.7 | 352.3 | 51 |
| Qilihe (QLH) | 7 | 1544.5 | 103.74 | 36.07 | 10.5 | 360.0 | 60 |
| Yongjing (YJ) | 3 | 1967.3 | 103.39 | 35.98 | 9.7 | 273.7 | 59 |
| Linxia (LX) | 9 | 2025.1 | 103.19 | 35.61 | 7.3 | 501.3 | 67 |
| No. | Character | Abbreviation | Unit |
|---|---|---|---|
| 1 | Tree height | TH | m |
| 2 | Crown diameter | CrD | cm |
| 3 | Diameter at breast height | DBH | cm |
| 4 | Under-branch height | UBH | cm |
| 5 | Branching angle | BA | ° |
| 6 | Branching number | BN | – |
| 7 | Leaf length | LL | mm |
| 8 | Leaf width | LW | mm |
| 9 | Leaf thickness | LTh | mm |
| 10 | Petiole length | PL | mm |
| 11 | Leaf shape index | LSI | – |
| 12 | Leaf area | LA | mm2 |
| 13 | Annual branch length | ABL | mm |
| 14 | Number of flower clusters on annual branches | NFC | – |
| 15 | Ratio of flower cluster number to branch length | RFB | – |
| 16 | Flower diameter | FD | mm |
| 17 | Calyx tube length | CTL | mm |
| 18 | Calyx tube width | CTW | mm |
| 19 | Flower stalk length | FSL | mm |
| 20 | Flower color number | FCN | – |
| 21 | Floret number in axils of leaves | FNAL | – |
| No. | Character | Abbreviation | Grading Assignment | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| 1 | Trunk type | TT | Multi-trunk shrubs | Less trunk shrubs | Arbor | – | – | – | – |
| 2 | Crown roundness | CR | Messy | Moderate | Rounded | – | – | – | – |
| 3 | Canopy density | CaDe | Low | Medium | High | – | – | – | – |
| 4 | Tree growth vigor | TGV | Low | Medium | High | – | – | – | – |
| 5 | Branch thorn | BT | Present | Absent | – | – | – | – | – |
| 6 | Branch color | BC | Hay yellow | Brownish green | Red brown | Dark brown | – | – | – |
| 7 | Leaf shape | LS | Oval-shaped | Long oval | Ovate | Lanceolate | Narrow lanceolate | – | – |
| 8 | Leaf apex shape | LAS | Obtuse | Acuminate | – | – | – | – | – |
| 9 | Speckles on leaves | SL | Low | Medium | High | – | – | – | – |
| 10 | Leaf upper surface color | LUSC | Aloe gray(a) * | Aloe gray(b) | Aloe gray(c) | Aloe gray(d) | Dark olive green | Grass green(a) | Grass green(b) |
| 11 | Leaf lower surface color | LLSC | Aloe gray(a) | Aloe gray(b) | Aloe gray(c) | – | – | – | – |
| 12 | Petal spreading state | PSS | Explanate | Curling outward | – | – | – | – | – |
| 13 | Flower density degree | FDD | Low | Medium | High | – | – | – | – |
| 14 | Flower color | FC | Rice white | Beige | Pale yellow | Bright yellow | Orange | – | – |
| Traits | F Value | Proportion of Variance Components (%) | Phenotypic Differentiation Coefficients (%) | |||
|---|---|---|---|---|---|---|
| Among Populations | Within Populations | Among Populations | Within Populations | Random Errors | ||
| TH | 4.125 ** | 1.919 ** | 27.58 | 27.09 | 45.33 | 50.45 |
| CrD | 2.458 ** | 0.8 | 22.50 | 15.46 | 62.04 | 59.28 |
| DBH | 4.075 ** | 1.949 ** | 32.76 | 24.37 | 42.87 | 57.34 |
| UBH | 3.076 ** | 0.682 | 32.48 | 11.21 | 56.31 | 74.34 |
| BA | 0.996 | 1.217 | 12.11 | 23.02 | 64.86 | 34.47 |
| BN | 0.503 | 1.117 | 6.64 | 22.95 | 70.41 | 22.45 |
| LL | 3.319 ** | 1.751 | 24.06 | 26.80 | 49.14 | 47.31 |
| LW | 4.416 ** | 2.048 ** | 28.45 | 27.87 | 43.68 | 50.52 |
| LTh | 4.527 ** | 1.22 | 32.50 | 18.75 | 48.75 | 63.41 |
| PL | 2.992 ** | 1.16 | 24.49 | 20.04 | 55.47 | 54.99 |
| LSI | 2.793 ** | 1.432 | 22.17 | 24.01 | 53.82 | 48.01 |
| LA | 4.824 ** | 1.636 | 32.04 | 22.95 | 45.02 | 58.27 |
| ABL | 7.656 ** | 1.416 | 43.94 | 17.16 | 38.90 | 71.92 |
| NFC | 3.849 ** | 2.122 ** | 25.48 | 29.65 | 44.87 | 46.22 |
| RFB | 2.779 ** | 2.394 ** | 19.02 | 34.59 | 46.39 | 35.48 |
| FD | 4.658 ** | 0.963 | 34.59 | 15.09 | 50.32 | 69.62 |
| CTL | 2.574 ** | 0.776 | 23.42 | 14.91 | 61.67 | 61.10 |
| CTW | 17.57 ** | 4.657 ** | 51.41 | 28.76 | 19.83 | 64.12 |
| FSL | 5.911 ** | 1.254 | 38.55 | 17.26 | 44.20 | 69.07 |
| FCN | 2.031 | 0.595 | 20.18 | 12.47 | 67.35 | 61.80 |
| FNAL | 14.212 ** | 0.539 | 64.23 | 5.14 | 30.63 | 92.59 |
| TT | 2.05 ** | 1.591 | 16.82 | 27.56 | 55.62 | 37.90 |
| CR | 3.576 ** | 1.09 | 28.26 | 18.18 | 53.56 | 60.85 |
| CaDe | 2.447 ** | 1.222 | 20.73 | 21.86 | 57.41 | 48.68 |
| TGV | 3.334 ** | 0.982 | 27.36 | 17.02 | 55.62 | 61.65 |
| BT | 12.341 ** | 2.288 ** | 51.53 | 20.16 | 28.30 | 71.88 |
| BC | 2.169 ** | 0.962 | 19.76 | 18.50 | 61.75 | 51.64 |
| LS | 2.203 ** | 0.81 | 20.61 | 15.99 | 63.40 | 56.31 |
| LAS | 3.348 ** | 0.946 | 27.62 | 16.47 | 55.91 | 62.65 |
| SL | 1.909 | 0.938 | 17.89 | 18.56 | 63.54 | 49.09 |
| LUSC | 1.348 | 1.338 | 12.31 | 25.80 | 61.89 | 32.29 |
| LLSC | 1.816 | 1.48 | 15.50 | 26.66 | 57.85 | 36.76 |
| PSS | 11.82 ** | 2.826 ** | 48.12 | 24.29 | 27.59 | 66.45 |
| FDD | 1.348 | 2.36 ** | 10.28 | 38.01 | 51.71 | 21.29 |
| FC | 2.316 ** | 1.33 | 19.46 | 23.59 | 56.95 | 45.20 |
| Mean | 27.28 | 21.49 | 51.23 | 54.15 | ||
| Traits | Coefficient of Variation (CV) (%) | Diversity | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DH | SZ | LZ | GZ | YC | MQ | GL | QLH | YJ | LX | Total | ||
| TH | 37.92 | 28.24 | 27.65 | 10.47 | 16.57 | 46.47 | 16.52 | 33.06 | 36.66 | 43.43 | 37.12 | 1.9570 |
| CrD | 64.17 | 25.65 | 19.03 | 11.19 | 42.61 | 26.56 | 21.21 | 57.22 | 68.54 | 40.08 | 41.64 | 1.7890 |
| DBH | 60.76 | 21.28 | 26.64 | 15.09 | 25.92 | 58.15 | 70.13 | 95.17 | 75.98 | 76.90 | 60.87 | 1.9219 |
| UBH | 30.94 | 75.83 | 42.93 | 47.86 | 56.19 | 33.00 | 38.64 | 95.44 | 45.48 | 23.39 | 61.61 | 1.6802 |
| BA | 47.26 | 28.04 | 45.22 | 23.89 | 20.39 | 48.32 | 38.03 | 64.44 | 31.73 | 31.40 | 43.64 | 1.9106 |
| BN | 37.97 | 30.87 | 32.22 | 20.32 | 40.00 | 37.67 | 27.20 | 30.62 | 33.33 | 29.75 | 32.79 | 1.0927 |
| LL | 18.42 | 14.57 | 26.38 | 27.67 | 24.62 | 17.67 | 16.49 | 20.23 | 11.56 | 17.82 | 21.82 | 2.0415 |
| LW | 20.37 | 13.13 | 27.57 | 21.94 | 23.07 | 25.04 | 14.58 | 18.43 | 7.55 | 14.81 | 23.15 | 1.9679 |
| LTh | 14.64 | 12.35 | 19.63 | 17.03 | 9.25 | 19.81 | 16.80 | 14.48 | 5.26 | 3.54 | 17.93 | 2.0265 |
| PL | 23.42 | 22.38 | 23.97 | 24.64 | 24.77 | 20.67 | 18.45 | 16.62 | 7.67 | 10.40 | 23.06 | 1.9730 |
| LSI | 17.23 | 13.79 | 16.56 | 8.73 | 14.56 | 18.45 | 18.12 | 15.20 | 12.68 | 20.45 | 17.98 | 2.0085 |
| LA | 46.36 | 24.40 | 36.72 | 46.63 | 46.96 | 41.90 | 23.36 | 35.04 | 12.93 | 25.61 | 46.93 | 1.8967 |
| ABL | 26.11 | 40.24 | 25.52 | 23.31 | 20.75 | 21.86 | 13.83 | 23.23 | 18.26 | 30.32 | 31.65 | 2.0228 |
| NFC | 17.53 | 13.84 | 20.18 | 5.27 | 10.75 | 28.46 | 25.39 | 24.38 | 22.05 | 46.43 | 25.91 | 1.9858 |
| RFB | 24.17 | 25.19 | 34.13 | 30.23 | 20.08 | 27.03 | 16.87 | 15.80 | 15.78 | 24.87 | 26.66 | 2.0117 |
| FD | 13.16 | 7.47 | 9.09 | 7.33 | 18.57 | 8.86 | 8.06 | 5.27 | 9.12 | 4.68 | 11.05 | 2.0723 |
| CTL | 11.76 | 14.49 | 13.21 | 12.63 | 7.26 | 10.53 | 10.23 | 9.95 | 13.28 | 5.63 | 12.04 | 2.0556 |
| CTW | 13.10 | 10.03 | 10.18 | 10.83 | 14.85 | 23.55 | 9.63 | 13.35 | 16.75 | 9.73 | 19.71 | 1.9836 |
| FSL | 27.96 | 14.04 | 37.46 | 19.87 | 44.76 | 29.83 | 14.63 | 20.19 | 26.03 | 28.64 | 35.55 | 1.9125 |
| FCN | 37.40 | 47.13 | 34.00 | 24.83 | 24.83 | 31.44 | 28.41 | 34.08 | 34.55 | 35.35 | 36.52 | 0.9593 |
| FNAL | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 59.88 | 17.49 | 0.2449 |
| Mean | 28.13 | 23.00 | 25.16 | 19.51 | 24.13 | 27.39 | 21.27 | 30.58 | 24.06 | 27.77 | 30.72 | 1.7864 |
| Traits | Distribution Frequency of Each Grade | Diversity | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| TT | 11.1 | 8.9 | 80.0 | – | – | – | – | 0.6378 |
| CR | 55.6 | 23.3 | 21.1 | – | – | – | – | 0.9945 |
| CaDe | 33.3 | 36.7 | 30.0 | – | – | – | – | 1.0953 |
| TGV | 8.9 | 30.0 | 61.1 | – | – | – | – | 0.8773 |
| BT | 65.6 | 34.4 | – | – | – | – | – | 0.6439 |
| BC | 5.6 | 32.2 | 20.0 | 42.2 | – | – | – | 1.2114 |
| LS | 26.7 | 6.7 | 12.2 | 48.9 | 5.6 | – | – | 1.3003 |
| LAS | 37.8 | 62.2 | – | – | – | – | – | 0.6630 |
| SL | 26.7 | 41.1 | 32.2 | – | – | – | – | 1.0828 |
| LUSC | 4.4 | 24.4 | 17.8 | 5.6 | 16.7 | 21.1 | 10.0 | 1.8076 |
| LLSC | 43.3 | 44.4 | 12.2 | – | – | – | – | 0.9797 |
| PSS | 77.8 | 22.2 | – | – | – | – | – | 0.5297 |
| FDD | 20.0 | 28.9 | 51.1 | – | – | – | – | 1.0236 |
| FC | 11.1 | 27.8 | 24.4 | 16.7 | 20.0 | – | – | 1.5648 |
| Mean | 1.0294 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, R.; Zhu, Z.; Shi, N.; Li, Y.; Dang, J.; Wang, Y.; Ma, Y.; Xu, X.; Liu, T. Phenotypic Diversity Analysis in Elaeagnus angustifolia Populations in Gansu Province, China. Forests 2023, 14, 1143. https://doi.org/10.3390/f14061143
Shi R, Zhu Z, Shi N, Li Y, Dang J, Wang Y, Ma Y, Xu X, Liu T. Phenotypic Diversity Analysis in Elaeagnus angustifolia Populations in Gansu Province, China. Forests. 2023; 14(6):1143. https://doi.org/10.3390/f14061143
Chicago/Turabian StyleShi, Rongrong, Zhu Zhu, Ningrui Shi, Yongmei Li, Jun Dang, Yanli Wang, Yonglong Ma, Xiangyun Xu, and Ting Liu. 2023. "Phenotypic Diversity Analysis in Elaeagnus angustifolia Populations in Gansu Province, China" Forests 14, no. 6: 1143. https://doi.org/10.3390/f14061143