
Application of MaxEnt Model in Biomass Estimation: An Example of Spruce Forest in the Tianshan Mountains of the Central-Western Part of Xinjiang, China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Calculation of AGB on the Sample Field
2.3. Processing and Selecting Environment Variables
2.4. Modeling of AGB Estimation
3. Results
3.1. Evaluation of Model Accuracy
3.2. Analysis of Environmental Variables Affecting the Distribution of AGB
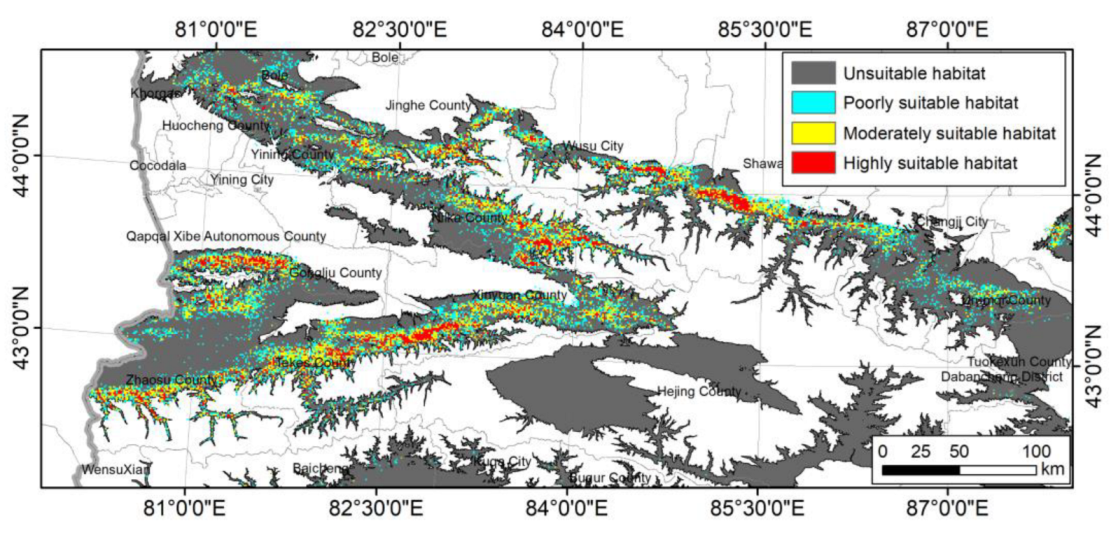
3.3. Spatial Distribution Pattern of AGB
3.4. Accuracy and Uncertainty Analysis of AGB Estimation
4. Discussion
4.1. Dominant Factors Influencing AGB Distribution
4.2. Accuracy and Applicability of AGB Estimation Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bellard, C.; Marino, C.; Courchamp, F. Ranking threats to biodiversity and why it doesn’t matter. Nat. Commun. 2022, 13, 2616. [Google Scholar] [CrossRef] [PubMed]
- Balik, J.A.; Greig, H.S.; Taylor, B.W.; Wissinger, S.A. Consequences of climate-induced range expansions on multiple ecosystem functions. Commun. Biol. 2023, 6, 390. [Google Scholar] [CrossRef]
- Robbins Schug, G.; Buikstra, J.E.; DeWitte, S.N.; Baker, B.J.; Berger, E.; Buzon, M.R.; Davies-Barrett, A.M.; Goldstein, L.; Grauer, A.L.; Gregoricka, L.A.; et al. Climate change, human health, and resilience in the Holocene. Proc. Natl. Acad. Sci. USA 2023, 120, e2209472120. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Global Forest Resources Assessment 2020; FAO: Rome, Italy, 2020.
- Watson, R.T.; Noble, I.R.; Bolin, B.; Ravindranath, N.; Verardo, D.J.; Dokken, D.J. Land Use, Land-Use Change and Forestry: A Special Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lu, N.; Fu, B.; Wang, S.; Deng, L.; Wang, L. Current and future carbon stocks of natural forests in China. For. Ecol. Manag. 2022, 511, 120137. [Google Scholar] [CrossRef]
- Devillers, R.; Desjardin, E.; De Runz, C. Imperfection of Geographic Information: Concepts and Terminologies. Geogr. Data Imperfection 1 Theory Appl. 2019, 2, 11–24. [Google Scholar] [CrossRef]
- Koven, C.D.; Chambers, J.Q.; Georgiou, K.; Knox, R.; Negron-Juarez, R.; Riley, W.J.; Arora, V.K.; Brovkin, V.; Friedlingstein, P.; Jones, C.D. Controls on terrestrial carbon feedbacks by productivity versus turnover in the CMIP5 Earth System Models. Biogeosciences 2015, 12, 5211–5228. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment Update 2005 (FRA 2005): Terms and Definitions; FAO: Rome, Italy, 2004. [Google Scholar]
- Walker, W.S.; Gorelik, S.R.; Cook-Patton, S.C.; Baccini, A.; Farina, M.K.; Solvik, K.K.; Ellis, P.W.; Sanderman, J.; Houghton, R.A.; Leavitt, S.M.; et al. The global potential for increased storage of carbon on land. Proc. Natl. Acad. Sci. USA 2022, 119, e2111312119. [Google Scholar] [CrossRef]
- Santoro, M.; Cartus, O.; Carvalhais, N.; Rozendaal, D.M.A.; Avitabile, V.; Araza, A.; de Bruin, S.; Herold, M.; Quegan, S.; Rodríguez-Veiga, P.; et al. The global forest above-ground biomass pool for 2010 estimated from high-resolution satellite observations. Earth Syst. Sci. Data 2021, 13, 3927–3950. [Google Scholar] [CrossRef]
- Hynynen, J.; Salminen, H.; Ahtikoski, A.; Huuskonen, S.; Ojansuu, R.; Siipilehto, J.; Lehtonen, M.; Eerikäinen, K. Long-term impacts of forest management on biomass supply and forest resource development: A scenario analysis for Finland. Eur. J. For. Res. 2015, 134, 415–431. [Google Scholar] [CrossRef]
- Dias, A.M.; Machado, J.S.; Dias, A.M.P.G.; Silvestre, J.D.; de Brito, J. Influence of the Wood Species, Forest Management Practice and Allocation Method on the Environmental Impacts of Roundwood and Biomass. Forests 2022, 13, 1357. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Houghton, R.A.; Dos Santos AlvalÁ, R.C.; Soares, J.V.; Yu, Y. Distribution of aboveground live biomass in the Amazon basin. Glob. Chang. Biol. 2007, 13, 816–837. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, J.; Liang, S.; Li, X.; Li, M. An Evaluation of Eight Machine Learning Regression Algorithms for Forest Aboveground Biomass Estimation from Multiple Satellite Data Products. Remote Sens. 2020, 12, 4015. [Google Scholar] [CrossRef]
- Kushwaha, S.P.S.; Nandy, S. Forest Biomass Assessment Integrating Field Inventory and Optical Remote Sensing Data: A Systematic Review. Int. J. Plant Environ. 2021, 7, 181–186. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Buebos-Esteve, D.E.; Mamasig, G.D.N.S.; Ringor, A.M.D.; Layog, H.N.B.; Murillo, L.C.S.; Dagamac, N.H.A. Modeling the potential distribution of two immortality flora in the Philippines: Applying MaxEnt and GARP algorithms under different climate change scenarios. Model. Earth Syst. Environ. 2023, 7, 1–7. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Alegria, C.; Almeida, A.M.; Roque, N.; Fernandez, P.; Ribeiro, M.M. Species Distribution Modelling under Climate Change Scenarios for Maritime Pine (Pinus pinaster Aiton) in Portugal. Forests 2023, 14, 591. [Google Scholar] [CrossRef]
- Ahmadi, M.; Hemami, M.-R.; Kaboli, M.; Shabani, F. MaxEnt brings comparable results when the input data are being completed; Model parameterization of four species distribution models. Ecol. Evol. 2023, 13, e9827. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef]
- Rodríguez-Veiga, P.; Saatchi, S.; Tansey, K.; Balzter, H. Magnitude, spatial distribution and uncertainty of forest biomass stocks in Mexico. Remote Sens. Environ. 2016, 183, 265–281. [Google Scholar] [CrossRef]
- Xu, L.; Saatchi, S.S.; Shapiro, A.; Meyer, V.; Ferraz, A.; Yang, Y.; Bastin, J.F.; Banks, N.; Boeckx, P.; Verbeeck, H.; et al. Spatial Distribution of Carbon Stored in Forests of the Democratic Republic of Congo. Sci. Rep. 2017, 7, 15030. [Google Scholar] [CrossRef]
- Harris, N.L.; Hagen, S.C.; Saatchi, S.S.; Pearson, T.R.H.; Woodall, C.W.; Domke, G.M.; Braswell, B.H.; Walters, B.F.; Brown, S.; Salas, W.; et al. Attribution of net carbon change by disturbance type across forest lands of the conterminous United States. Carbon Balance Manag. 2016, 11, 24. [Google Scholar] [CrossRef]
- Yu, Y.; Saatchi, S.; Domke, G.M.; Walters, B.; Woodall, C.; Ganguly, S.; Li, S.; Kalia, S.; Park, T.; Nemani, R.; et al. Making the US national forest inventory spatially contiguous and temporally consistent. Environ. Res. Lett. 2022, 17, 065002. [Google Scholar] [CrossRef]
- Ferreira, I.J.M.; Campanharo, W.A.; Fonseca, M.G.; Escada, M.I.S.; Nascimento, M.T.; Villela, D.M.; Brancalion, P.; Magnago, L.F.S.; Anderson, L.O.; Nagy, L.; et al. Potential aboveground biomass increase in Brazilian Atlantic Forest fragments with climate change. Glob. Chang. Biol. 2023, 29, 3098–3113. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Jones, S.D.; Soto-Berelov, M.; Haywood, A.; Hislop, S. Monitoring aboveground forest biomass dynamics over three decades using Landsat time-series and single-date inventory data. Int. J. Appl. Earth Obs. Geoinf. 2020, 84, 101952. [Google Scholar] [CrossRef]
- Puliti, S.; Hauglin, M.; Breidenbach, J.; Montesano, P.; Neigh, C.S.R.; Rahlf, J.; Solberg, S.; Klingenberg, T.F.; Astrup, R. Modelling above-ground biomass stock over Norway using national forest inventory data with ArcticDEM and Sentinel-2 data. Remote Sens. Environ. 2020, 236, 111501. [Google Scholar] [CrossRef]
- Campbell, M.J.; Dennison, P.E.; Kerr, K.L.; Brewer, S.C.; Anderegg, W.R.L. Scaled biomass estimation in woodland ecosystems: Testing the individual and combined capacities of satellite multispectral and lidar data. Remote Sens. Environ. 2021, 262, 112511. [Google Scholar] [CrossRef]
- Ahmad, N.; Ullah, S.; Zhao, N.; Mumtaz, F.; Ali, A.; Ali, A.; Tariq, A.; Kareem, M.; Imran, A.B.; Khan, I.A.; et al. Comparative Analysis of Remote Sensing and Geo-Statistical Techniques to Quantify Forest Biomass. Forests 2023, 14, 379. [Google Scholar] [CrossRef]
- Nesha, K.; Herold, M.; De Sy, V.; de Bruin, S.; Araza, A.; Malaga, N.; Gamarra, J.G.P.; Hergoualc’h, K.; Pekkarinen, A.; Ramirez, C.; et al. Exploring characteristics of national forest inventories for integration with global space-based forest biomass data. Sci. Total Environ. 2022, 850, 157788. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Forest aboveground biomass estimation using Landsat 8 and Sentinel-1A data with machine learning algorithms. Sci. Rep. 2020, 10, 9952. [Google Scholar] [CrossRef]
- Purohit, S.; Aggarwal, S.P.; Patel, N.R. Estimation of forest aboveground biomass using combination of Landsat 8 and Sentinel-1A data with random forest regression algorithm in Himalayan Foothills. Trop. Ecol. 2021, 62, 288–300. [Google Scholar] [CrossRef]
- Silveira, E.M.d.O.; Cunha, L.I.F.; Galvão, L.S.; Withey, K.D.; Acerbi Júnior, F.W.; Scolforo, J.R.S. Modelling aboveground biomass in forest remnants of the Brazilian Atlantic Forest using remote sensing, environmental and terrain-related data. Geocarto Int. 2019, 36, 281–298. [Google Scholar] [CrossRef]
- Xu, W.; Yang, L.; Chen, X.; GAO, Y.; Wang, L. Carbon storage, spatial distribution and the influence factors in Tianshan forests. Chin. J. Plant Ecol. 2016, 40, 364–373. [Google Scholar] [CrossRef]
- Adilai, S.; Chang, S.; Zhang, Y.; Sun, X.; Li, J.; LI, X. A decade variation of species composition and community structure of spruce forest in Tianshan Mountain. Chin. J. Ecol. 2021, 40, 3033–3040. [Google Scholar] [CrossRef]
- Zhu, H. Effects of Different Carbon Input Manipulations on Soil Carbon, Nitrogen and Biological Characteristics of Schrenk’s Spruce (Picea schenrenkiana) Forest. Ph.D. Thesis, Xinjiang University, Urumqi, China, 2021. [Google Scholar]
- Zhao, C.; Ding, Y.; Ye, B.; Zhao, Q. Spatial distribution of precipitation in Tianshan Mountains and its estimation. Adv. Water Sci. 2011, 22, 315–322. [Google Scholar] [CrossRef]
- Eli, A. Spatial Distribution of Tianshan Mountains Forests’s Soil Organic Carbon and Its Influencing Factors. Master’s Thesis, Xinjiang University, Urumqi, China, 2014. [Google Scholar]
- Sun, J. The Productivity of Forest Stand and the Distribution Regularity of Forest Types and in Tianshan Forest. Arid Zone Res. 1994, 11, 1–6. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the structure and allometry of plant vascular systems. Nature 1999, 400, 664–667. [Google Scholar] [CrossRef]
- Vorster, A.G.; Evangelista, P.H.; Stovall, A.E.L.; Ex, S. Variability and uncertainty in forest biomass estimates from the tree to landscape scale: The role of allometric equations. Carbon Balance Manag. 2020, 15, 8. [Google Scholar] [CrossRef]
- Liu, G. The Study on the Growth Rule of Picea schrenkiana var. Tianschanica and the Productivity of Communities in Tianshan. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2006. [Google Scholar]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A.M. Want to model a species niche? A step-by-step guideline on correlative ecological niche modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Zlateva, I.; Raykov, V.; Slabakova, V.; Stefanova, E.; Stefanova, K. Habitat suitability models of five keynote Bulgarian Black Sea fish species relative to specific abiotic and biotic factors. Oceanologia 2022, 64, 665–674. [Google Scholar] [CrossRef]
- Zhao, X.; Lei, M.; Wei, C.; Guo, X. Assessing the suitable regions and the key factors for three Cd-accumulating plants (Sedum alfredii, Phytolacca americana, and Hylotelephium spectabile) in China using MaxEnt model. Sci. Total Environ. 2022, 852, 158202. [Google Scholar] [CrossRef]
- Tao, Z. Predicting the changes in suitable habitats for six common woody species in Central Asia. Int. J. Biometeorol. 2023, 67, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; White, M.; Newell, G.; Pearson, R. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, B.; Wan, F.; Xiao, Q.; Dai, L. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodivers. Sci. 2007, 15, 365–372. [Google Scholar] [CrossRef]
- Khadanga, S.S.; Jayakumar, S. Tree biomass and carbon stock: Understanding the role of species richness, elevation, and disturbance. Trop. Ecol. 2020, 61, 128–141. [Google Scholar] [CrossRef]
- Taripanah, F.; Ranjbar, A. Quantitative analysis of spatial distribution of land surface temperature (LST) in relation Ecohydrological, terrain and socio- economic factors based on Landsat data in mountainous area. Adv. Space Res. 2021, 68, 3622–3640. [Google Scholar] [CrossRef]
- Li, X.; Gong, L.; Wei, B.; Ding, Z.; Zhu, H.; Li, Y.; Zhang, H.; Yonggang, M. Effects of climate change on potential distribution and niche differentiation of Picea schrenkiana in Xinjiang. Acta Ecol. Sin. 2022, 42, 4091–4100. [Google Scholar] [CrossRef]
- Qin, L.; Liu, K.; Shang, H.; Zhang, T.; Yu, S.; Zhang, R. Minimum temperature during the growing season limits the radial growth of timberline Schrenk spruce (P. schrenkiana). Agric. For. Meteorol. 2022, 322, 109004. [Google Scholar] [CrossRef]
- Shishov, V.V.; Arzac, A.; Popkova, M.I.; Yang, B.; He, M.; Vaganov, E.A. Experimental and theoretical analysis of tree-ring growth in cold climates. In Boreal Forests in the Face of Climate Change: Sustainable Management; Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 295–321. [Google Scholar]
- Reich, P.B.; Luo, Y.; Bradford, J.B.; Poorter, H.; Perry, C.H.; Oleksyn, J. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. Proc. Natl. Acad. Sci. USA 2014, 111, 13721–13726. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C. Responses of tree leaf gas exchange to elevated CO2 combined with changes in temperature and water availability: A global synthesis. Glob. Ecol. Biogeogr. 2021, 30, 2500–2512. [Google Scholar] [CrossRef]
- Peng, Z.; Zhang, Y.; Zhu, L.; Lu, Q.; Mo, Q.; Cai, J.; Guo, M. Divergent Tree Growth and the Response to Climate Warming and Humidification in the Tianshan Mountains, China. Forests 2022, 13, 886. [Google Scholar] [CrossRef]
- Wang, T.; Ren, H.; Ma, K. Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Trees 2005, 19, 736–742. [Google Scholar] [CrossRef]
- Zhang, R.; Yuan, Y.; Gou, X.; Zhang, T.; Zou, C.; Ji, C.; Fan, Z.; Qin, L.; Shang, H.; Li, X. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia 2016, 40, 36–42. [Google Scholar] [CrossRef]
- Wang, T.; Bao, A.; Xu, W.; Yu, R.; Zhang, Q.; Jiang, L.; Nzabarinda, V. Tree-ring-based assessments of drought variability during the past 400 years in the Tianshan mountains, arid Central Asia. Ecol. Indic. 2021, 126, 107702. [Google Scholar] [CrossRef]
- Bennett, A.C.; Penman, T.D.; Arndt, S.K.; Roxburgh, S.H.; Bennett, L.T. Climate more important than soils for predicting forest biomass at the continental scale. Ecography 2020, 43, 1692–1705. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Arets, E.J.; Ascarrunz, N.; Enquist, B.J.; Finegan, B.; Licona, J.C.; Martínez-Ramos, M.; Mazzei, L.; Meave, J.A. Biodiversity and climate determine the functioning of Neotropical forests. Glob. Ecol. Biogeogr. 2017, 26, 1423–1434. [Google Scholar] [CrossRef]
- Bean, W.T.; Stafford, R.; Brashares, J.S. The effects of small sample size and sample bias on threshold selection and accuracy assessment of species distribution models. Ecography 2012, 35, 250–258. [Google Scholar] [CrossRef]
- Stockwell, D.R.B.; Peterson, A.T. Effects of sample size on accuracy of species distribution models. Ecol. Model. 2002, 148, 1–13. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Sun, X.; Li, G.; Wang, M.; Fan, Z. Analyzing the Uncertainty of Estimating Forest Aboveground Biomass Using Optical Imagery and Spaceborne LiDAR. Remote Sens. 2019, 11, 722. [Google Scholar] [CrossRef]
- Jackson, C.R.; Robertson, M.P. Predicting the potential distribution of an endangered cryptic subterranean mammal from few occurrence records. J. Nat. Conserv. 2011, 19, 87–94. [Google Scholar] [CrossRef]
- Proosdij, A.S.J.; Sosef, M.S.M.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2015, 39, 542–552. [Google Scholar] [CrossRef]
- Santoro, M.; Cartus, O.; Wegmüller, U.; Besnard, S.; Carvalhais, N.; Araza, A.; Herold, M.; Liang, J.; Cavlovic, J.; Engdahl, M.E. Global estimation of above-ground biomass from spaceborne C-band scatterometer observations aided by LiDAR metrics of vegetation structure. Remote Sens. Environ. 2022, 279, 113114. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGB Class | Number of Samples | Range of the AGB (t·hm−2) | Average (t·hm−2) | Median (t·hm−2) | Standard Deviation (t·hm−2) |
|---|---|---|---|---|---|
| 1 | 86 | <312 | 183.66 | 192.33 | 66.38 |
| 2 | 34 | 312~602 | 434.96 | 438.60 | 79.62 |
| 3 | 17 | 602~1071 | 784.33 | 789.25 | 99.89 |
| 4 | 7 | ≥1071 | 1422.41 | 1403.10 | 168.09 |
| Variables | Meaning | AGB Class | Variables | Meaning | AGB Class |
|---|---|---|---|---|---|
| Aspect | aspect (°) | 1; 2; 3; 4 | SWC | soil water content (volumetric %) for 33 kPa at 30 cm depth | 1; 2; 3 |
| Elevation | elevation (m) | 1; 2; 3 | Band2 | blue band (0.450–0.515 μm) | 1; 2 |
| Slope | slope (°) | 1; 2; 3; 4 | Band3 | green band (0.525–0.600 μm) | 1; 2 |
| Bio1 | annual mean temperature (°C) | 1; 2; 3 | Band4 | red band (0.636–0.680 μm) | 1; 2; 4 |
| Bio2 | mean diurnal range (mean of monthly (max temp–min temp)) (°C) | 1 | Band5 | near-infrared band (0.845–0.885 μm) | 1; 3 |
| Bio3 | isothermality (BIO2/BIO7) (×100) | 1 | Band7 | short-wave (length) infrared band (2.100–2.300 μm) | 2 |
| Bio4 | temperature seasonality (standard deviation ×100) | 2; 3 | BRI | brightness | 2 |
| Bio6 | min temperature of the coldest month (°C) | 2 | GRE | greenness | 2; 3 |
| Bio7 | temperature annual range (°C) | 1 | WET | wetness | 2; 3 |
| Bio8 | mean temperature of the wettest quarter (°C) | 1; 2; 3 | EVI | enhanced vegetation index | 2; 3 |
| Bio14 | precipitation of driest month (mm) | 2 | NDVI | normalized difference vegetation index | 1; 4 |
| Bio15 | precipitation seasonality (coefficient of variation) | 1; 2; 3 | COR2_W5 | correlation | 1; 2; 4 |
| Bio17 | precipitation of driest quarter (mm) | 3 | COR2_W7 | correlation | 1; 2 |
| Bio18 | precipitation of warmest quarter (mm) | 1; 2 | COR5_W5 | correlation | 1; 3 |
| CC | clay content in % (kg/kg) | 1; 3; 4 | COR5_W7 | correlation | 1; 2 |
| SC | sand content in % (kg/kg) | 2; 3 | HOM1_W7 | homogeneity | 1; 2; 3; 4 |
| AGB Class | Training AUC | Testing AUC | AUC Standard Deviation |
|---|---|---|---|
| 1 | 0.982 | 0.959 | 0.024 |
| 2 | 0.990 | 0.957 | 0.026 |
| 3 | 0.949 | 0.823 | 0.109 |
| 4 | 0.964 | 0.847 | 0.124 |
| Variables | Percent Contribution (%) | Variables | Percent Contribution (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1st AGB Class | 2nd AGB Class | 3rd AGB Class | 4th AGB Class | 1st AGB Class | 2nd AGB Class | 3rd AGB Class | 4th AGB Class | ||
| Altitude | 20.4 | 9.5 | 3.8 | SC | 2.4 | 1 | |||
| Aspect | 2.6 | 1 | 0.9 | 10.5 | Band2 | 0.5 | 0.4 | ||
| Slope | 5.1 | 12.7 | 0.5 | 15.8 | Band3 | 1.5 | 0.4 | ||
| Bio1 | 4.5 | 50.5 | Band4 | 2.3 | 1.5 | 26.7 | |||
| Bio2 | 4.8 | Band5 | 2 | 9.6 | |||||
| Bio3 | 0.6 | Band7 | 0.3 | ||||||
| Bio4 | 6.6 | 2.3 | BRI | 6.6 | |||||
| Bio6 | 4.9 | EVI | 1.8 | 0.7 | |||||
| Bio7 | 2 | 0.6 | GRE | 1.7 | 0.7 | ||||
| Bio8 | 0.4 | 4.2 | 1.6 | NDVI | 7.8 | 4.7 | 9.7 | ||
| Bio14 | 0.2 | WET | 0.8 | 0.8 | |||||
| Bio15 | 12 | 12.7 | 8.8 | COR2_W5 | 0.4 | 3 | 11.9 | ||
| Bio17 | 2.4 | COR2_W7 | 1.4 | 1.5 | |||||
| Bio18 | 20 | 15.7 | COR5_W5 | 0.8 | 2.4 | ||||
| CC | 1.2 | 0.7 | 9.3 | COR5_W7 | 1.7 | 1.3 | |||
| SWC | 7.8 | 4.8 | 10.4 | HOM1_W7 | 0.5 | 0.5 | 2.8 | 16.1 | |
| Contrast Items | Size of Sample | Fitted Equation | R-Value | Significance | RMSE | |
|---|---|---|---|---|---|---|
| K-Means clustering | Total AGB | 141 | y = 0.473x + 389.578 | 0.613 | 0.05 | 196.375 |
| 1st AGB class | 85 | y = −0.096 + 508.809 | 0.031 | 0.05 | 208.806 | |
| 2nd AGB class | 34 | y = 0.979x + 124.322 | 0.445 | 0.05 | 161.614 | |
| 3rd AGB class | 15 | y = 0.521x + 335.111 | 0.279 | 0.05 | 191.510 | |
| 4th AGB class | 7 | y = 0.182x + 881.589 | 0.242 | 0.05 | 144.930 | |
| Interval classification | Total AGB | 139 | y = 0.476x + 346.070 | 0.639 | 0.05 | 184.966 |
| ≤250 | 68 | y = −0.136x + 474.184 | 0.039 | 0.05 | 185.507 | |
| 250 < AGB ≤ 500 | 43 | y = 0.502x + 282.457 | 0.231 | 0.05 | 163.809 | |
| 500 < AGB ≤ 750 | 12 | y = =0.464x + 350.952 | 0.179 | 0.05 | 193.964 | |
| 750 < AGB ≤ 1000 | 9 | y = 0.798x + 91.597 | 0.198 | 0.05 | 263.067 | |
| AGB > 1000 | 7 | y = 0.332x + 629.763 | 0.355 | 0.05 | 173.885 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Xu, Z.; Wang, Y. Application of MaxEnt Model in Biomass Estimation: An Example of Spruce Forest in the Tianshan Mountains of the Central-Western Part of Xinjiang, China. Forests 2023, 14, 953. https://doi.org/10.3390/f14050953
Ding X, Xu Z, Wang Y. Application of MaxEnt Model in Biomass Estimation: An Example of Spruce Forest in the Tianshan Mountains of the Central-Western Part of Xinjiang, China. Forests. 2023; 14(5):953. https://doi.org/10.3390/f14050953
Chicago/Turabian StyleDing, Xue, Zhonglin Xu, and Yao Wang. 2023. "Application of MaxEnt Model in Biomass Estimation: An Example of Spruce Forest in the Tianshan Mountains of the Central-Western Part of Xinjiang, China" Forests 14, no. 5: 953. https://doi.org/10.3390/f14050953