
A Meta-Analysis of the Effects of Harvesting on the Abundance and Richness of Soil Fauna in Boreal and Temperate Forests

Abstract
:1. Introduction
- How do various groups of soil fauna respond to forest harvesting? Soil fauna comprises a diverse array of taxa that exhibit significant variations in biology and ecology. As a result, their response to forest harvesting can be highly heterogeneous [13]. However, existing research on this topic has primarily focused on one or two large taxa and rarely includes multiple groups [14,15]. Consequently, our comprehension of the differences in the responses of various soil fauna groups to logging and the reasons for these differences is limited.
- Does the type of harvesting and the type of forest have a modifying effect on soil fauna response? Forest harvesting can be carried out through various practices. Clear-cutting, which is historically the most common example of even-aged silviculture practice in temperate and boreal biomes [11], may result in significant changes in environmental conditions. This includes altered light, humidity, wind speed, and other conditions which can constrain forest biota, e.g., [16]. Partial cutting or retention forestry is another practice in which some parts of the trees are left on-site to maintain organic matter inputs and nutrient cycles [17] and provide a refuge for belowground organisms [18]. The response of soil fauna to harvesting may differ depending on the practice used [9,10,11]. However, we still have a poor understanding of the differences in the response of individual groups of soil fauna to harvesting practices. The forest type is another important factor that can modify the impact of forest harvesting on soil fauna. Coniferous and deciduous forests, for example, are quite different from each other in terms of soil and microclimatic conditions. This is reflected in the dissimilarity of the composition and structure of soil fauna [14,19,20]. Despite this knowledge, there is still a significant gap in our understanding of the differences in the reaction of soil fauna to harvesting in different forest types.
- What are the temporal dynamics of the forest harvesting effect on soil fauna? Harvesting leads to significant disturbance of the forest ecosystem, while also significantly activating succession processes. The high dynamics of changing conditions in such processes may determine temporal changes in the soil fauna response. However, due to the limited number of sampling periods in individual studies, we have limited knowledge of how quickly soil animals react to harvesting and what the rate of their recovery is.
2. Materials and Methods
2.1. Literature Search and Inclusion Criteria
2.2. Meta-Analysis
3. Results
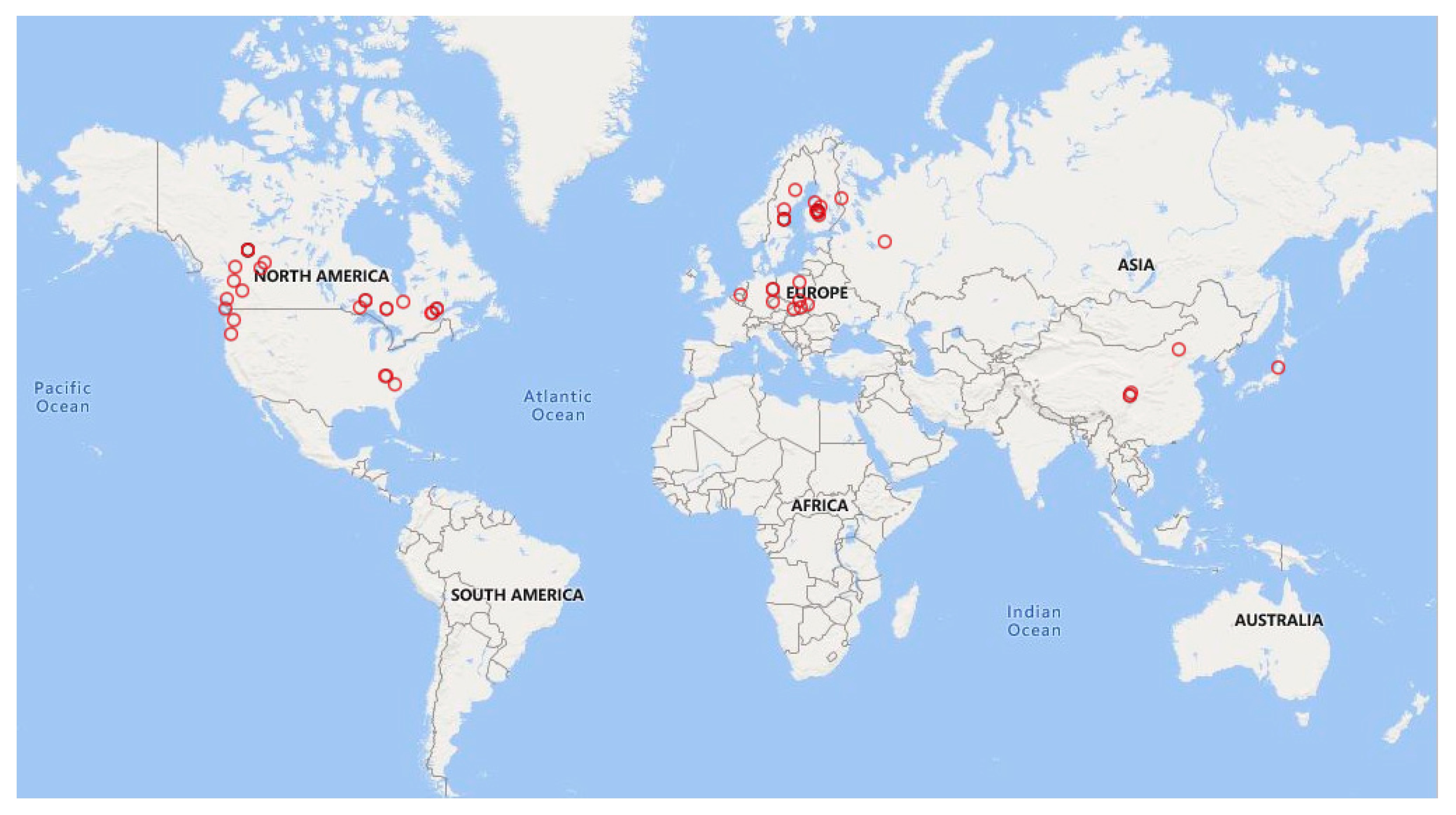
3.1. Dataset Description
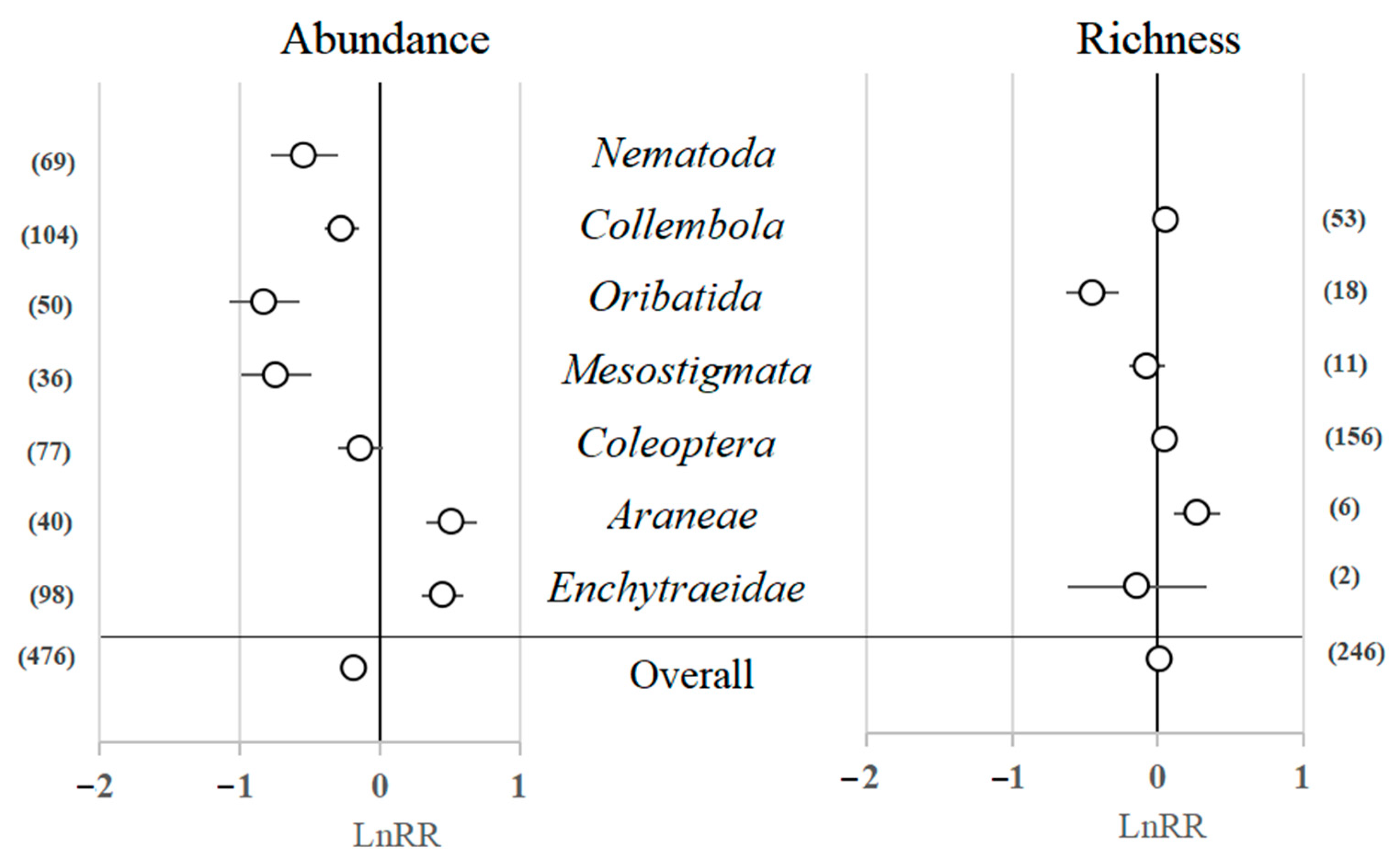
3.2. The Effect of Forest Harvesting on Different Groups of Soil Fauna
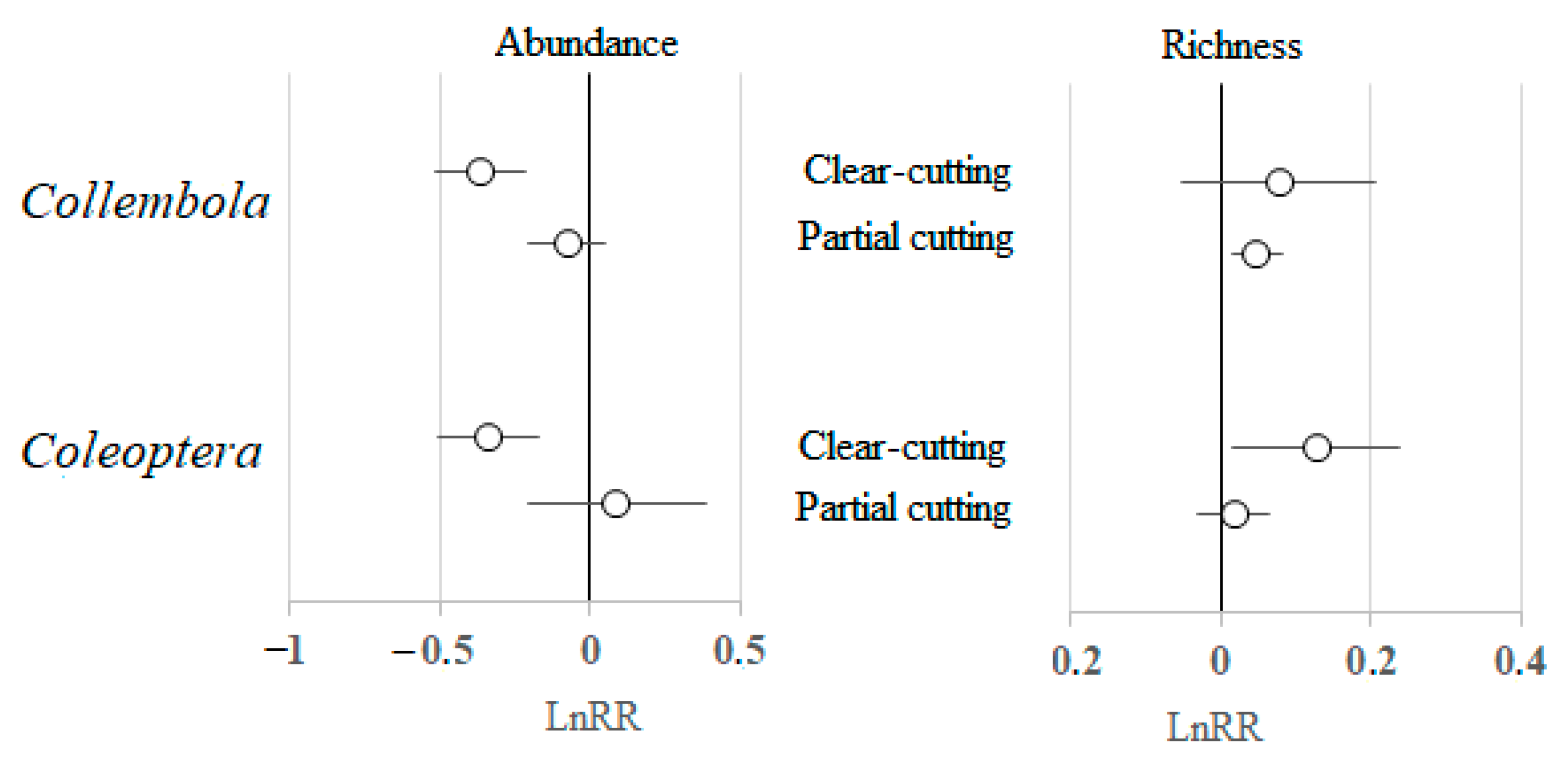
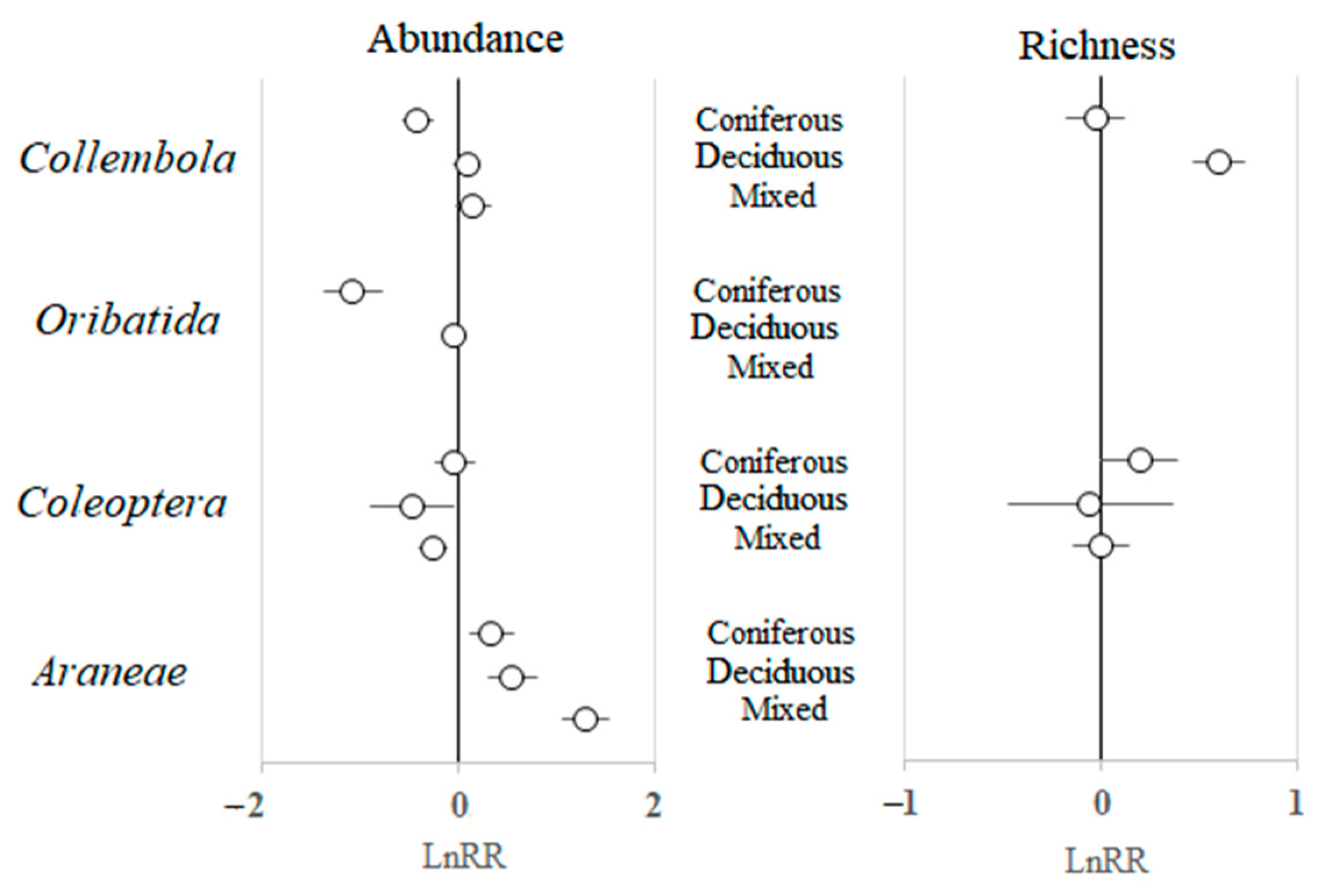
3.3. Type of Harvesting and Type of Forest as Modifying Factors of Soil Fauna Response
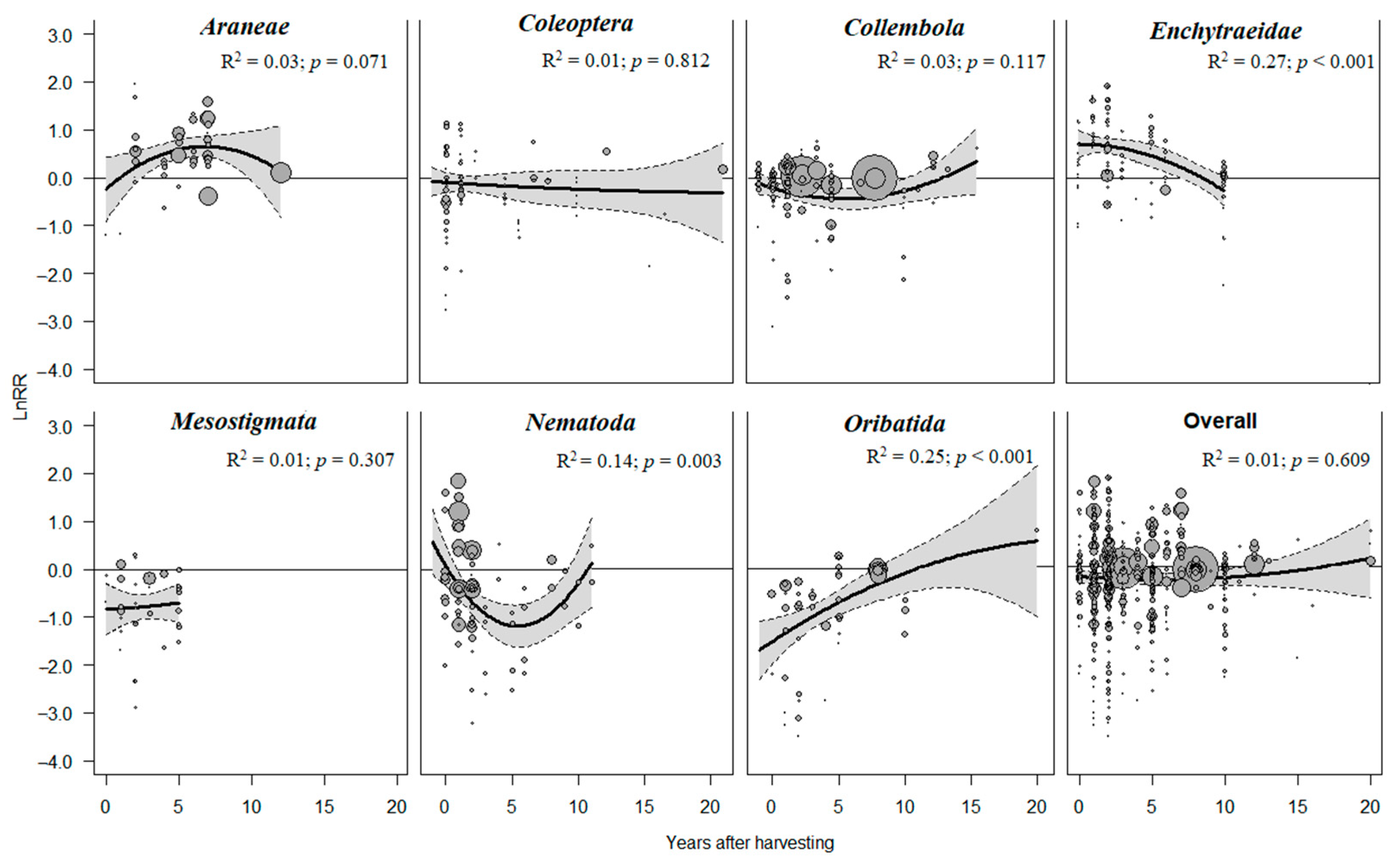
3.4. Temporal Dynamics of the Effects of Forest Harvesting on Soil Fauna
4. Discussion
4.1. Effect of Forest Harvesting on Different Groups of Soil Fauna
4.2. Types of Harvesting and Forests as Modifying Factors of Soil Fauna Response
4.3. Temporal Dynamics of the Effects of Forest Harvesting on Soil Fauna
4.4. Data Set Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef]
- Díaz, S.; Fargione, J.; Chapin, F.S.; Tilman, D. Biodiversity loss threatens human wellbeing. PLoS Biol. 2006, 4, e277. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment; Paper 163; Food and Agricultural Organization of the United Nations: Rome, Italy; FAO Forestry: Rome, Italy, 2010; pp. 1–378. [Google Scholar]
- Global Biodiversity. Outlook 3; Secretariat of the Convention on Biological Diversity: Montreal, QC, USA, 2010; p. 94. [Google Scholar]
- James, J.; Harrison, J. The Effect of Harvest on Forest Soil Carbon: A Meta-Analysis. Forests 2016, 7, 308. [Google Scholar] [CrossRef]
- Jerabkova, L.; Prescott, C.E.; Titus, B.D.; Hope, G.D.; Walters, M.B. A meta-analysis of the effects of clearcut and variable-retention harvesting on soil nitrogen fluxes in boreal and temperate forests. Can. J. For. Res. 2011, 41, 1852–1870. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y. The Effect of Harvest on Forest Soil N2O Fluxes: A Review. In The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–19. [Google Scholar] [CrossRef]
- Michał, B.; Rafał, Z. Responses of small mammals to clear-cutting in temperate and boreal forests of Europe: A meta-analysis and review. Eur. J. For. Res. 2014, 133, 1–11. [Google Scholar] [CrossRef]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Odor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, E.; et al. Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of forest management on species richness: Global meta-analysis and economic trade-offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef]
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Malmström, A.; Persson, T.; Ahlström, K.; Gongalsky, K.B.; Bengtsson, J. Dynamics of soil meso and macrofauna during a 5-year period after clear-cut burning in a boreal forest. Appl. Soil Ecol. 2009, 43, 61–74. [Google Scholar] [CrossRef]
- Hasegawa, M.; Fukuyama, K.; Makino, S.; Okochi, I.; Goto, H.; Mizoguchi, T.; Sakata, T.; Tanaka, H. Collembolan community dynamics during deciduous forests regeneration in Japan. Pedobiologia 2006, 50, 117–126. [Google Scholar] [CrossRef]
- Háněl, L. Response of soil nematodes inhabiting spruce forests in the Šumava Mountains to disturbance by bark beetles and clear-cutting. For. Ecol. Manag. 2004, 202, 209–225. [Google Scholar] [CrossRef]
- Heithecker, T.D.; Halpern, C.B. Edge-related gradients in microclimate in forest aggregates following structural retention harvests in western Washington. For. Ecol. Manag. 2007, 248, 163–173. [Google Scholar] [CrossRef]
- Dahlgren, R.A.; Driscoll, C.T. The effects of whole-tree clear-cutting on soil processes at the Hubbard Brook Experimental Forest, New Hampshire, USA. Plant. Soil 1994, 158, 239–262. [Google Scholar] [CrossRef]
- Barg, A.K.; Edmonds, R.L. Influence of partial cutting on site microclimate, soil nitrogen dynamics, and microbial biomass in Douglas-fir stands in western Washington. Can. J. For. 1999, 29, 705–713. [Google Scholar] [CrossRef]
- Van Den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef]
- Fujii, S.; Berg, M.P.; Cornelissen, J.H. Living litter: Dynamic trait spectra predict fauna composition. Trends Ecol. Evol. 2020, 35, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Potapov, A.M.; Rozanova, O.L.; Semenina, E.E.; Leonov, V.D.; Belyakova, O.I.; Bogatyreva, V.Y.; Degtyarev, M.I.; Esaulov, A.S.; Korotkevich, A.Y.; Kudrin, A.A.; et al. Size compartmentalization of energy channeling in terrestrial belowground food webs. Ecology 2021, 102, e03421. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Abgrall, C.; Forey, E.; Chauvat, M. Soil fauna responses to invasive alien plants are determined by trophic groups and habitat structure: A global meta-analysis. Oikos 2019, 128, 1390–1401. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://wwwR-project.org/ (accessed on 15 January 2023).
- Viechtbauer, W. Conducting meta-analyses in R with the metafor. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A., Jr. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Zaitsev, A.S. The role of spatial heterogeneity of the environment in soil fauna recovery after fires. Dokl. Earth Sci. 2016, 471, 1265–1268. [Google Scholar] [CrossRef]
- Koivula, M.J. Under which conditions does retention harvesting support ground beetles of boreal forests? Balt. J. Coleopterol. 2012, 12, 7–26. [Google Scholar]
- Niemelä, J.; Koivula, M.; Kotze, D.J. The effects of forestry on carabid assemblages in boreal forests. J. Insect Conserv. 2007, 11, 5–18. [Google Scholar] [CrossRef]
- Pohl, G.R.; Langor, D.W.; Spence, J.R. Rove beetles and ground beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of harvest and regeneration practices in western Canadian Foothills forests. Biol. Conserv. 2007, 137, 294–307. [Google Scholar] [CrossRef]
- Zaitsev, A.A.; Gongalsky, K.B.; Malmström, A.; Persson, T.; Bengtsson, J. Why are forest fires generally neglected in soil fauna research? A mini-review. Appl. Soil Ecol. 2016, 98, 261–271. [Google Scholar] [CrossRef]
- Pressler, Y.; Moore, J.C.; Cotrufo, M.F. Belowground community responses to fire: Meta-analysis reveals contrasting responses of soil microorganisms and mesofauna. Oikos 2018, 128, 309–327. [Google Scholar] [CrossRef]
- Hedde, M.; van Oort, F.; Lamy, I. Functional traits of soil invertebrates as indicators for exposure to soil disturbance. Environ. Pollut. 2012, 164, 59–65. [Google Scholar] [CrossRef]
- Langlands, P.R.; Brennan, K.E.C.; Framenau, V.W.; Main, B.Y. Predicting the post-fire responses of animal assemblages: Testing a trait-based approach using spiders. J. Anim. Ecol. 2011, 80, 558–568. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Podgaiski, L.R.; Joner, F.; Lavorel, S.; Moretti, M.; Ibanez, S.; Mendonça, M.d.S., Jr.; Pillar, V.D. Spider trait assembly patterns and resilience under fire-induced vegetation change in south Brazilian grasslands. PLoS ONE 2013, 8, e60207. [Google Scholar] [CrossRef]
- Buckingham, S.; Murphy, N.; Gibb, H. The effects of fire severity on macroinvertebrate detritivores and leaf litter decomposition. PLoS ONE 2015, 10, e0124556. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.J.; Gillooly, A.A.; Savage, V.; West, G. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Woodward, G.; Ebenman, B.; Emmerson, M.; Montoya, J.M.; Olesen, J.M.; Valido, A.; Warren, P.H. Body size in ecological networks. Trends Ecol. Evol. 2005, 20, 402–409. [Google Scholar] [CrossRef]
- Turnbull, M.S.; George, P.B.; Lindo, Z. Weighing in: Size spectra as a standard tool in soil community analyses. Soil Biol. Biochem. 2014, 68, 366–372. [Google Scholar] [CrossRef]
- Negro, M.; Caprio, E.; Leo, K.; Maritano, U.; Roggero, A.; Vacchiano, G.; Claudia, P.; Rolando, A. The effect of forest management on endangered insects assessed by radio-tracking: The case of the ground beetle Carabus olympiae in European beech Fagus sylvatica stands. For. Ecol. Manag. 2017, 406, 125–137. [Google Scholar] [CrossRef]
- Buckingham, S.; Murphy, N.; Gibb, H. Effects of fire severity on the composition and functional traits of litter-dwelling macroinvertebrates in a temperate forest. For. Ecol. Manag. 2019, 434, 279–288. [Google Scholar] [CrossRef]
- Ojala, R.; Huhta, V. Dispersal of microarthropods in forest soil. Pedobiologia 2001, 45, 443–450. [Google Scholar] [CrossRef]
- Gongalsky, K.B. Perfugia as a mechanism for the recovery of soil fauna after ecosystem disturbances. Russ. J. Ecosyst. Ecol. 2017, 2, 1–12. [Google Scholar] [CrossRef]
- Dechene, A.; Buddle, C. Effects of experimental forest harvesting on oribatid mite biodiversity. For. Ecol. Manag. 2009, 258, 1331–1341. [Google Scholar] [CrossRef]
- Lóšková, J.; Luptacik, P.; Miklisová, D.; Kovac, L. The effect of clear-cutting and wildfire on soil Oribatida (Acari) in windthrown stands of the High Tatra Mountains (Slovakia). Eur. J. Soil Biol. 2013, 55, 131–138. [Google Scholar] [CrossRef]
- Lehmitz, R.; Russell, D.; Hohberg, K.; Christian, A.; Xylander, W.E. Active dispersal of oribatid mites into young soils. Appl. Soil Ecol. 2012, 55, 10–19. [Google Scholar] [CrossRef]
- Elek, Z.; Růžičková, J.; Ódor, P. Individual decisions drive the changes in movement patterns of ground beetles between forestry management types. Biologia 2021, 76, 3287–3296. [Google Scholar] [CrossRef]
- Bååth, E.; Frostegård, A.; Pennanen, T.; Fritze, H. Microbial community structure and pH response in relation to soil organic matter quality in wood ash fertilized, clear-cut or burned coniferous forest soils. Soil Biol. Biochem. 1995, 27, 229–240. [Google Scholar] [CrossRef]
- Moore-Kucera, J.; Dick, R.P. PLFA profiling of microbial community structure and seasonal shifts in soils of a Douglas-fir chronosequence. Microb. Ecol. 2008, 55, 500–511. [Google Scholar] [CrossRef]
- Hartmann, M.; Howes, C.; VanInsberghe, D.; Yu, H.; Bachar, D.; Christen, R.; Nilsson, R.H.; Hallam, S.; Mohn, W. Significant and persistent impact of timber harvesting on soil microbial communities in Northern coniferous forests. ISME J. 2012, 6, 2320. [Google Scholar] [CrossRef]
- Forge, T.; Simard, S. Trophic structure of nematode communities, microbial biomass, and nitrogen mineralization in soils of forests and clearcuts in the southern interior of British Columbia. Can. J. Soil Sci. 2000, 80, 401–410. [Google Scholar] [CrossRef]
- Remén, C.; Persson, T.; Finlay, R.; Ahlström, K. Responses of Oribatid mites to tree girdling and nutrient addition in boreal coniferous forests. Soil Biol. Biochem. 2008, 40, 2881–2890. [Google Scholar] [CrossRef]
- Kudrin, A.A.; Zuev, A.G.; Taskaeva, A.A.; Konakova, T.N.; Kolesnikova, A.A.; Gruzdev, I.V.; Gabov, D.N.; Yakovleva, E.V.; Tiunov, A.V. Spruce girdling decreases abundance of fungivorous soil nematodes in a boreal forest. Soil Biol. Biochem. 2021, 155, 108184. [Google Scholar] [CrossRef]
- Londo, A.J.; Messina, M.G.; Schoenholtz, S.H. Forest harvesting effects on soil temperature, moisture, and respiration in a bottomland hardwood forest. Soil Sci. Soc. Am. J. 1999, 63, 637–644. [Google Scholar] [CrossRef]
- Holden, S.R.; Treseder, K.K. A meta-analysis of soil microbial biomass responses to forest disturbances. Front. Microbiol. 2013, 4, 163. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef]
- Battigelli, J.P.; Spence, J.R.; Langor, D.W.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Can. J. For. Res. 2004, 34, 1136–1149. [Google Scholar] [CrossRef]
- Farská, J.; Prejzková, K.; Rusek, J. Management intensity affects traits of soil microarthropod community in montane spruce forest. Appl. Soil Ecol. 2014, 75, 71–79. [Google Scholar] [CrossRef]
- Rousseau, L.; Venier, L.; Fleming, R.; Hazlett, P.; Morris, D.; Handa, I. Long-term effects of biomass removal on soil mesofaunal communities in northeastern Ontario (Canada) jack pine (Pinus banksiana) stands. For. Ecol. Manag. 2018, 421, 72–83. [Google Scholar] [CrossRef]
- Sabais, A.; Scheu, S.; Eisenhauer, N. Plant species richness drives the density and diversity of Collembola in temperate grassland. Acta Oecologica 2011, 37, 195–202. [Google Scholar] [CrossRef]
- Siira-Pietikäinen, A.; Haimi, J. Changes in soil fauna 10 years after forest harvestings: Comparison between clear felling and green-tree retention methods. For. Ecol. Manag. 2009, 258, 332–338. [Google Scholar] [CrossRef]
- Saksa, T. Regeneration after stump harvesting in southern Finland. For. Ecol. Manag. 2013, 290, 79–82. [Google Scholar] [CrossRef]
- Lehtosalo, M.; Mäkelä, A.; Valkonen, S. Regeneration and tree growth dynamics of Picea abies, Betula pendula and Betula pubescens in regeneration areas treated with spot mounding in southern Finland. Scand. J. For. Res. 2010, 25, 213–223. [Google Scholar] [CrossRef]
- Wallander, H.; Johansson, U.; Sterkenburg, E.; Durling, M.B.; Lindahl, B.D. Production of ectomycorrhizal mycelium peaks during canopy closure in Norway spruce forests. New Phytol. 2010, 187, 1124–1134. [Google Scholar] [CrossRef]
- Pollierer, M.M.; Scheu, S. Stable isotopes of amino acids indicate that soil decomposer microarthropods pre-dominantly feed on saprotrophic fungi. Ecosphere 2021, 12, e03425. [Google Scholar] [CrossRef]
- Antsiferov, A.L. The details of strcture and development of ground beetle communities (Coleoptera, Carabidae) in annual felling areas in spruce forests of Kostroma’s Zavolzhje, Russia. Euroasian Entomol. J. 2016, 15, 463–476. [Google Scholar]
- Addison, J.A.; Barber, K.N. Response of Soil Invertebrates to Clearcutting and Partial Cutting in a Boreal Mixedwood Forest in Northern Ontario. Available online: https://cfs.nrcan.gc.ca/pubwarehouse/pdfs/9135.pdf (accessed on 15 January 2023).
- Chang, L.; Baifeng, W.A.N.G.; Xianghui, L.I.U.; Callaham Jr, M.A.; Feng, G.E. Recovery of Collembola in Pinus tabulaeformis plantations. Pedosphere 2017, 27, 129–137. [Google Scholar] [CrossRef]
- Chauvat, M.; Zaitsev, A.S.; Wolters, V. Successional changes of Collembola and soil microbiota during forest rotation. Oecologia 2003, 137, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Cobb, T.P.; Langor, D.W.; Spence, J.R. Biodiversity and multiple disturbances: Boreal forest ground beetle (Coleoptera: Carabidae) responses to wildfire, harvesting, and herbicide. Can. J. For. Res. 2007, 37, 1310–1323. [Google Scholar] [CrossRef]
- Čuchta, P.; Miklisová, D.; Kováč, Ľ. The succession of soil Collembola communities in spruce forests of the High Tatra Mountains five years after a windthrow and clear-cut logging. For. Ecol. Manag. 2019, 433, 504–513. [Google Scholar] [CrossRef]
- de Warnaffe, G.D.B.; Lebrun, P. Effects of forest management on carabid beetles in Belgium: Implications for biodiversity conservation. Biol. Conserv. 2004, 118, 219–234. [Google Scholar] [CrossRef]
- Duchesne, L.C.; Lautenschlager, R.A.; Bell, F.W. Effects of clear-cutting and plant competition control methods on carabid (Coleoptera: Carabidae) assemblages in northwestern Ontario. Environ. Monit. Assess. 1999, 56, 87–96. [Google Scholar] [CrossRef]
- Edmonds, R.L.; Marra, J.L.; Barg, A.K.; Sparks, G.B. Influence of Forest Harvesting on Soil Organisms and Decomposition in Western Washington. 2000. Available online: https://citeseerx.ist.psu.edu/viewdoc/download;jsessionid=74700AC79554C7095EB69914E80755D8?doi=10.1.1.164.8749&rep=rep1&type=pdf (accessed on 15 January 2023).
- Elek, Z.; Kovács, B.; Aszalós, R.; Boros, G.; Samu, F.; Tinya, F.; Ódor, P. Taxon-specific responses to different forestry treatments in a temperate forest. Sci. Rep. 2018, 8, 16990. [Google Scholar] [CrossRef] [PubMed]
- George, P.B.; Lindo, Z. Application of body size spectra to nematode trait-index analyses. Soil Biol. Biochem. 2015, 84, 15–20. [Google Scholar] [CrossRef]
- Halaj, J.; Halpern, C.B.; Yi, H. Responses of litter-dwelling spiders and carabid beetles to varying levels and patterns of green-tree retention. For. Ecol. Manag. 2008, 255, 887–900. [Google Scholar] [CrossRef]
- Heneghan, L.; Salmore, A.; Crossley, D.A., Jr. Recovery of decomposition and soil microarthropod communities in an Appalachian watershed two decades after a clearcut. For. Ecol. Manag. 2004, 189, 353–362. [Google Scholar] [CrossRef]
- Johansson, T.; Hjältén, J.; Olsson, J.; Dynesius, M.; Roberge, J.M. Long-term effects of clear-cutting on epigaeic beetle assemblages in boreal forests. For. Ecol. Manag. 2016, 359, 65–73. [Google Scholar] [CrossRef]
- Kataja-Aho, S.; Saari, E.; Fritze, H.; Haimi, J. Effects of stump removal on soil decomposer communities in undisturbed patches of the forest floor. Scand. J. For. Res. 2011, 26, 221–231. [Google Scholar] [CrossRef]
- Kataja-aho, S.; Fritze, H.; Haimi, J. Short-term responses of soil decomposer and plant communities to stump harvesting in boreal forests. For. Ecol. Manag. 2011, 262, 379–388. [Google Scholar] [CrossRef]
- Kataja-aho, S.; Hannonen, P.; Liukkonen, T.; Rosten, H.; Koivula, M.J.; Koponen, S.; Haimi, J. The arthropod community of boreal Norway spruce forests responds variably to stump harvesting. For. Ecol. Manag. 2016, 371, 75–83. [Google Scholar] [CrossRef]
- Klimaszewski, J.; Langor, D.W.; Work, T.T.; Pelletier, G.; Hammond, H.J.; Germain, C. The effects of patch harvesting and site preparation on ground beetles (Coleoptera, Carabidae) in yellow birch dominated forests of southeastern Quebec. Can. J. For. Res. 2005, 35, 2616–2628. [Google Scholar] [CrossRef]
- Koivula, M.J.; Venn, S.; Hakola, P.; Niemelä, J. Responses of boreal ground beetles (Coleoptera, Carabidae) to different logging regimes ten years post-harvest. For. Ecol. Manag. 2019, 436, 27–38. [Google Scholar] [CrossRef]
- Larrivée, M.; Fahrig, L.; Drapeau, P. Effects of a recent wildfire and clearcuts on ground-dwelling boreal forest spider assemblages. Can. J. For. Res. 2005, 35, 2575–2588. [Google Scholar] [CrossRef]
- Lindo, Z.; Visser, S. Forest floor microarthropod abundance and oribatid mite (Acari: Oribatida) composition following partial and clear-cut harvesting in the mixedwood boreal forest. Can. J. For. Res. 2004, 34, 998–1006. [Google Scholar] [CrossRef]
- Lundkvist, H. Effects of clear-cutting on the enchytraeids in a Scots pine forest soil in central Sweden. J. Appl. Ecol. 1983, 873–885. [Google Scholar] [CrossRef]
- Luo, T.H.; Yu, X.D.; Zhou, H.Z. Effects of reforestation practices on staphylinid beetles (Coleoptera: Staphylinidae) in southwestern China forests. Environ. Entomol. 2013, 42, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Magura, T.; Tóthmérész, B.; Elek, Z. Diversity and composition of carabids during a forestry cycle. Biodivers. Conserv. 2003, 12, 73–85. [Google Scholar] [CrossRef]
- Martikainen, P.; Kouki, J.; Heikkala, O. The effects of green tree retention and subsequent prescribed burning on ground beetles (Coleoptera: Carabidae) in boreal pine–dominated forests. Ecography 2006, 29, 659–670. [Google Scholar] [CrossRef]
- Moore, J.D.; Ouimet, R.; Camiré, C.; Houle, D. Effects of two silvicultural practices on soil fauna abundance in a northern hardwood forest, Québec, Canada. Can. J. Soil sci. 2002, 82, 105–113. [Google Scholar] [CrossRef]
- Nieminen, J.K. Are spruce boles hot spots for enchytraeids in clear-cut areas? Boreal Environ. Res. 2009, 14, 382–388. [Google Scholar]
- Pinzon, J.; Spence, J.R.; Langor, D.W. Responses of ground-dwelling spiders (Araneae) to variable retention harvesting practices in the boreal forest. For. Ecol. Manag. 2012, 266, 42–53. [Google Scholar] [CrossRef]
- Pinzon, J.; Spence, J.R.; Langor, D.W. Effects of prescribed burning and harvesting on ground-dwelling spiders in the Canadian boreal mixedwood forest. Biodivers. Conserv. 2013, 22, 1513–1536. [Google Scholar] [CrossRef]
- Panesar, T.S.; Marshall, V.G.; Barclay, H.J. The impact of clearcutting and partial harvesting systems on population dynamics of soil nematodes in coastal Douglas-fir forests. Pedobiologia 2000, 44, 641–665. [Google Scholar] [CrossRef]
- Rousseau, L.; Venier, L.; Aubin, I.; Gendreau-Berthiaume, B.; Moretti, M.; Salmon, S.; Handa, I.T. Woody biomass removal in harvested boreal forest leads to a partial functional homogenization of soil mesofaunal communities relative to unharvested forest. Soil Biol. Biochem. 2019, 133, 129–136. [Google Scholar] [CrossRef]
- Saint-Germain, M.; Larrivée, M.; Drapeau, P.; Fahrig, L.; Buddle, C.M. Short-term response of ground beetles (Coleoptera: Carabidae) to fire and logging in a spruce-dominated boreal landscape. For. Ecol. Manag. 2005, 212, 118–126. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley III, D.A. Microarthropod response following cable logging and clear-cutting in the southern Appalachians. Ecology 1981, 62, 126–135. [Google Scholar] [CrossRef]
- Siira-Pietikäinen, A.; Pietikäinen, J.; Fritze, H.; Haimi, J. Short-term responses of soil decomposer communities to forest management: Clear felling versus alternative forest harvesting methods. Can. J. For. Res. 2001, 31, 88–99. [Google Scholar] [CrossRef]
- Sławska, M.; Sławski, M. Does alternative methods of oak plantations establishment create favourable conditions for forest collembolan assemblages (Collembola, Hexapoda)? Sylwan 2018, 162, 580–589. [Google Scholar]
- Sohlenius, B. Short-term influence of clear-cutting on abundance of soil-microfauna (Nematoda, Rotatoria and Tardigrada) in a Swedish pine forest soil. J. Appl. Ecol. 1982, 349–359. [Google Scholar] [CrossRef]
- Sohlenius, B. Influence of clear-cutting and forest age on the nematode fauna in a Swedish pine forest soil. Appl. Soil Ecol. 2002, 19, 261–277. [Google Scholar] [CrossRef]
- Sterzyńska, M.; Shrubovych, J.; Kaprus, I. Effect of hydrologic regime and forest age on Collembola in riparian forests. Appl. Soil Ecol. 2014, 75, 199–209. [Google Scholar] [CrossRef]
- Toivanen, T.; Heikkilä, T.; Koivula, M.J. Emulating natural disturbances in boreal Norway spruce forests: Effects on ground beetles (Coleoptera, Carabidae). For. Ecol. Manag. 2014, 314, 64–74. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L.; Horn, S.; Kilgo, J.C.; Moorman, C.E. The response of ground beetles (Coleoptera: Carabidae) to selection cutting in a South Carolina bottomland hardwood forest. Biodivers. Conserv. 2006, 15, 261–274. [Google Scholar] [CrossRef]
- Wu, L.; He, F.; Spence, J.R. Recovery of a boreal ground-beetle (Coleoptera: Carabidae) fauna 15 years after variable retention harvest. J. Appl. Ecol. 2000, 57, 1717–1729. [Google Scholar] [CrossRef]
- Yu, X.D.; Luo, T.H.; Zhou, H.Z. Distribution of ground-dwelling beetles (Coleoptera) across a forest-clearcut ecotone in Wolong Natural Reserve, southwestern China. Insect Sci. 2006, 13, 127–135. [Google Scholar] [CrossRef]
- Yu, X.D.; Luo, T.H.; Zhou, H.Z. Distribution of carabid beetles among 40-year-old regenerating plantations and 100-year-old naturally regenerated forests in Southwestern China. For. Ecol. Manag. 2008, 255, 2617–2625. [Google Scholar] [CrossRef]
- Zaitsev, A.S.; Chauvat, M.; Pflug, A.; Wolters, V. Oribatid mite diversity and community dynamics in a spruce chronosequence. Soil Biol. Biochem. 2002, 34, 1919–1927. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Fauna | Abundance | Richness | ||
|---|---|---|---|---|
| Harvesting Type | Forest Type | Harvesting Type | Forest Type | |
| Nematoda | 0.151 | - | - | - |
| Collembola | 0.006 | <0.001 | 0.737 | <0.001 |
| Oribatida | - | <0.001 | - | - |
| Mesostigmata | - | - | - | - |
| Coleoptera | <0.001 | 0.086 | 0.038 | 0.001 |
| Araneae | 0.244 | 0.004 | - | - |
| Enchytraeidae | 0.528 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudrin, A.; Perminova, E.; Taskaeva, A.; Ditts, A.; Konakova, T. A Meta-Analysis of the Effects of Harvesting on the Abundance and Richness of Soil Fauna in Boreal and Temperate Forests. Forests 2023, 14, 923. https://doi.org/10.3390/f14050923
Kudrin A, Perminova E, Taskaeva A, Ditts A, Konakova T. A Meta-Analysis of the Effects of Harvesting on the Abundance and Richness of Soil Fauna in Boreal and Temperate Forests. Forests. 2023; 14(5):923. https://doi.org/10.3390/f14050923
Chicago/Turabian StyleKudrin, Alexey, Evgenia Perminova, Anastasia Taskaeva, Alla Ditts, and Tatiana Konakova. 2023. "A Meta-Analysis of the Effects of Harvesting on the Abundance and Richness of Soil Fauna in Boreal and Temperate Forests" Forests 14, no. 5: 923. https://doi.org/10.3390/f14050923