

Health and Growth of Black Pine outside Its Natural Distribution Range in the Romanian Carpathians

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
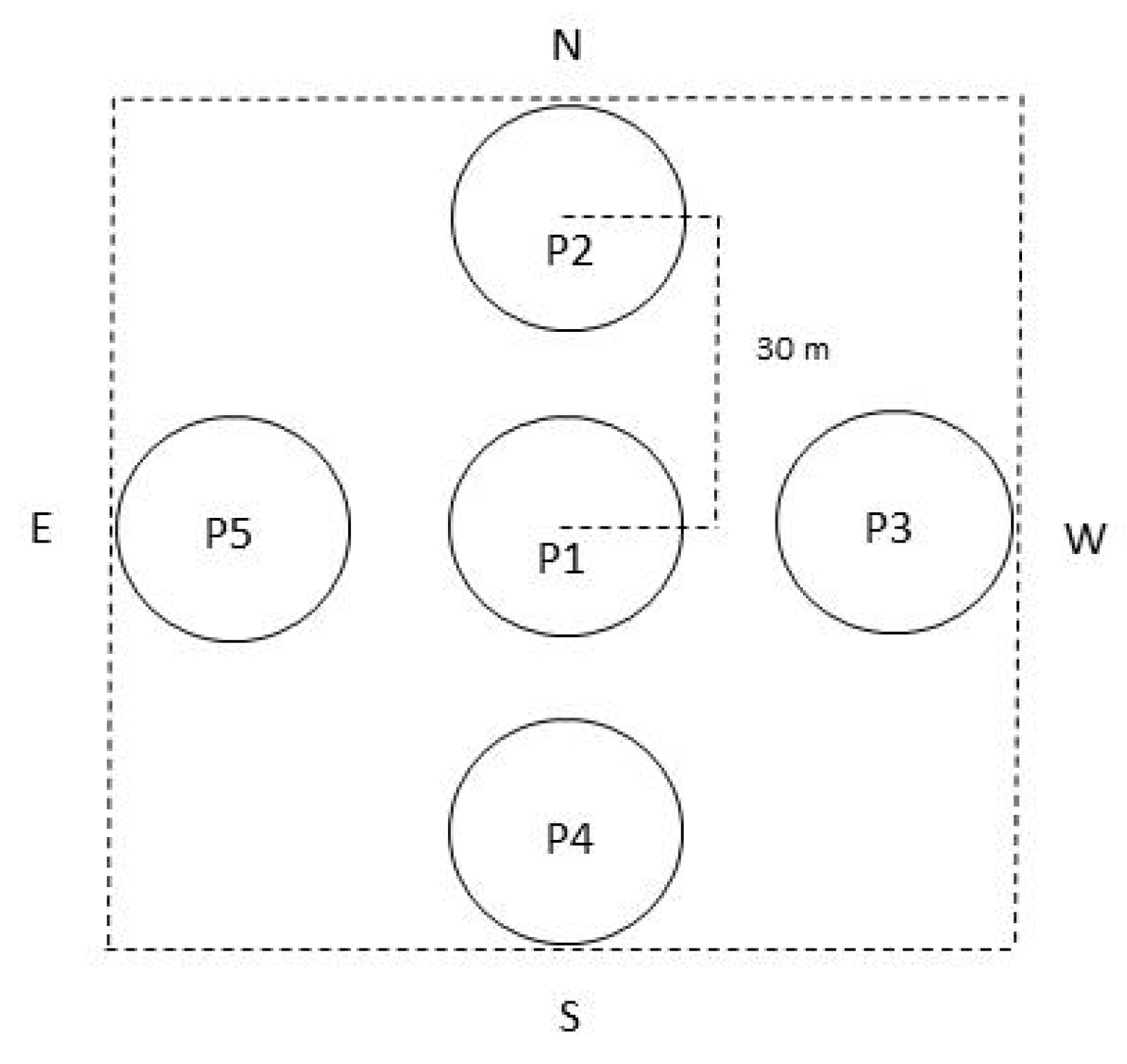
2.1. Study Area
2.2. Crown Defoliation
2.3. Radial Growth
2.4. Data Processing
2.4.1. Defoliation Explained by Biometric Variables (at Tree Level and Stand Level)
2.4.2. Relationship between Radial Growth and Crown Defoliation
2.4.3. Growth Driven by Climate
3. Results
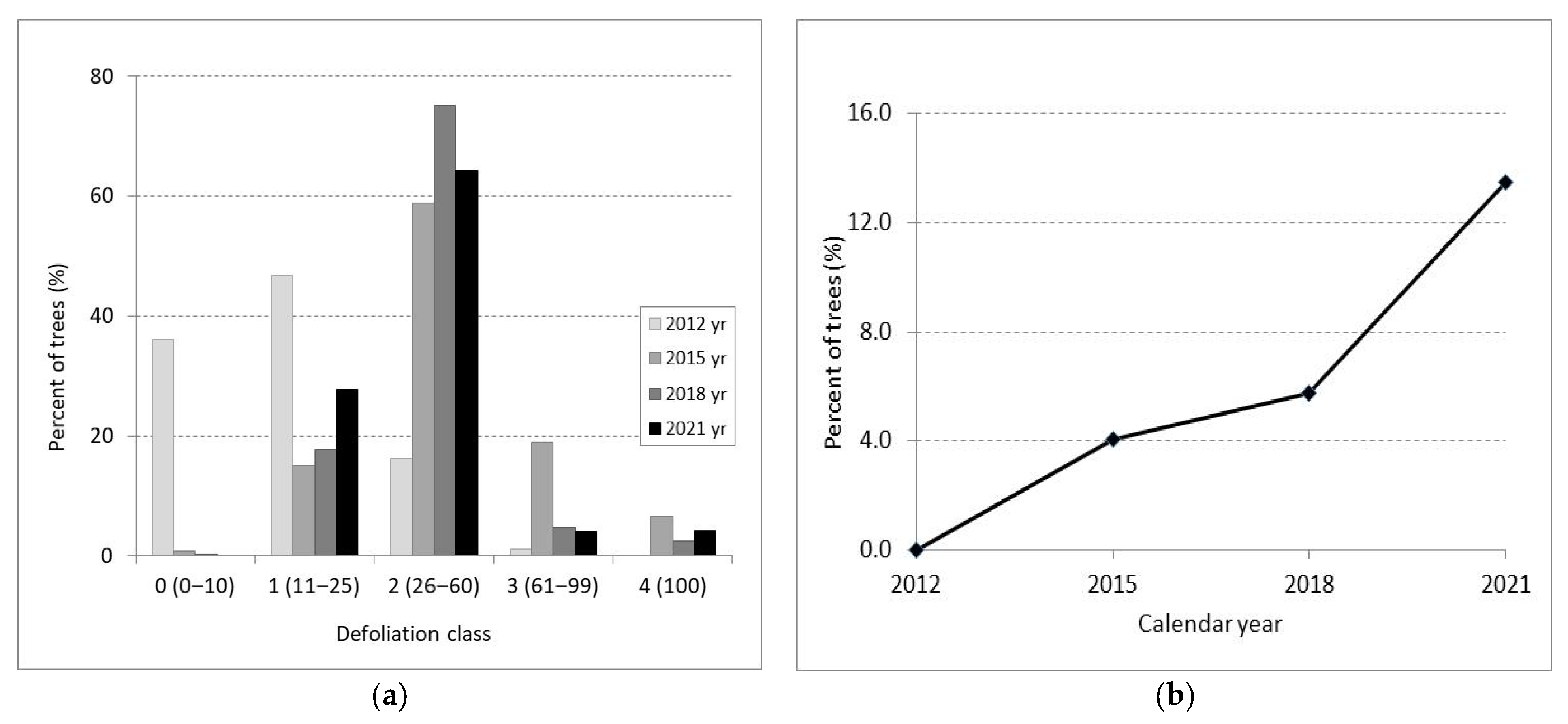
3.1. Crown Defoliation
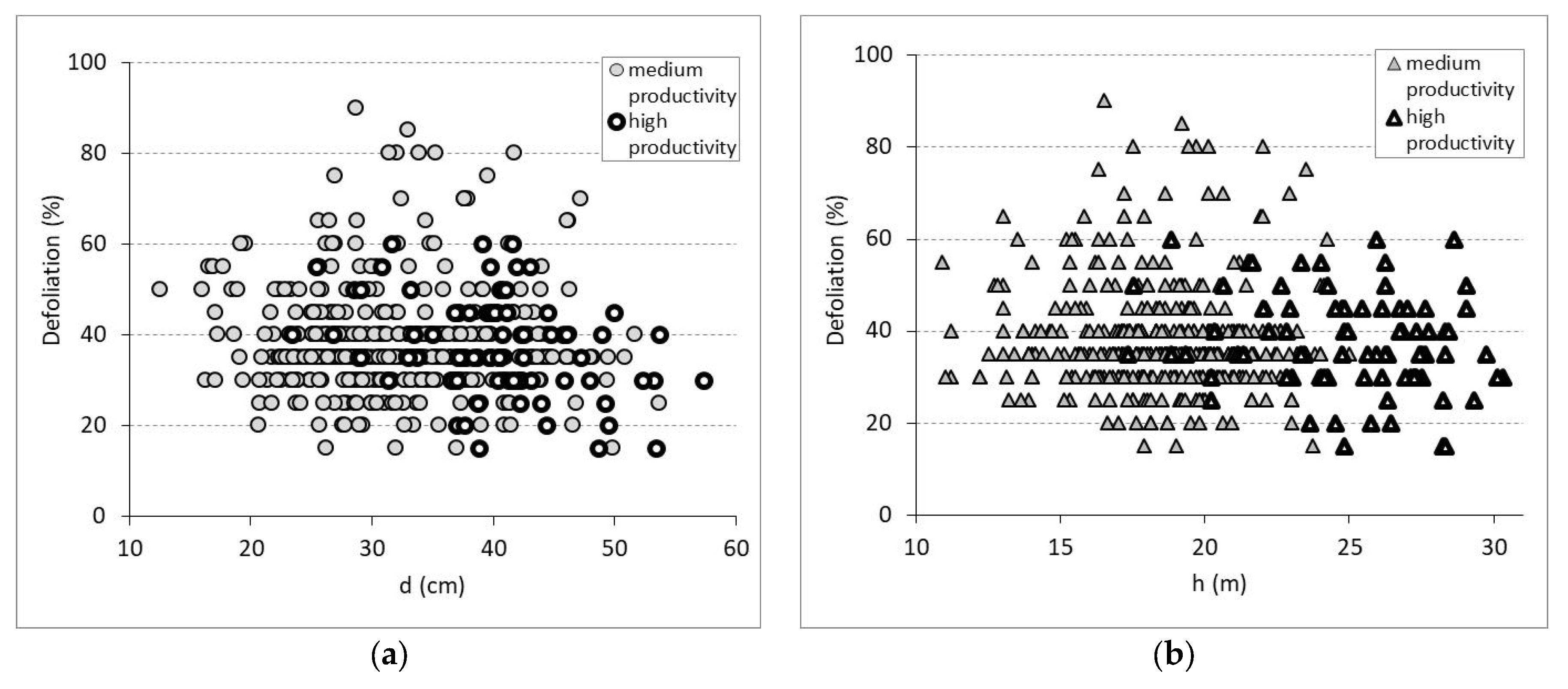
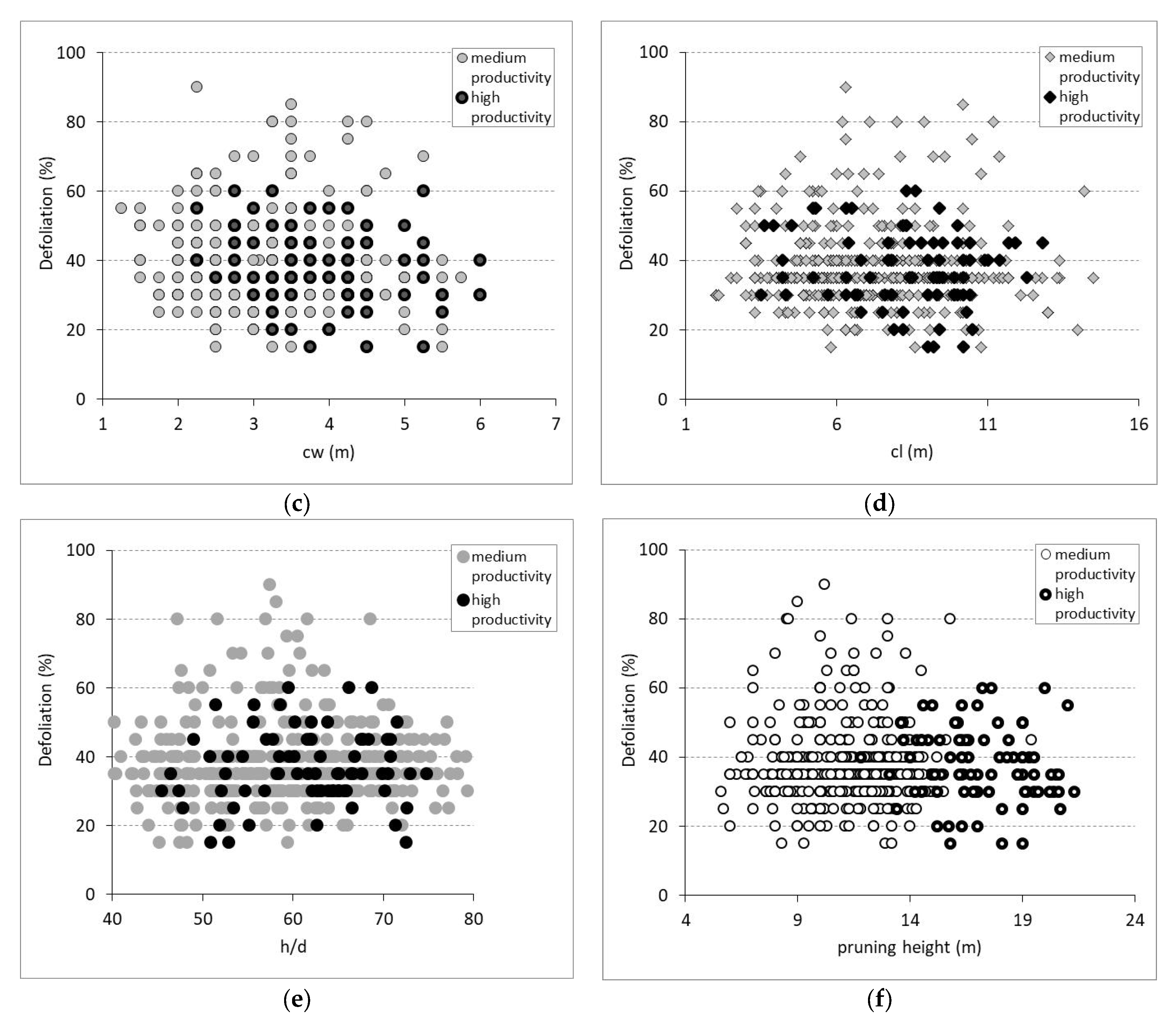
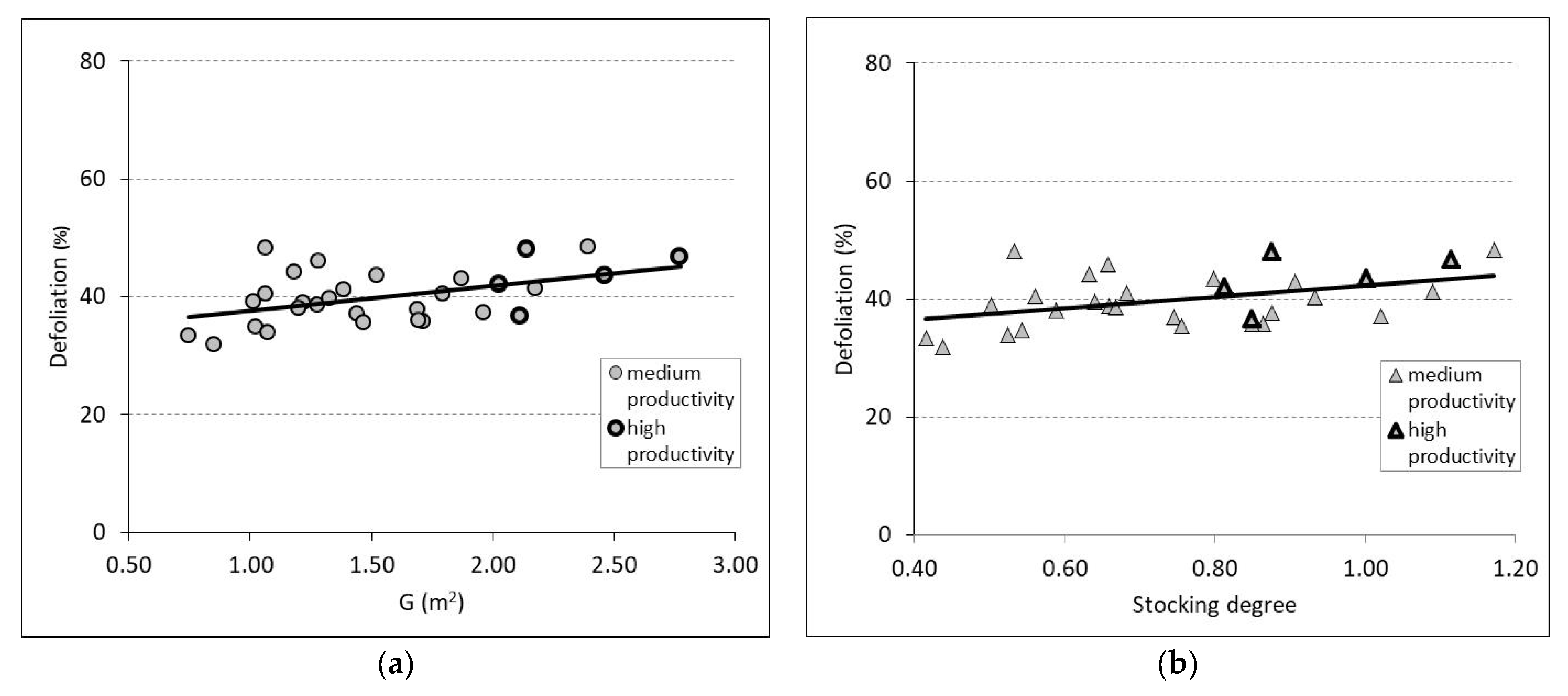
3.2. Defoliation and Biometric Parameters in Trees and Stands
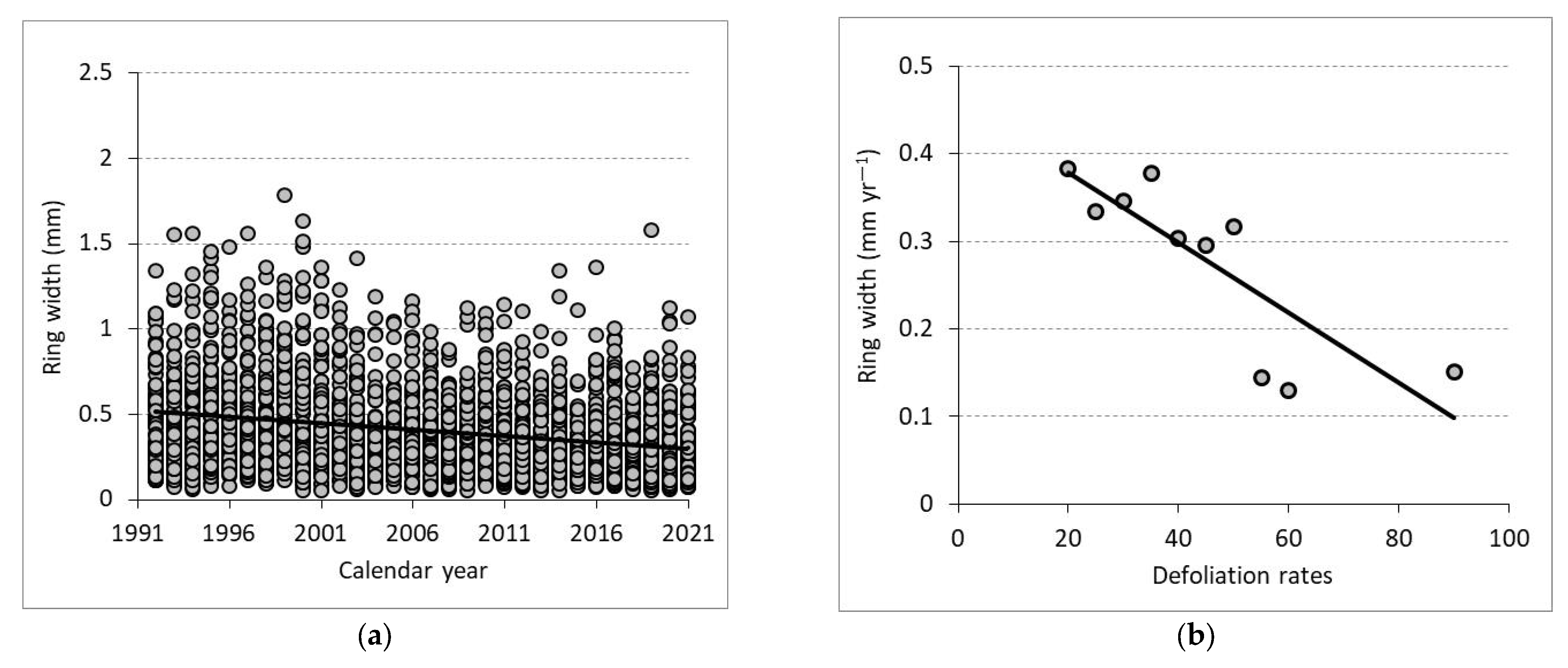
3.3. Radial Growth and Crown Defoliation
3.4. Radial Growth and Climatic Conditions
4. Discussion
4.1. Crown Defoliation
4.2. Relationship between Radial Growth and Defoliation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, M.; Antwi-Agyei, P.; Aragon-Durand, F.; Babiker, M.; Bertoldi, P.; Bind, M.; Brown, S.; Buckeridge, M.; Camilloni, I.; Cartwright, A. Technical Summary: Global Warming of 1.5 C. An IPCC Special Report on the Impacts of Global Warming of 1.5 C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; IPCC—The Intergovernamental Panel on Climate Change: Geneva, Switzerland, 2019. [Google Scholar]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries: Natural disturbances in the european forests. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Guillén Bolaños, T.; Bindi, M.; Brown, S.; Camilloni, I.A.; Diedhiou, A.; Djalante, R.; Ebi, K. The human imperative of stabilizing global climate change at 1.5 C. Science 2019, 365, 6459. [Google Scholar] [CrossRef] [PubMed]
- Ficko, A.; Poljanec, A.; Boncina, A. Do changes in spatial distribution, structure and abundance of silver fir (Abies alba Mill.) indicate its decline? For. Ecol. Manag. 2011, 261, 844–854. [Google Scholar] [CrossRef]
- Čavlović, J.; Bončina, A.; Božić, M.; Goršić, E.; Simončič, T.; Teslak, K. Depression and growth recovery of silver fir in uneven-aged Dinaric forests in Croatia from 1901 to 2001. Forestry 2015, 88, 586–598. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; Mcdowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef]
- McDowell, N.G.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Eichhorn, J.; Roskams, P.; Pototic, N.; Timmerman, V.; Ferretti, M.; Mues, V.; Szepesi, A.; Durrant, D.; Seletcovic, I.; Schöck, H.W.; et al. Part IV: Visual assessment of crown condition and damaging agents. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Coordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 54. Available online: http://www.icp-forests.org/Manual.htm (accessed on 26 September 2022).
- Ferretti, M.; Ghosh, S.; Gottardini, E. Stem radial growth is negatively related to tree defoliation and damage in conifers, Northern Italy. Front. For. Glob. Chang. 2021, 4, 775600. [Google Scholar] [CrossRef]
- Sicard, P.; De Marco, A.; Dalstein–Richier, L.; Tagliaferro, F.; Renou, C.; Paoletti, E. An epidemiological assessment of stomatal ozone flux–based critical levels for visible ozone injury in Southern European forests. Sci. Total Environ. 2016, 541, 729–741. [Google Scholar] [CrossRef]
- Badea, O. Manual on Methodology for Long–Term Monitoring of Forest Ecosystems Status under Air Pollution and Climate Change Influence; Editura Silvică: Bucharest, Romania, 2008; pp. 5–97. [Google Scholar]
- Grote, R.; Gessler, A.; Hommel, R.; Poschenrieder, W.; Priesack, E. Importance of tree height and social position for drought-related stress on tree growth and mortality. Trees 2016, 30, 1467–1482. [Google Scholar] [CrossRef]
- Mina, M.; Bugmann, H.; Cordonnier, T.; Irauschek, F.; Klopcic, M.; Pardos, M.; Cailleret, M. Future ecosystem services from European mountain forests under climate change. J. Appl. Ecol. 2017, 54, 389–401. [Google Scholar] [CrossRef]
- Rajczak, J.; Pall, P.; Schär, C. Projections of extreme precipitation events in regional climate simulations for Europe and the Alpine Region. J. Geophys. Res.-Atmos. 2013, 118, 1–17. [Google Scholar] [CrossRef]
- Sidor, C.G.; Camarero, J.J.; Popa, I.; Badea, O.; Apostol, E.N.; Vlad, R. Forest vulnerability to extreme climatic events in Romanian Scots pine forests. Sci. Total Environ. 2019, 678, 721–727. [Google Scholar] [CrossRef]
- Popa, A.; Popa, I.; Roibu, C.-C.; Badea, O.N. Do Different Tree-Ring Proxies Contain Different emperature Signals? A Case Study of Norway Spruce (Picea Abies (L.) Karst) in the Eastern Carpathians. Plants 2022, 11, 2428. [Google Scholar] [CrossRef]
- Tudoran, G.M.; Zotta, M. Adapting the planning and management of Norway spruce forests in mountain areas of Romania to environmental conditions including climate change. Sci. Total Environ. 2020, 698, 133761. [Google Scholar] [CrossRef] [PubMed]
- Tudoran, G.M.; Cicșa, A.; Ciceu, A.; Dobre, A.C. Growth Relationships in Silver Fir Stands at Their Lower-Altitude Limit in Romania. Forests 2021, 12, 439. [Google Scholar] [CrossRef]
- Tudoran, G.M.; Cicșa, A.; Ciceu, A.; Dobre, A.C. Management of Recreational Forests in the Romanian Carpathians. Forests 2022, 13, 1369. [Google Scholar] [CrossRef]
- Cicşa, A.; Tudoran, G.M.; Boroeanu, M.; Dobre, A.C.; Spârchez, G. Effect of Species Composition on Growth and Yield in Mixed Beech–Coniferous Stands. Forests 2022, 13, 1651. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernandez-Cancio, A. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Hereş, A.M.; Polanco-Martínez, J.M.; Petritan, I.C.; Petritan, A.M.; Jorge Curiel Yuste, J. The stationary and non-stationary character of the silver fir, black pine and Scots pine tree-growth-climate relationships. Agric. For. Meteorol. 2022, 325, 109146. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian Mountains. Trees 2009, 23, 95–106. [Google Scholar] [CrossRef]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Schurman, J.S.; Babst, F.; Björklund, J.; Rydval, M.; Bače, R.; Čada, V.; Janda, P.; Mikolas, M.; Saulnier, M.; Trotsiuk, V.; et al. The climatic drivers of primary Picea forest growth along the Carpathian arc are changing under rising temperatures. Glob. Chang. Biol. 2019, 25, 3136–3150. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Bacaro, G.; Brunialti, G.; Calderisi, M.; Croisé, L.; Frati, L.; Nicolas, M. Tree canopy defoliation can reveal growth decline in mid-latitude temperate forests. Ecol. Indic. 2021, 127, 107749. [Google Scholar] [CrossRef]
- Doniţǎ, N.; Popescu, A.; Paucǎ-Comǎnescu, M.; Mihǎilescu, S.; Biriş, I.A. Habitatele din România; Editura Tehnicǎ Silvica: Bucureşti, Romania, 2005; pp. 254–255. ISSN 973-96001-4-X. [Google Scholar]
- Silvestru-Grigore, C.V.; Dinulică, F.; Spârchez, G.; Hălălișan, A.F.; Dincă, L.C.; Enescu, R.E.; Crișan, V.E. Radial Growth Behavior of Pines on Romanian Degraded Lands. Forests 2018, 9, 213. [Google Scholar] [CrossRef]
- Şofletea, N.; Curtu, A.L. Dendrologie; Editura Universitǎţii Transilvania: Brașov, Romania, 2007; pp. 64–67. [Google Scholar]
- Hereş, A.M.; Petritan, I.C.; Bigler, C.; Curtu, A.L.; Petrea, Ş.; Petritan, A.M.; Polanco-Martínez, J.M.; Rigling, A.; Curiel Yuste, J. Legacies of past forest management determine current responses to severe drought events of conifer species in the Romanian Carpathians. Sci. Total Environ. 2021, 751, 141851. [Google Scholar] [CrossRef]
- Przybylski, P.; Mohytych, V.; Rutkowski, P.; Tereba, A.; Tyburski, Ł.; Fyalkowska, K. Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests. Sustainability 2021, 13, 1239. [Google Scholar] [CrossRef]
- Przybylski, P.; Jastrzȩbowski, S.; Ukalski, K.; Tyburski, L.; Konatowska, M. Quantitative and qualitative assessment of pine seedlings under controlled undergrowth disturbance: Fire and soil scarification. Front. For. Glob. Chang. 2022, 5, 225. [Google Scholar] [CrossRef]
- Pollastrini, M.; Puletti, N.; Selvi, F.; Iacopetti, G.; Bussotti, F. Widespread crown defoliation after a drought and heat wave in the forests of Tuscany (Central Italy) and their recovery—A case study from summer 2017. Front. For. Glob. Chang. 2019, 2, 74. [Google Scholar] [CrossRef]
- Ferretti, M.; Nicolas, M.; Bacaro, G.; Brunialti, G.; Calderisi, M.; Croisé, L.; Frati, L.; Lanier, M.; Maccherini, S.; Santi, E.; et al. Plot-scale modelling to detect size, extent, and correlates of changes in tree defoliation in French high forests. For. Ecol. Manag. 2014, 311, 56–69. [Google Scholar] [CrossRef]
- Rohner, B.; Kumar, S.; Liechti, K.; Gessler, A.; Ferretti, M. Tree Vitality Indicators Revealed a Rapid Response of Beech Forests to the 2018 Drought. Ecol. Indic. 2021, 120, 106903. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Tudoran, G.M. Using mathematical models based on unmanned aerial vehicle optical imagery to estimate tree and stand characteristics. Bull. Transilv. Univ. Braşov. 2022, 15, 1. [Google Scholar] [CrossRef]
- Giurgiu, V.; Drăghiciu, D. Modele Matematico-Auxologice Și Tabele de Producție Pentru Arborete; Editura Ceres: București, Romania, 2004; pp. 132–136. [Google Scholar]
- Timmermann, V.; Potočić, N.; Ognjenović, M.; Kirchner, T. Part B: Tree crown condition in 2021. In Forest Condition in Europe: The 2022 Assessment. ICP Forests Technical Report under the UNECE Convention on Long-Range Transboundary Air Pollution (Air Convention); Thünen Institute: Eberswalde, Germany, 2022. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program cofecha. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Giurgiu, V.; Decei, I.; Drăghiciu, D. Metode Şi Tabele Dendrometrice; Editura Ceres: București, Romania, 2004; pp. 53–54. [Google Scholar]
- Tudoran, G.-M.; Cicșa, A.; Boroeanu, M.; Dobre, A.-C.; Pascu, I.-S. Forest Dynamics after Five Decades of Management in the Romanian Carpathians. Forests 2021, 12, 783. [Google Scholar] [CrossRef]
- Available online: https://www.meteoblue.com/ro/vreme/archive/windrose/bra%C8%99ov_rom%C3%A2nia_683844 (accessed on 17 March 2022).
- Badea, O.; Tanase, M.; Georgeta, J.; Anisoara, L.; Peiov, A.; Uhlirova, H.; Pajtikc, J.; Wawrzoniakd, J.; Shparyk, Y. Forest health status in the Carpathian Mountains over the period 1997–2001. Environ. Pollut. 2004, 130, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Bytnerowitz, A.; Badea, O.; Popescu, F.; Musselman, R.; Tanase, M.; Barbu, I.; Fraczek, W.; Gembasu, N.; Surdu, A.; Danescu, F.; et al. Air pollution, precipitation chemistry and forest health in the Retezat Mountains, Southern Carpathians, Romania. Environ. Pollut. 2005, 137, 546–567. [Google Scholar] [CrossRef]
- Candel-Pérez, D.; Lucas-Borja, M.E.; García-Cervigón, A.I.; Tíscar, P.A.; Andivia, E.A.; Bose, K.; Sánchez-Salguero, R.; Camarero, J.J.; Linares, J.C. Forest structure drives the expected growth of Pinus nigra along its latitudinal gradient under warming climate. For. Ecol. Manag. 2022, 505, 119818, ISSN 0378-1127. [Google Scholar] [CrossRef]
- Móricz, N.; Garamszegi, B.; Rasztovits, E.; Bidló, A.; Horváth, A.; Jagicza, A.; Illés, G.; Vekerdy, Z.; Somogyi, Z.; Gálos, B. Recent Drought-Induced Vitality Decline of Black Pine (Pinus nigra Arn.) in South-West Hungary—Is This Drought-Resistant Species under Threat by Climate Change? Forests 2018, 9, 414. [Google Scholar] [CrossRef]
- Eickenscheidt, N.; Augustin, N.H.; Wellbrock, N. Spatio-temporal modelling of forest monitoring data: Modelling German tree defoliation data collected between 1989 and 2015 for trend estimation and survey grid examination using GAMMs. iForest 2019, 12, 338–348. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Bose, A.K.; Andivia, E.; Candel-Pérez, D.; Plaza-Álvarez, P.A.; Linares, J.C. Assessing Tree Drought Resistance and Climate-Growth Relationships under Different Tree Age Classes in a Pinus nigra Arn ssp. Salzmannii Forests. Forests 2021, 12, 1161. [Google Scholar] [CrossRef]
- Marchin, R.M.; Esperon-Rodriguez, M.; Tjoelker, M.G.; Ellsworth, D.S. Crown dieback and mortality of urban trees linked to heatwaves during extreme drought. Sci. Total Environ. 2022, 850, 157915. [Google Scholar] [CrossRef] [PubMed]
- Bond, B.J. Age-related changes in photosynthesis of woody plants. Trends Plant Sci. 2000, 5, 349–353. [Google Scholar] [CrossRef]
- Koch, G.; Sillett, S.; Jennings, G.; Davis, S. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Jacquet, J.S.; Orazio, C.; Jactel, H. Defoliation by processionary moth significantly reduces tree growth: A quantitative review. Ann. For. Sci. 2012, 69, 857–866. [Google Scholar] [CrossRef]
- Westerband, A.C.; Kagawa-Viviani, A.K.; Bogner, K.K.; Beilman, D.; Knight, M.; Barton, K. Seedling drought tolerance and functional traits vary in response to the timing of water availability in a keystone Hawaiian tree species. Plant Ecol. 2019, 220, 321–344. [Google Scholar] [CrossRef]
- Tudoran, A.; Bylund, H.; Nordlander, G.; Oltean, I.; Puentes, A. Using associational effects of European beech on Norway spruce to mitigate damage by a forest regeneration pest, the pine weevil Hylobius abietis. For. Ecol. Manag. 2021, 486, 118980, ISSN 0378-1127. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SP | Inventoried Area (ha) | Species | N | dg ± SD (cm) | hg ± SD (m) | cwg ± SD (m) | clg ± SD (m) |
|---|---|---|---|---|---|---|---|
| SP1 | 0.25 | black pine | 71 | 34.98 ± 8.18 | 19.3 ± 3.3 | 3.3 ± 0.8 | 7.3 ± 2.2 |
| SP2 | 0.25 | black pine | 62 | 36.77 ± 7.9 | 18.2 ± 3.1 | 4.0 ± 0.9 | 8.0 ± 2.7 |
| SP3 | 0.25 | black pine | 61 | 36.66 ± 7.7 | 19.1 ± 3.6 | 3.7 ± 0.8 | 8.0 ± 2.5 |
| SP4 | 0.25 | black pine | 90 | 40.37 ± 6.6 | 25.3 ± 3.0 | 3.9 ± 0.9 | 7.8 ± 2.6 |
| SP5 | 0.25 | black pine | 116 | 29.73 ± 4.7 | 19.4 ± 2.5 | 3.0 ± 0.5 | 6.6 ± 2.2 |
| SP6 | 0.25 | black pine | 108 | 29.92 ± 6.1 | 18.5 ± 2.3 | 2.7 ± 0.6 | 6.2 ± 1.9 |
| Total | 1.50 | –– | 508 | –– | –– | –– | –– |
| Biometric Parameter | n | Intercept | Slope (Value/std. Error/t) | R2 | p-Value | MAE | RMSE |
|---|---|---|---|---|---|---|---|
| dg | 30 | 21.847 | 0.524 | 0.249 | 0.005 | −0.0016 | 3.9207 |
| 6.008 | 0.172 | ||||||
| 3.636 | 3.043 | ||||||
| V | 30 | 34.852 | 0.339 | 0.242 | 0.006 | −0.0026 | 3.9412 |
| 1.873 | 0.113 | ||||||
| 18.604 | 2.99 | ||||||
| G | 30 | 33.25 | 4.302 | 0.232 | 0.007 | 0.0005 | 3.9662 |
| 2.433 | 1.477 | ||||||
| 13.664 | 2.912 | ||||||
| SSD | 30 | 32.574 | 9.797 | 0.19 | 0.016 | 0.0002 | 4.0744 |
| 2.991 | 3.818 | ||||||
| 10.89 | 2.566 | ||||||
| clg | 30 | 27.827 | 1.663 | 0.182 | 0.019 | 0.0003 | 4.0937 |
| 4.931 | 0.666 | ||||||
| 5.643 | 2.498 | ||||||
| cwg | 30 | 27.3 | 3.732 | 0.179 | 0.02 | 0.001 | 4.1012 |
| 5.193 | 1.51 | ||||||
| 5.27 | 2.472 | ||||||
| hg | 30 | 26.053 | 0.698 | 0.152 | 0.033 | 0.0087 | 4.175 |
| 6.266 | 0.311 | ||||||
| 4.158 | 2.242 |
| Equation (RW) | Coefficient | Std. Error (SE) | t | p | R2 | MAE | RMSE |
|---|---|---|---|---|---|---|---|
| Intercept | 15.256 | 1.1961 | 7.78 | 0.000 | 0.67 | −0.0025 | 0.0379 |
| Slope | –0.0074 | 0.001 | −7.577 |
| Equation (DEF) | Coefficient | SE | t | p | R2 | MAE | RMSE |
|---|---|---|---|---|---|---|---|
| Intercept | −263.692 | 15.452 | −17.065 | 0.003 | 0.99 | 0.0028 | 0.8194 |
| Slope | 38.858 | 2 | 19.425 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tudoran, G.-M.; Cicșa, A.; Dobre, A.-C.; Cicșa, M.; Pascu, I.-S.; Leca, Ş. Health and Growth of Black Pine outside Its Natural Distribution Range in the Romanian Carpathians. Forests 2023, 14, 884. https://doi.org/10.3390/f14050884
Tudoran G-M, Cicșa A, Dobre A-C, Cicșa M, Pascu I-S, Leca Ş. Health and Growth of Black Pine outside Its Natural Distribution Range in the Romanian Carpathians. Forests. 2023; 14(5):884. https://doi.org/10.3390/f14050884
Chicago/Turabian StyleTudoran, Gheorghe-Marian, Avram Cicșa, Alexandru-Claudiu Dobre, Maria Cicșa, Ionuț-Silviu Pascu, and Ştefan Leca. 2023. "Health and Growth of Black Pine outside Its Natural Distribution Range in the Romanian Carpathians" Forests 14, no. 5: 884. https://doi.org/10.3390/f14050884