
Responses of Soil Microbial Diversity to Forest Management Practices after Pine Wilt Disease Infection

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Methods
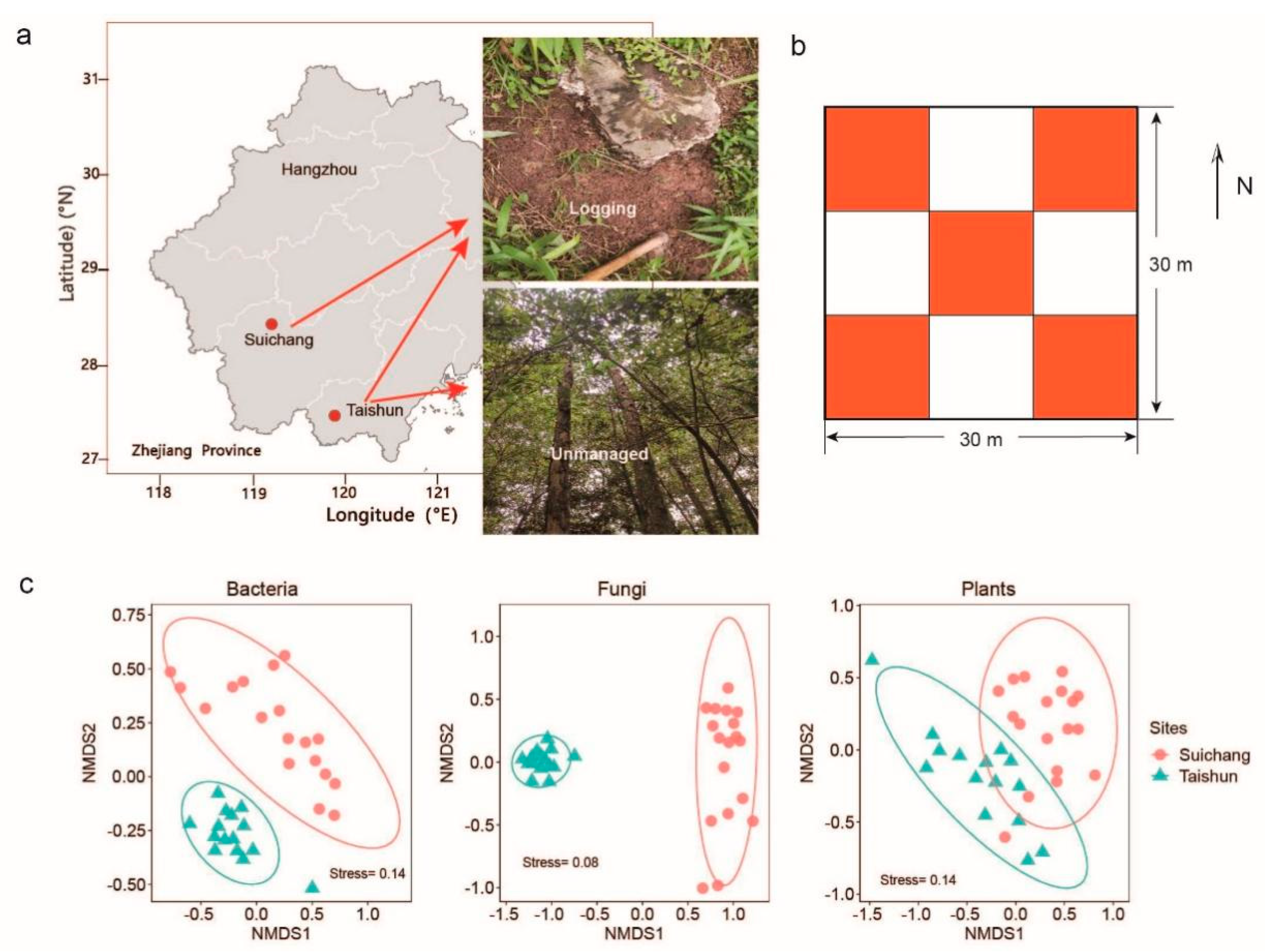
2.1. Study Area
2.2. Soil Sampling
2.3. DNA Extraction and Sequencing
2.4. Bioinformatics
2.5. Statistical Analyses
3. Results
3.1. General Characterization of Soil Microbial Communities
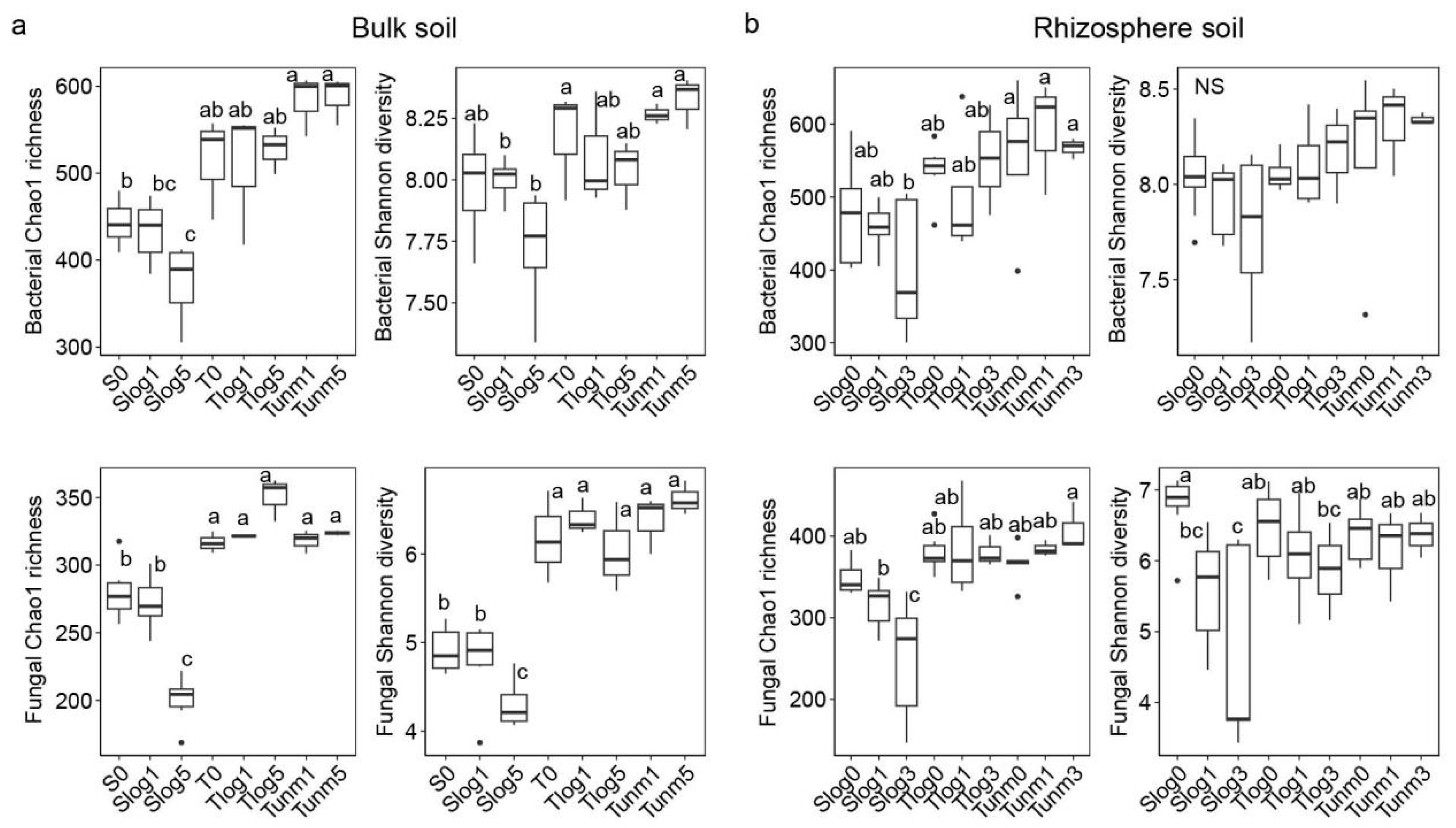
3.2. Variations in Microbial Community Alpha Diversity
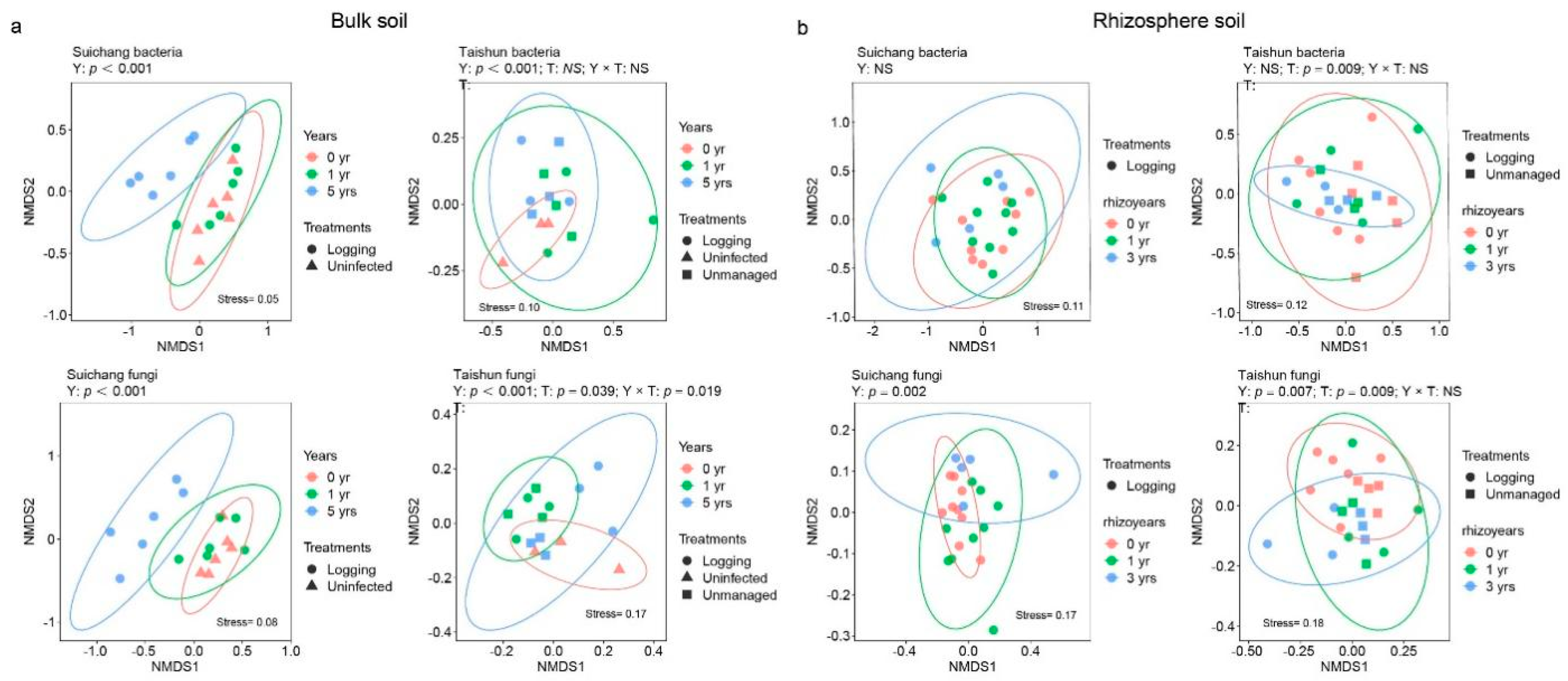
3.3. Variations in Microbial Community Composition
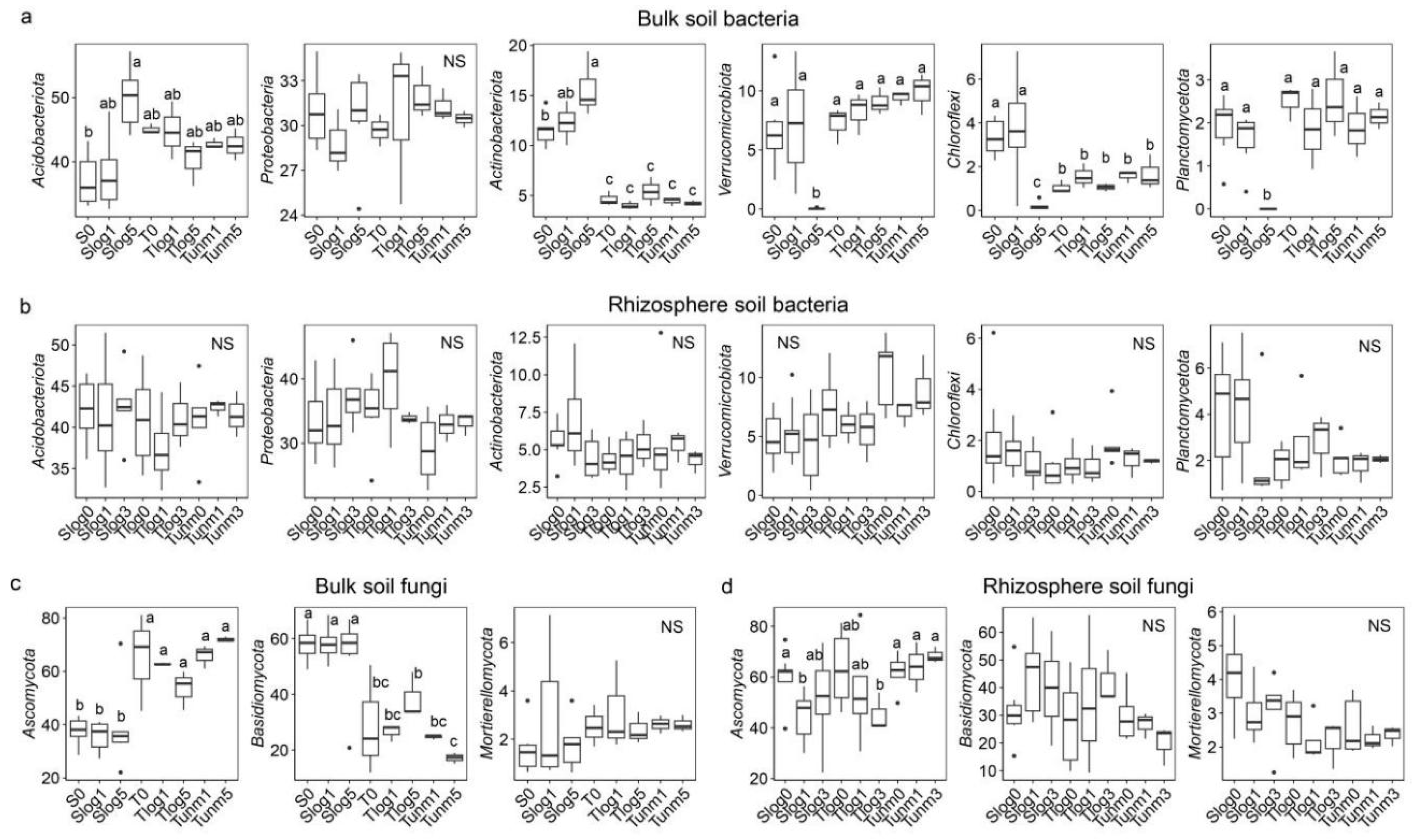
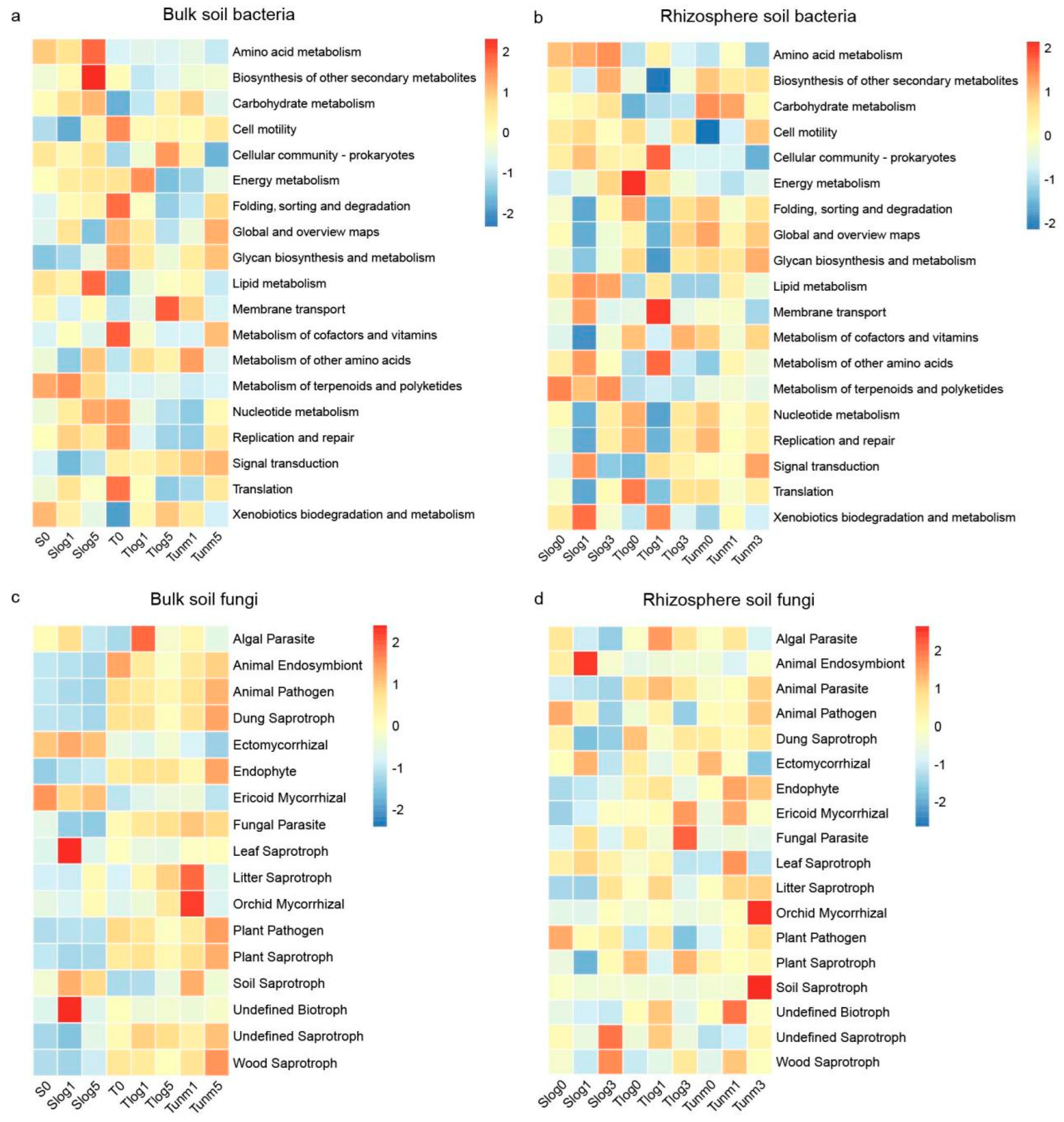
3.4. Variations in Microbial Community Functional Groups
4. Discussion
4.1. Decrease in Soil Bacterial and Fungal Diversity to PWD
4.2. Response of Microbial Functional Groups to PWD
4.3. Comparison between Logging and Unmanaged Practice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, G.; Wang, K.; Liu, X.; Peng, S. Simulation of the Biomass Dynamics of Masson Pine Forest under Different Management. J. For. Res. 2006, 17, 305–311. [Google Scholar] [CrossRef]
- Tóth, Á. Bursaphelenchus xylophilus, the Pinewood Nematode: Its Significance and a Historical Review. Acta Biol. Szeged. 2011, 55, 213–217. [Google Scholar] [CrossRef]
- Yu, M.; Xu, X.; Ding, P. Economic Loss versus Ecological Gain: The Outbreaks of Invaded Pinewood Nematode in China. Biol. Invasions 2011, 13, 1283–1290. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests 2023, 14, 174. [Google Scholar] [CrossRef]
- Zhao, L.; Jiang, P.; Humble, L.M.; Sun, J. Within-Tree Distribution and Attractant Sampling of Propagative Pinewood Nematode, Bursaphelenchus xylophilus: An Early Diagnosis Approach. For. Ecol. Manag. 2009, 258, 1932–1937. [Google Scholar] [CrossRef]
- Fukasawa, Y.; Matsuoka, S. Communities of Wood-Inhabiting Fungi in Dead Pine Logs along a Geographical Gradient in Japan. Fungal Ecol. 2015, 18, 75–82. [Google Scholar] [CrossRef]
- Chen, J.; Xu, H.; He, D.; Li, Y.; Luo, T.; Yang, H.; Lin, M. Historical Logging Alters Soil Fungal Community Composition and Network in a Tropical Rainforest. For. Ecol. Manag. 2019, 433, 228–239. [Google Scholar] [CrossRef]
- Qu, Z.; Braima, A.; Liu, B.; Ma, Y.; Sun, H. Soil Fungal Community Structure and Function Shift during a Disease-Driven Forest Succession. Microbiol. Spectr. 2022, 10, e0079522. [Google Scholar] [CrossRef] [PubMed]
- Pennanen, T.; Liski, J.; Bååth, E.; Kitunen, V.; Uotila, J.; Westman, C.J.; Fritze, H. Structure of the Microbial Communities in Coniferous Forest Soils in Relation to Site Fertility and Stand Development Stage. Microb. Ecol. 1999, 38, 168–179. [Google Scholar] [CrossRef]
- Talbot, J.M.; Allison, S.D.; Treseder, K.K. Decomposers in Disguise: Mycorrhizal Fungi as Regulators of Soil C Dynamics in Ecosystems under Global Change. Funct. Ecol. 2008, 22, 955–963. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The Unseen Majority: Soil Microbes as Drivers of Plant Diversity and Productivity in Terrestrial Ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Dang, P.; Gao, Y.; Liu, J.; Yu, S.; Zhao, Z. Effects of Thinning Intensity on Understory Vegetation and Soil Microbial Communities of a Mature Chinese Pine Plantation in the Loess Plateau. Sci. Total Environ. 2018, 630, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Ushio, M.; Wagai, R.; Balser, T.C.; Kitayama, K. Variations in the Soil Microbial Community Composition of a Tropical Montane Forest Ecosystem: Does Tree Species Matter? Soil Biol. Biochem. 2008, 40, 2699–2702. [Google Scholar] [CrossRef]
- Chu, H.; Wang, C.; Wang, H.; Chen, H.; Tang, M. Pine Wilt Disease Alters Soil Properties and Root-Associated Fungal Communities in Pinus tabulaeformis Forest. Plant Soil 2016, 404, 237–249. [Google Scholar] [CrossRef]
- Kim, C.; Jang, K.S.; Kim, J.B.; Byun, J.K.; Lee, C.H.; Jeon, K.S. Relationship between Soil Properties and Incidence of Pine Wilt Disease at Stand Level. Landsc. Ecol. Eng. 2010, 6, 119–124. [Google Scholar] [CrossRef]
- Lloret, F.; Mattana, S.; Curiel Yuste, J. Climate-Induced Die-off Affects Plant–Soil–Microbe Ecological Relationship and Functioning. FEMS Microbiol. Ecol. 2015, 91, 1–12. [Google Scholar] [CrossRef]
- Jin, X.; Liu, Y.; Hu, W.; Wang, G.; Kong, Z.; Wu, L.; Ge, G. Soil Bacterial and Fungal Communities and the Associated Nutrient Cycling Responses to Forest Conversion after Selective Logging in a Subtropical Forest of China. For. Ecol. Manag. 2019, 444, 308–317. [Google Scholar] [CrossRef]
- Veselá, P.; Vašutová, M.; Edwards-Jonášová, M.; Cudlín, P. Soil Fungal Community in Norway Spruce Forests under Bark Beetle Attack. Forests 2019, 10, 109. [Google Scholar] [CrossRef]
- Rodriguez-Ramos, J.C.; Cale, J.A.; Cahill, J.F.; Simard, S.W.; Karst, J.; Erbilgin, N. Changes in Soil Fungal Community Composition Depend on Functional Group and Forest Disturbance Type. New Phytol. 2021, 229, 1105–1117. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Domínguez-Begines, J.; Villa-Sanabria, E.; García, L.V.; Muñoz-Pajares, A.J. Tree Decline and Mortality Following Pathogen Invasion Alters the Diversity, Composition and Network Structure of the Soil Microbiome. Soil Biol. Biochem. 2022, 166, 108560. [Google Scholar] [CrossRef]
- Addison, S.L.; Smaill, S.J.; Garrett, L.G.; Wakelin, S.A. Effects of Forest Harvest and Fertiliser Amendment on Soil Biodiversity and Function Can Persist for Decades. Soil Biol. Biochem. 2019, 135, 194–205. [Google Scholar] [CrossRef]
- Song, P.; Ren, H.; Jia, Q.; Guo, J.; Zhang, N.; Ma, K. Effects of Historical Logging on Soil Microbial Communities in a Subtropical Forest in Southern China. Plant Soil 2015, 397, 115–126. [Google Scholar] [CrossRef]
- Bini, D.; dos Santos, C.A.; do Carmo, K.B.; Kishino, N.; Andrade, G.; Zangaro, W.; Nogueira, M.A. Effects of Land Use on Soil Organic Carbon and Microbial Processes Associated with Soil Health in Southern Brazil. Eur. J. Soil Biol. 2013, 55, 117–123. [Google Scholar] [CrossRef]
- Chatterjee, A.; Vance, G.F.; Pendall, E.; Stahl, P.D. Timber Harvesting Alters Soil Carbon Mineralization and Microbial Community Structure in Coniferous Forests. Soil Biol. Biochem. 2008, 40, 1901–1907. [Google Scholar] [CrossRef]
- Hartmann, M.; Howes, C.G.; Vaninsberghe, D.; Yu, H.; Bachar, D.; Christen, R.; Henrik Nilsson, R.; Hallam, S.J.; Mohn, W.W. Significant and Persistent Impact of Timber Harvesting on Soil Microbial Communities in Northern Coniferous Forests. ISME J. 2012, 6, 2199–2218. [Google Scholar] [CrossRef]
- Reva, V.; Fonseca, L.; Lousada, J.L.; Abrantes, I.; Viegas, D.X. Impact of the Pinewood Nematode, Bursaphelenchus xylophilus, on Gross Calorific Value and Chemical Composition of Pinus pinaster Woody Biomass. Eur. J. For. Res. 2012, 131, 1025–1033. [Google Scholar] [CrossRef]
- Dannenmann, M.; Simon, J.; Gasche, R.; Holst, J.; Naumann, P.S.; Kögel-Knabner, I.; Knicker, H.; Mayer, H.; Schloter, M.; Pena, R.; et al. Tree Girdling Provides Insight on the Role of Labile Carbon in Nitrogen Partitioning between Soil Microorganisms and Adult European Beech. Soil Biol. Biochem. 2009, 41, 1622–1631. [Google Scholar] [CrossRef]
- Deng, J.; Yu, D.; Zhou, W.; Zhou, L.; Zhu, W. Variations of Phyllosphere and Rhizosphere Microbial Communities of Pinus koraiensis Infected by Bursaphelenchus xylophilus. Microb. Ecol. 2022, 84, 285–301. [Google Scholar] [CrossRef]
- Han, G.; Mannaa, M.; Kim, N.; Jeon, H.W.; Jung, H.; Lee, H.H.; Kim, J.; Park, J.; Park, A.R.; Kim, J.C.; et al. Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease. Microorganisms 2021, 9, 688. [Google Scholar] [CrossRef]
- Hong, C.; Si, Y.; Xing, Y.; Li, Y. Illumina MiSeq Sequencing Investigation on the Contrasting Soil Bacterial Community Structures in Different Iron Mining Areas. Environ. Sci. Pollut. Res. 2015, 22, 10788–10799. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fang, L.; Beiyuan, J.; Cui, Y.; Peng, Q.; Zhu, S.; Wang, M.; Zhang, X. Improvement of Alfalfa Resistance against Cd Stress through Rhizobia and Arbuscular Mycorrhiza Fungi Co-Inoculation in Cd-Contaminated Soil. Environ. Pollut. 2021, 277, 116758. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A.; Bonfante, P.; Bianciotto, V. Unravelling Soil Fungal Communities from Different Mediterranean Land-Use Backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Martin, M. Cut Adapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Author Correction: Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a Unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 10 June 2022).
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A Review of the Roles of Forest Canopy Gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Li, S.; Huang, X.; Lang, X.; Xu, F.; Li, H.; Zheng, M.; Su, J. Effect of Selective Logging on Soil Microbial Communities in a Pinus yunnanensis Forest. Land Degrad. Dev. 2020, 31, 2268–2280. [Google Scholar] [CrossRef]
- Qu, Z.L.; Liu, B.; Ma, Y.; Xu, J.; Sun, H. The Response of the Soil Bacterial Community and Function to Forest Succession Caused by Forest Disease. Funct. Ecol. 2020, 34, 2548–2559. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree Species Influence on Microbial Communities in Litter and Soil: Current Knowledge and Research Needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of Fungal and Bacterial Communities in Forest Litter and Soil Is Largely Determined by Dominant Trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef]
- Jones, D.L.; Hodge, A.; Kuzyakov, Y. Plant and Mycorrhizal Regulation of Rhizodeposition. New Phytol. 2004, 163, 459–480. [Google Scholar] [CrossRef]
- Ma, Y.; Qu, Z.L.; Liu, B.; Tan, J.J.; Asiegbu, F.O.; Sun, H. Bacterial Community Structure of Pinus thunbergii Naturally Infected by the Nematode Bursaphelenchus xylophilus. Microorganisms 2020, 8, 307. [Google Scholar] [CrossRef]
- Xiong, Q.; Li, L.; Luo, X.; He, X.; Zhang, L.; Pan, K.; Liu, C.; Sun, H. Driving Forces for Recovery of Forest Vegetation after Harvesting a Subalpine Oak Forest in Eastern Tibetan Plateau. Environ. Sci. Pollut. Res. 2021, 28, 67748–67763. [Google Scholar] [CrossRef]
- Malik, A.A.; Swenson, T.; Weihe, C.; Morrison, E.W.; Martiny, J.B.H.; Brodie, E.L.; Northen, T.R.; Allison, S.D. Drought and Plant Litter Chemistry Alter Microbial Gene Expression and Metabolite Production. ISME J. 2020, 14, 2236–2247. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, X.; Su, Y.; Li, X.; Yin, H.; Li, X.; Guo, M.; He, Y. Forest Gaps Influence Fungal Community Assembly in a Weeping Cypress Forest. Appl. Microbiol. Biotechnol. 2019, 103, 3215–3224. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PWD Infected Years | Suichang Logging | Taishun Logging | Taishun Unmanaged |
|---|---|---|---|
| PWD-infected plots | |||
| Uninfected (0 year) | 6 | 3 | |
| 1 year | 6 | 3 | 3 |
| 5 years | 6 | 3 | 3 |
| PWD-infected pine individuals | |||
| Uninfected (0 year) | 8 | 6 | 6 |
| 1 year | 13 | 5 | 6 |
| 3 years | 6 | 5 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Gong, X.; Yu, S.; Wei, B.; Chu, L.; Liu, J.; He, X.; Yu, M. Responses of Soil Microbial Diversity to Forest Management Practices after Pine Wilt Disease Infection. Forests 2023, 14, 862. https://doi.org/10.3390/f14050862
Guo J, Gong X, Yu S, Wei B, Chu L, Liu J, He X, Yu M. Responses of Soil Microbial Diversity to Forest Management Practices after Pine Wilt Disease Infection. Forests. 2023; 14(5):862. https://doi.org/10.3390/f14050862
Chicago/Turabian StyleGuo, Jing, Xiaofei Gong, Shuisheng Yu, Boliang Wei, Liying Chu, Jinliang Liu, Xiaoyong He, and Mingjian Yu. 2023. "Responses of Soil Microbial Diversity to Forest Management Practices after Pine Wilt Disease Infection" Forests 14, no. 5: 862. https://doi.org/10.3390/f14050862