
Effects of Acid Rain Stress on the Physiological and Biochemical Characteristics of Three Plant Species
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Materials
2.3. Experimental Design
2.3.1. Acid Rain Solutions
2.3.2. Spray Application
2.4. Sample Collection and Indices
2.4.1. Determination of Photosynthetic Physiological Indicators
2.4.2. Determination of Biochemical Indicators
2.5. Data Analysis
3. Results
3.1. Effect of Acid Rain Stress on Photosynthetic Physiological Characteristics of Plants
3.2. Effects of Acid Rain Stress on Plant Biochemical Characteristics
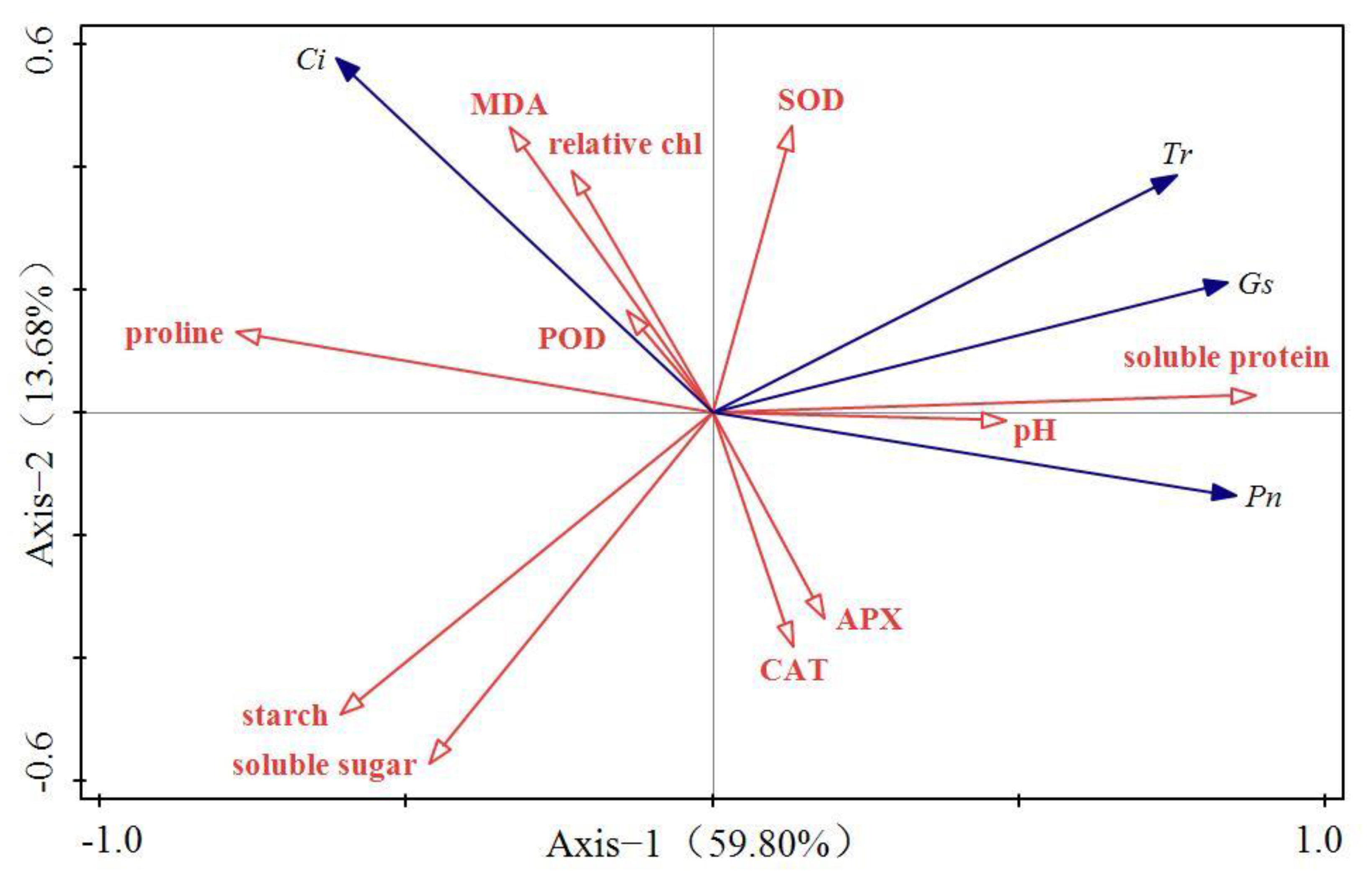
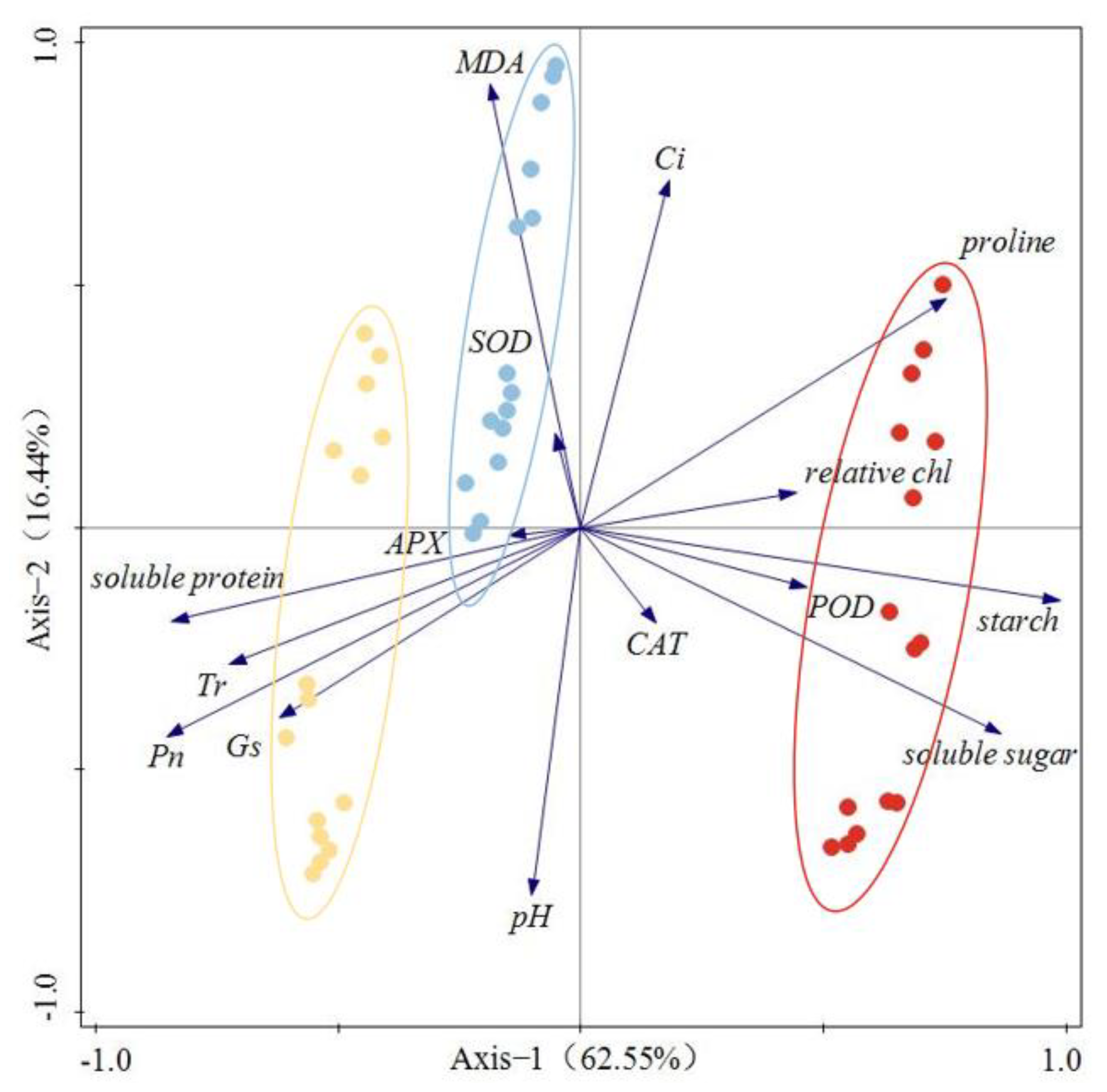
3.3. RDA Results of Photosynthetic and Biochemical Indices of Plant Seedlings under Acid Rain Stress
4. Discussion
4.1. Effects of Acid Rain Stress on Photosynthetic Physiological Characteristics of Three Plant Species
4.2. Effects of Acid Rain Stress on Biochemical Characteristics of the Tree Species
4.3. Relationship between Photosynthetic and Biochemical Characteristics of Plant Seedlings under Acid Rain Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yu, H.; He, N.; Wang, Q.; Zhu, J.; Gao, Y.; Zhang, Y.; Jia, Y.; Yu, G. Development of atmospheric acid deposition in China from the 1990s to the 2010s. Environ. Pollut. 2017, 231, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, J.; Mulder, J.; Wang, Q.; Liu, C.; He, N. High environmental costs behind rapid economic development: Evidence from economic loss caused by atmospheric acid deposition. J. Environ. Manag. 2023, 334, 117511. [Google Scholar] [CrossRef] [PubMed]
- Tong, D.; Cheng, J.; Liu, Y.; Yu, S.; Yan, L.; Hong, C.; Qin, Y.; Zhao, H.; Zheng, Y.; Geng, G.; et al. Dynamic projection of anthropogenic emissions in China: Methodology and 2015–2050 emission pathways under a range of socio-economic, climate policy, and pollution control scenarios. Atmos. Chem. Phys. 2020, 20, 5729–5757. [Google Scholar] [CrossRef]
- Ministry of Ecology and Environment of the People’s Republic of China. Report on the State of the Ecological Environment in China. [EB/OL]; Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 2018.
- Zhang, Q.; Zhu, J.; Wang, Q.; Xu, L.; Li, M.; Dai, G.; Mulder, J.; Xi, Y.; He, N. Soil acidification in China’s forests due to atmospheric acid deposition from 1980 to 2050. Sci. Bull. 2022, 67, 914–917. [Google Scholar] [CrossRef]
- Chang, M. SO2 Pollution Control: Successful and Transferable Practices (Chongqing, China); Institute for Global Environmental Strategies: Kanagawa, Japan, 2003. [Google Scholar]
- Acid Deposition Monitoring Network in East Asia. 2021. Available online: https://monitoring.eanet.asia/document/public/index (accessed on 20 February 2023).
- Galloway, J.N.; Likens, G.E.; Hawley, M.E. Acid Precipitation: Natural Versus Anthropogenic Components. Science 1984, 226, 829–831. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, X.X.; Zhang, G.L. Current status of acid rain pollution in Chongqing. Green Technol. 2018, 16, 11–14. [Google Scholar] [CrossRef]
- Whitmarsh, J.; Govindjee. The Photosynthetic Process. In Concepts in Photobiology; Springer: Dordrecht, The Netherlands, 1999; pp. 11–51. [Google Scholar] [CrossRef]
- Wen, K.; Liang, C.; Wang, L.; Hu, G.; Zhou, Q. Combined effects of lanthanumion and acid rain on growth, photosynthesis and chloroplast ultrastructure in soybean seedlings. Chemosphere 2011, 84, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, C.; Yu, T.; Dayananda, B.; Fu, B.; Senaratne, S.L.; Wu, C.; Li, J. Differential effects of acid rain on photosynthetic performance and pigment composition of the critically endangered Acer amplum subsp. catalpifolium. Glob. Ecol. Conserv. 2021, 30, e01773. [Google Scholar] [CrossRef]
- Dong, D.; Du, E.; Sun, Z.; Zeng, X.; de Vries, W. Non-linear direct effects of acid rain on leaf photosynthetic rate of terrestrial plants. Environ. Pollut. 2017, 231, 1442–1445. [Google Scholar] [CrossRef]
- Macaulay, B.M.; Enahoro, G.E. Effects of simulated acid rain on the morphology, phenology and dry biomass of a local variety of maize (Suwan-1) in Southwestern Nigeria. Environ. Monit. Assess. 2015, 187, 622. [Google Scholar] [CrossRef]
- Andrade, G.C.; Silva, L.C. Responses of tropical legumes from the Brazilian Atlantic Rainforest to simulated acid rain. Protoplasma 2016, 254, 1639–1649. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.C.; Castro, L.N.; da Silva, L.C. Micromorphological alterations induced by simulated acid rain on the leaf surface of Joannesia princeps Vell. (Euphorbiaceae). Ecol. Indic. 2020, 116, 106526. [Google Scholar] [CrossRef]
- Diatta, J.; Youssef, N.; Tylman, O.; Grzebisz, W.; Markert, B.; Drobek, L.; Wünschmann, S.; Bebek, M.; Mitko, K.; Lejwoda, P. Acid rain induced leakage of Ca, Mg, Zn, Fe from plant photosynthetic organs—Testing for deciduous and dicotyledons. Ecol. Indic. 2021, 121, 107210. [Google Scholar] [CrossRef]
- Hu, H.; Wang, L.; Liao, C.; Fan, C.; Zhou, Q.; Huang, X. Combined Effects of Lead and Acid Rain on Photosynthesis in Soybean Seedlings. Biol. Trace Element Res. 2014, 161, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, L.; Chen, M.; Wang, L.; Liang, C.; Zhou, Q.; Huang, X. Interactive effects of cadmium and acid rain on photosynthetic light reaction in soybean seedlings. Ecotoxicol. Environ. Saf. 2012, 79, 62–68. [Google Scholar] [CrossRef]
- Du, E.; Dong, D.; Zeng, X.; Sun, Z.; Jiang, X.; de Vries, W. Direct effect of acid rain on leaf chlorophyll content of terrestrial plants in China. Sci. Total. Environ. 2017, 605, 764–769. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Xiao, Z.; Lu, T.; Ren, X.; Wei, H. Effects of acid rain on plant growth: A meta-analysis. J. Environ. Manag. 2021, 297, 113213. [Google Scholar] [CrossRef]
- Ren, X.; Zhu, J.; Liu, H.; Xu, X.; Liang, C. Response of antioxidative system in rice (Oryza sativa) leaves to simulated acid rain stress. Ecotoxicol. Environ. Saf. 2018, 148, 851–856. [Google Scholar] [CrossRef]
- Ma, S.; Chen, W.; Zhang, J.; Shen, H. Influence of simulated acid rain on the physiological response of flowering Chinese cabbage and variation of soil nutrients. Plant Soil Environ. 2020, 66, 648–657. [Google Scholar] [CrossRef]
- Hua, T.; Tan, S.; Liu, Z. Effects of cadmium and simulated acid rain on growth and physiological characteristics of bermudagrass seedling. Acta Physiol. Plant. 2022, 44, 17. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Zhang, Y.; Wang, Y.; Pei, C. Response of soil chemical properties and enzyme activity of four species in the Three Gorges Reservoir area to simulated acid rain. Ecotoxicol. Environ. Saf. 2021, 208, 111457. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Y.; Zheng, Y.; Li, Y. Effects of Simulated Acid Rain on Soil Enzyme Activity and Related Chemical Indexes in Woodlands. Forests 2022, 13, 860. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Wang, Y.; Wang, B. Effects of simulated acid rain on soil respiration and its component in a mixed coniferous-broadleaved forest of the three gorges reservoir area in Southwest China. For. Ecosyst. 2019, 6, 32. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Ye, Z.P.; Yu, Q. Comparison of a new model of photosynthetic light response with the traditional model. J. Shenyang Agric. Univ. 2007, 38, 771–775. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Debnath, B.; Irshad, M.; Mitra, S.; Li, M.; Rizwan, H.M.; Liu, S.; Pan, T.; Qiu, D. Acid Rain Deposition Modulates Photosynthesis, Enzymatic and Non-enzymatic Antioxidant Activities in Tomato. Int. J. Environ. Res. 2018, 12, 203–214. [Google Scholar] [CrossRef]
- Xu, J.; Guo, L.; Liu, L. Exogenous silicon alleviates drought stress in maize by improving growth, photosynthetic and antioxidant metabolism. Environ. Exp. Bot. 2022, 201, 104974. [Google Scholar] [CrossRef]
- Sun, J.; Hu, H.; Li, Y.; Wang, L.; Zhou, Q.; Huang, X. Effects and mechanism of acid rain on plant chloroplast ATP synthase. Environ. Sci. Pollut. Res. 2016, 23, 18296–18306. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Physiological and molecular responses to magnesium nutritional imbalance in plants. Plant Soil 2013, 368, 87–99. [Google Scholar] [CrossRef]
- Velikova, V.; Tsonev, T.; Yordanov, I. Light and CO2 responses of photosynthesis and chlorophyll fluorescence characteristics in bean plants after simulated acid rain. Physiol. Plant. 1999, 107, 77–83. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, K.; Zhang, Q.; Wang, W. Ecophysiological responses of Jatropha curcas L. seedlings to simulated acid rain under different soil types. Ecotoxicol. Environ. Saf. 2019, 185, 109705. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Sanavy, S.A.M.M.; Gholamhoseini, M.; Joghan, A.K.; Majdi, M.; Kashkooli, A.B. The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol. Mol. Biol. Plants 2013, 19, 189–198. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Zhou, Q.; Huang, X. Combined effects of lanthanum (III) chloride and acid rain on photosynthetic pa-rameters in rice. Chemosphere 2014, 112, 355–361. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, T.; Ma, W.; Dayananda, B.; Iwasaki, K.; Li, J. Morphological, Physiological and Photophysiological Responses of Critically Endangered Acer catalpifolium to Acid Stress. Plants 2021, 10, 1958. [Google Scholar] [CrossRef]
- Yao, F.-F.; Ding, H.-M.; Feng, L.-L.; Chen, J.-J.; Yang, S.-Y.; Wang, X.-H. Photosynthetic and growth responses of Schima superba seedlings to sulfuric and nitric acid depositions. Environ. Sci. Pollut. Res. 2016, 23, 8644–8658. [Google Scholar] [CrossRef]
- Haines, B.L.; Jernstedt, J.A.; Neufeld, H.S. Direct foliar effects of simulated acid rain: I. Damage, growth and gas exchange. New Phytol. 1985, 99, 389–405. [Google Scholar] [CrossRef]
- Da Silva, L.C.; Oliva, M.A.; Azevedo, A.A.; Araújo, J.M.; Aguiar, R.M. Micromorphological and anatomical alterations caused by simulated acid rain in Restinga plants: Eugenia uniflora and Clusia hilariana. Water Air Soil Pollut. 2005, 168, 129–143. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, Z.; Wang, X.; Niu, J. Responses of native broadleaved woody species to elevated ozone in subtropical China. Environ. Pollut. 2012, 163, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wong, M.; Yang, J.; Ye, Z.; Jiang, P.; Zheng, S.; Zheng, S.J. Dynamics of Carbon Accumulation During the Fast Growth Period of Bamboo Plant. Bot. Rev. 2011, 77, 287–295. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Kumar, M.; Patel, M.K.; Kumar, N.; Bajpai, A.B.; Siddique, K.H.M. Metabolomics and Molecular Approaches Reveal Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 9108. [Google Scholar] [CrossRef] [PubMed]
- Moharekar, S.; Lokhande, S.; Hara, T.; Tanaka, R.; Tanaka, A.; Chavan, P. Effect of Salicylic Acid on Chlorophyll and Carotenoid Contents of Wheat and Moong Seedlings. Photosynthetica 2003, 41, 315–317. [Google Scholar] [CrossRef]
- Huang, J.; Wang, H.; Zhong, Y.; Huang, J.; Fu, X.; Wang, L.; Teng, W. Growth and physiological response of an endangered tree, Horsfieldia hainanensis merr., to simulated sulfuric and nitric acid rain in southern China. Plant Physiol. Biochem. 2019, 144, 118–126. [Google Scholar] [CrossRef]
- Shan, Y.F. Acid Rain, Atmospheric Pollution and Plants; China Environmental Science Press: Beijing, China, 1993. [Google Scholar]
- Liu, M.; Korpelainen, H.; Dong, L.; Yi, L. Physiological responses of Elaeocarpus glabripetalus seedlings exposed to simulated acid rain and cadmium. Ecotoxicol. Environ. Saf. 2019, 175, 118–127. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.Q.; Wang, Y.J.; Zhu, J.Q.; Pei, C.M.; Li, X.L. Effects of acid rain stress on photosynthetic physiological char-acteristics of seedlings of Sargassum pine in Jinyun Mountain. Resour. Environ. Yangtze Basin 2022, 31, 124–134. [Google Scholar] [CrossRef]
- Amini, F.; A Ehsanpour, A. Soluble Proteins, Proline, Carbohydrates and Na+/K+ Changes in Two Tomato (Lycopersicon esculentum Mill.) Cultivars under in vitro Salt Stress. Am. J. Biochem. Biotechnol. 2005, 1, 212–216. [Google Scholar] [CrossRef]
- Bao, G.; Tang, W.; An, Q.; Liu, Y.; Tian, J.; Zhao, N.; Zhu, S. Physiological effects of the combined stresses of freezing-thawing, acid precipitation and deicing salt on alfalfa seedlings. BMC Plant Biol. 2020, 20, 204. [Google Scholar] [CrossRef]
- Bao, G.; Tang, W.; He, F.; Chen, W.; Zhu, Y.; Fan, C.; Zhang, M.; Chang, Y.; Sun, J.; Ding, X. Physiological response in the leaf and stolon of white clover under acid precipitation and freeze–thaw stress. Funct. Plant Biol. 2020, 47, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yi, X.; Gao, X.; Wang, M.; Shao, C.; Lv, Z.; Chen, J.; Liu, Z.; Shen, C. Physiological and biochemical responses of tea seedlings (Camellia sinensis) to simulated acid rain conditions. Ecotoxicol. Environ. Saf. 2020, 192, 110315. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Hu, X.-H. Exogenous spermidine-induced changes at physiological and biochemical parameters levels in tomato seedling grown in saline-alkaline condition. Bot. Stud. 2014, 55, 58. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Debnath, B.; Sikdar, A.; Islam, S.; Hasan, K.; Li, M.; Qiu, D. Physiological and Molecular Responses to Acid Rain Stress in Plants and the Impact of Melatonin, Glutathione and Silicon in the Amendment of Plant Acid Rain Stress. Molecules 2021, 26, 862. [Google Scholar] [CrossRef]
- Zhang, X.; Du, Y.; Wang, L.; Zhou, Q.; Huang, X.; Sun, Z. Combined Effects of Lanthanum (III) and Acid Rain on Antioxidant Enzyme System in Soybean Roots. PLoS ONE 2015, 10, e0134546. [Google Scholar] [CrossRef]
- Sun, C.Q.; Yang, Y.J.; Guo, Z.L.; Qu, F. Effects of fertilizer application and density on soluble sugars, soluble protein and nitrate reductase in hybrid grain. J. Plant Nutr. 2015, 21, 1169–1177. [Google Scholar] [CrossRef]
- Debnath, B.; Hussain, M.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Qiu, D. Exogenous Melatonin Mitigates Acid Rain Stress to Tomato Plants through Modulation of Leaf Ultrastructure, Photosynthesis and Antioxidant Potential. Molecules 2018, 23, 388. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Average Plant Height (cm) | Average Ground Diameter (mm) | Source |
|---|---|---|---|
| Pinus massoniana | 21.83 ± 4.87 | 4.40 ± 0.91 | Chongqing |
| Phyllostachys edulis | 32.29 ± 14.91 | 2.13 ± 0.63 | Chongqing |
| Cinnamomum camphora | 33.99 ± 3.87 | 4.21 ± 1.21 | Chongqing |
| Month | May | June | July | August |
|---|---|---|---|---|
| Spraying volume (mm) | 175 | 206 | 233 | 179 |
| Tree Species | Treatment | LSP | LCP | Pmax | AQY | Rd |
|---|---|---|---|---|---|---|
| A | CK | 1654.67 ± 281.22 a | 20.09 ± 8.11 a | 26.43 ± 4.15 b | 0.063 ± 0.028 b | 1.83 ± 0.42 a |
| NR | 1055.07 ± 81.79 b | 13.87 ± 1.15 a | 33.40 ± 1.62 a | 0.119 ± 0.011 a | 2.39 ± 0.58 a | |
| pH 4.5 | 1182.22 ± 128.21 ab | 18.25 ± 6.59 a | 26.60 ± 4.48 b | 0.059 ± 0.030 b | 1.38 ± 0.66 a | |
| pH3.5 | 1271.32 ± 334.57 ab | 10.85 ± 1.12 a | 23.87 ± 0.01 b | 0.091 ± 0.013 ab | 1.63 ± 0.53 a | |
| pH2.5 | 1357.80 ± 558.47 ab | 11.54 ± 1.17 a | 22.22 ± 1.90 b | 0.069 ± 0.010 b | 1.30 ± 0.33 a | |
| B | CK | 895.00 ± 90.57 b | 15.47 ± 5.61 ab | 8.89 ± 0.12 bc | 0.037 ± 0.001 a | 1.51 ± 0.70 a |
| NR | 1315.03 ± 51.06 a | 8.13 ± 4.03 b | 10.93 ± 0.19 a | 0.025 ± 0.012 a | 0.21 ± 0.08 b | |
| pH4.5 | 1086.57 ± 53.49 ab | 4.47 ± 4.51 b | 9.33 ± 1.01 ab | 0.024 ± 0.011 a | 0.26 ± 0.04 b | |
| pH3.5 | 1146.23 ± 33.93 ab | 21.13 ± 9.10 a | 7.17 ± 0.01 c | 0.023 ± 0.007 a | 1.34 ± 0.74 ab | |
| pH2.5 | 1135.21 ± 244.48 ab | 7.04 ± 3.66 b | 7.52 ± 1.01 c | 0.022 ± 0.011 a | 0.31 ± 0.21 b | |
| C | CK | 884.32 ± 201.88 b | 10.73 ± 4.54 b | 14.20 ± 0.73 a | 0.059 ± 0.009 a | 1.13 ± 0.39 ab |
| NR | 960.64 ± 96.07 b | 21.57 ± 7.79 ab | 12.07 ± 0.68 abc | 0.045 ± 0.007 ab | 1.37 ± 0.34 ab | |
| pH4.5 | 1452.34 ± 90.28 a | 29.25 ± 11.49 ab | 13.37 ± 0.02 ab | 0.037 ± 0.016 ab | 1.63 ± 0.16 a | |
| pH3.5 | 847.07 ± 141.31 b | 11.62 ± 2.86 b | 10.18 ± 0.01 bc | 0.044 ± 0.005 ab | 0.94 ± 0.08 b | |
| pH2.5 | 759.05 ± 42.98 b | 35.04 ± 12.61 a | 8.84 ± 1.56 c | 0.029 ± 0.015 b | 1.71 ± 0.01 a |
| Name | Explains % | Contribution % | pseudo-F | p |
|---|---|---|---|---|
| Soluble protein | 47.3 | 56.1 | 38.6 | 0.002 |
| MDA | 11.0 | 13.1 | 11.1 | 0.002 |
| Relative chlorophyll | 9.0 | 10.7 | 11.3 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yang, F.; Wang, Y.; Zheng, Y.; Zhu, J. Effects of Acid Rain Stress on the Physiological and Biochemical Characteristics of Three Plant Species. Forests 2023, 14, 1067. https://doi.org/10.3390/f14051067
Zhang Y, Yang F, Wang Y, Zheng Y, Zhu J. Effects of Acid Rain Stress on the Physiological and Biochemical Characteristics of Three Plant Species. Forests. 2023; 14(5):1067. https://doi.org/10.3390/f14051067
Chicago/Turabian StyleZhang, Yuxuan, Feng Yang, Yunqi Wang, Yonglin Zheng, and Junlin Zhu. 2023. "Effects of Acid Rain Stress on the Physiological and Biochemical Characteristics of Three Plant Species" Forests 14, no. 5: 1067. https://doi.org/10.3390/f14051067