
Preliminary Study: Micropropagation Using Five Types of Chelated Iron and the Subsequent Acclimation of Blue Honeysuckle (Lonicera caerulea var. kamtschatica Sevast.)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chemicals
Nutrient Medium and Growth Conditions
2.2. Morphological Measurements and Observations
2.3. Statistical Analysis
3. Results
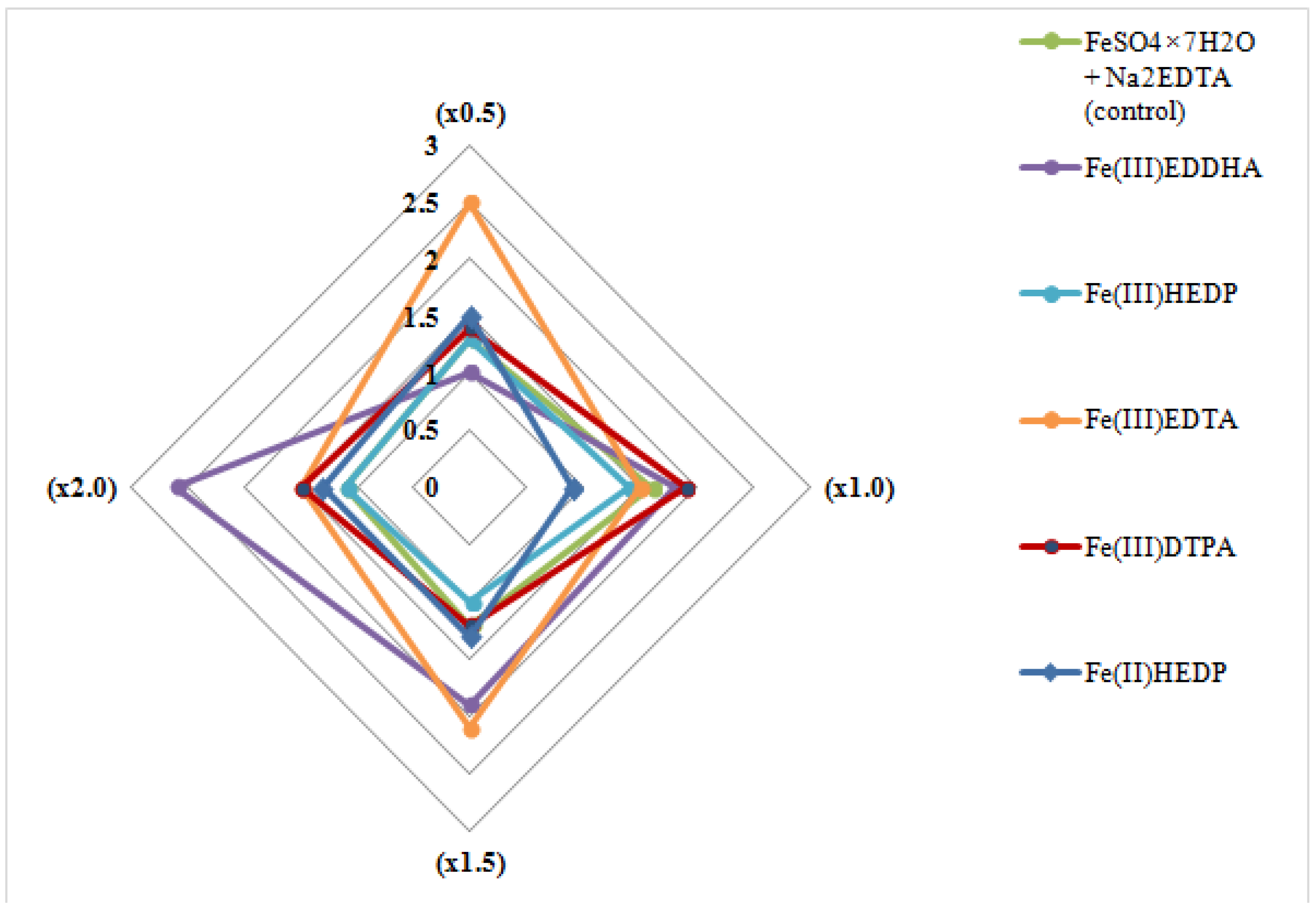
3.1. The Effect of Nutrient Medium Modification on the Multiplication Ability of the Blue Honeysuckle Explants (Stage S1)
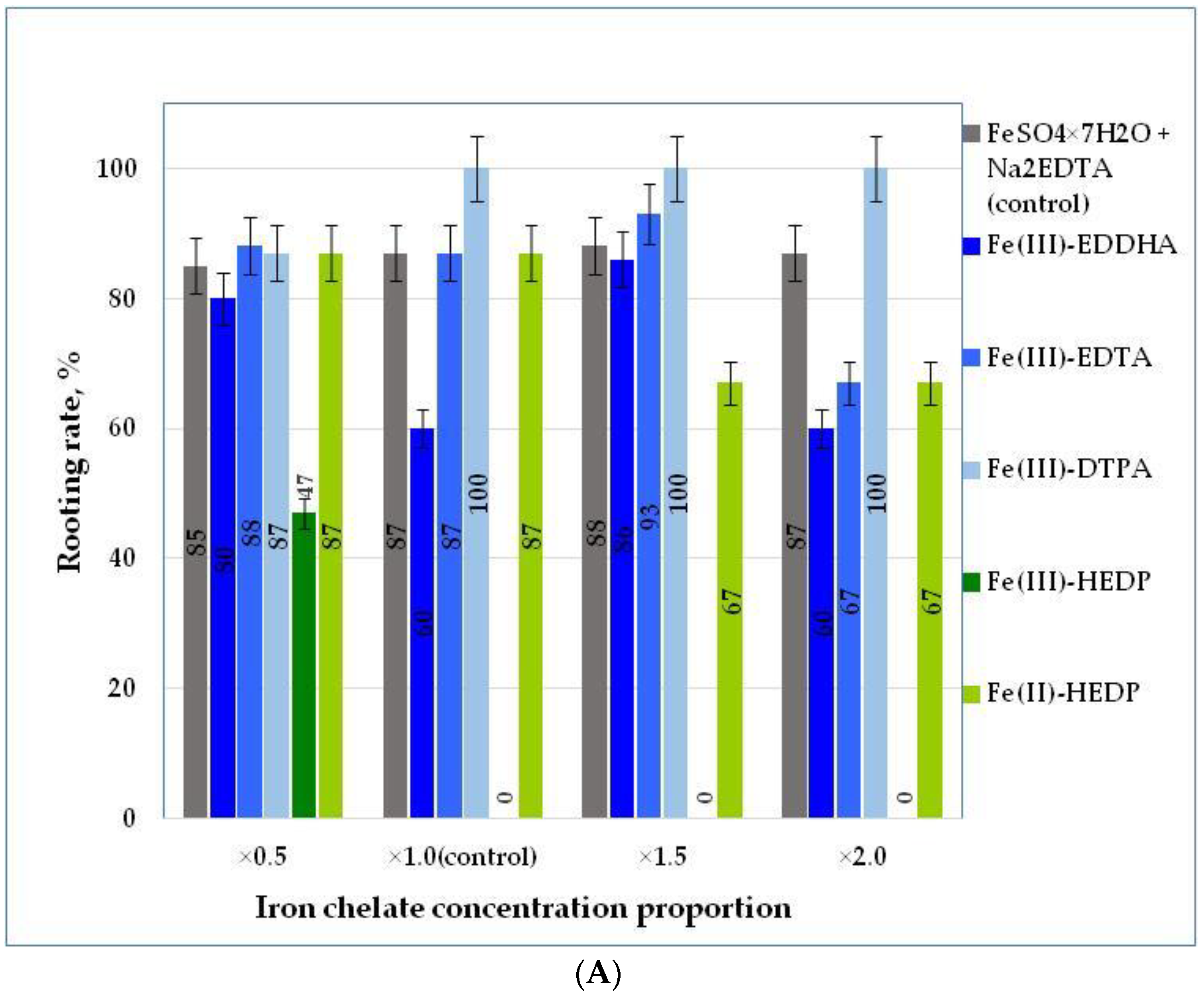
3.2. The Effect of Nutrient Medium Modification on the Rooting Rate of Blue Honeysuckle Explants during the Rooting Stage (Stage S2)
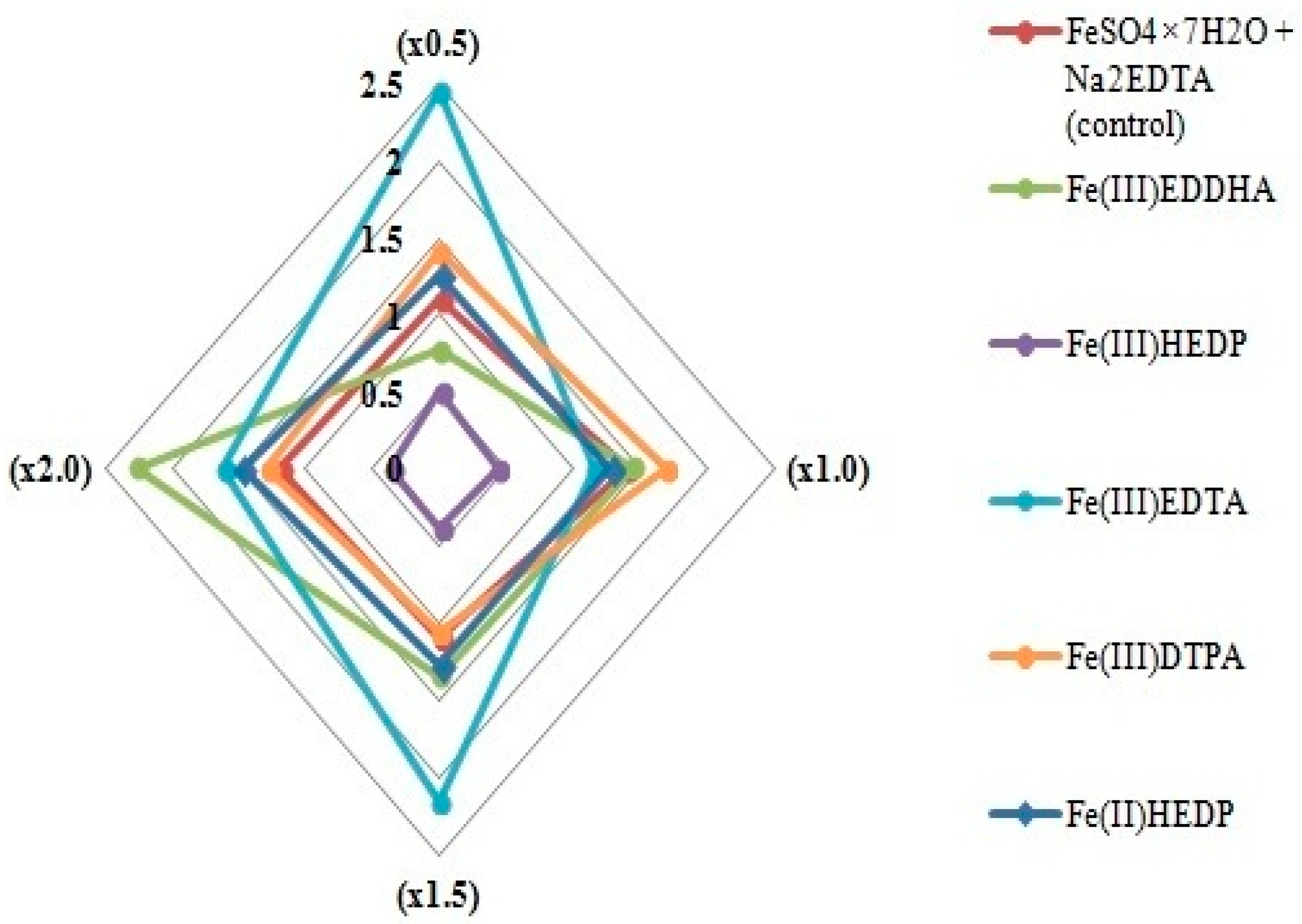
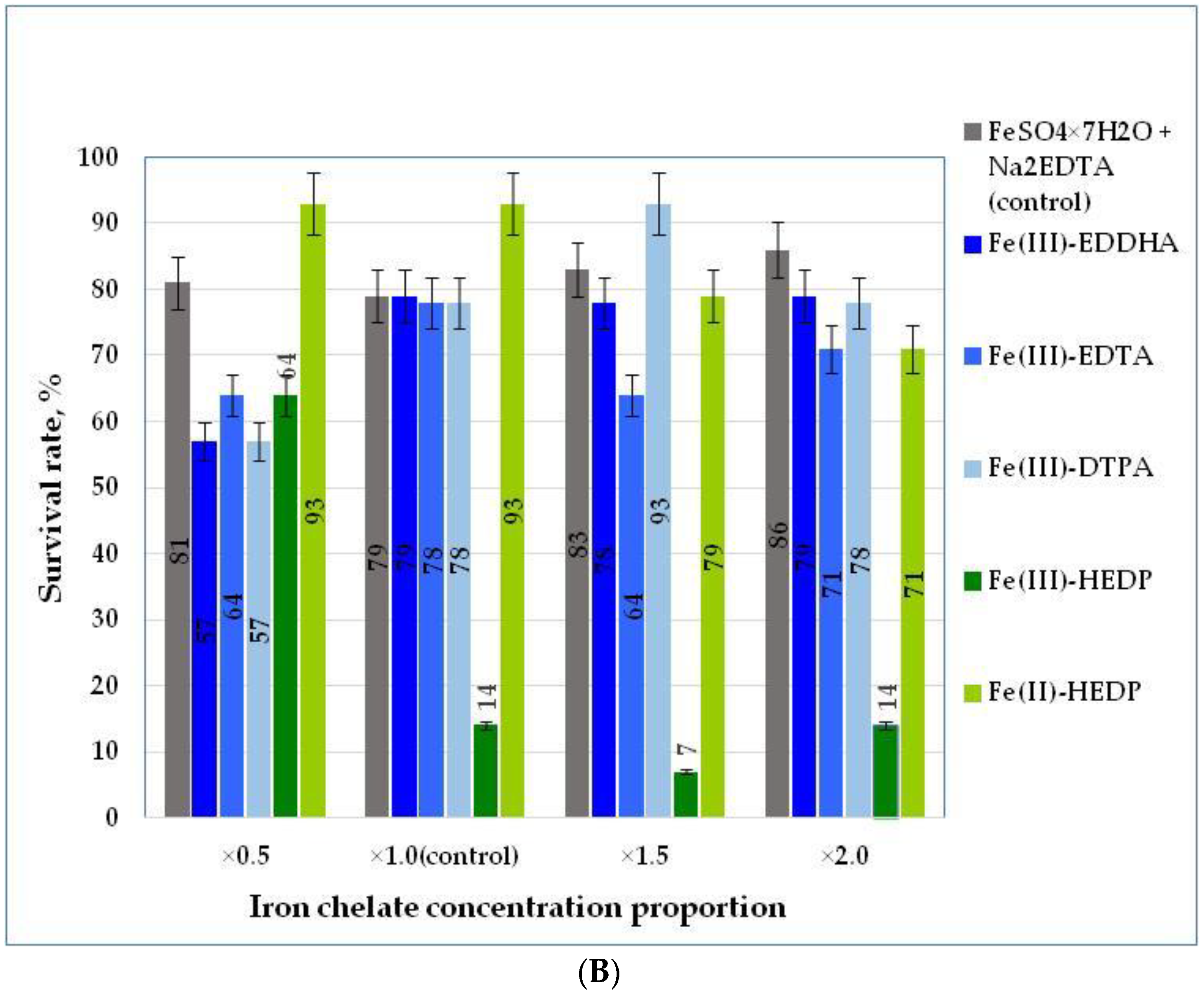
3.3. Acclimation to Nonsterile Conditions of Blue Honeysuckle Microplants Rooted on Modified Nutrient Media (Stage S3)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- McLennan, A.K. Book review: Food Health. Nutrition, technology, and public health. (Research methods for anthropological studies of food and nutrition (Ed. by by Jane Chrzan, John Brett). Anthropos 2018, 113, 710–711. [Google Scholar] [CrossRef]
- Waterlander, W.E.; Ni Mhurchu, C.; Eyles, H.; Vandevijvere, S.; Cleghorn, C.; Scarborough, P.; Swinburn, B.; Seidell, J. Food Future: Developing effective food systems interventions to improve public health nutrition. Agric. Syst. 2018, 160, 124–131. [Google Scholar] [CrossRef]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Superfruits: Phytochemicals, antioxidant efficacies, and health effect—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1580–1604. [Google Scholar] [CrossRef]
- Sharma, A.; Lee, H.-J. Lonicera caerulea: An updated account of its phytoconstituents and health-promoting activities. Trends Food Sci. Technol. 2021, 107, 130–149. [Google Scholar] [CrossRef]
- Caprioli, G.; Iannarelli, R.; Innocenti, M.; Bellumori, M.; Fiorini, D.; Sagratini, G.; Vittori, S.; Buccioni, M.; Santinelli, C.; Bramucci, M.; et al. Blue honeysuckle fruit (Lonicera caerulea L.) from eastern Russia: Phenolic composition, nutritional value and biological activities of its polar extracts. Food Funct. 2016, 7, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Celli, G.B.; Ghanem, A.; Brooks, M.S.L. Haskap berries (Lonicera caerulea L.)—A critical review of antioxidant capacity and health-related studies for potential value-added products. Food Bioprocess Technol. 2014, 7, 1541–1554. [Google Scholar] [CrossRef]
- Plekhanova, M.N. Blue honeysuckle (Lonicera caerulea L.)—A new commercial berry crop for temperate climate: Genetic resources and breeding. Acta Hortic. 2000, 538, 159–164. [Google Scholar] [CrossRef]
- Kolbanova, E.V. Simultaneous direct ex vitro rooting and adaptation of the blue honeysuckle micro-sprouts (Lonicera caerulea L. var. kamtschatica). Vesti Natsyyanal’nay Akademii Navuk Belarusi. Serya agrarnykh navuk = Proc. Natl. Acad. Sci. Belarus Agrarian Ser. 2020, 58, 298–310. [Google Scholar] [CrossRef]
- Zapolsky, Y.; Medvedeva, T.; Natalchuk, T.; Bublyk, M. Propagation of edible honeysuckle (Lonicera edulis Turcz) in in vitro conditions. Agric. Sci. Pract. 2018, 5, 18–26. [Google Scholar] [CrossRef]
- Sedlák, J.; Paprštein, F. In vitro propagation of blue honeysuckle. Hortic. Sci. 2007, 34, 129–131. [Google Scholar] [CrossRef]
- Nunková, J.; Szabóová, M.; Gajdošová, A. Protocols for adventitious regeneration of Amelanchier alnifolia var. cusickii and Lonicera kamtschatica ‘Jugana’. Plants 2021, 10, 1155. [Google Scholar] [CrossRef]
- Al, F.; Clapa, D.; Cristea, V.; Plopa, C. In vitro propagation of Lonicera kamtschatica. Agric.–Sci. Pract. 2014, 89, 90–99. [Google Scholar]
- Dziedzic, E. Propagation of blue honeysuckle (Lonicera caerulea var. kamtschatica Pojark.) in in vitro culture. J. Fruit Ornam. Plant Res. 2008, 16, 93–100. [Google Scholar]
- Krupa-Małkiewicz, M.; Ochmian, I. Propagation of blue honeysuckle (Lonicera caerulea L.) in in vitro culture. J. Basic Appl. Sci. 2014, 10, 164–169. [Google Scholar] [CrossRef]
- Krupa-Małkiewicz, M.; Ochmian, I.; Smolik, M.; Ostrowska, K.M. Composition of propagation method in in vitro and in vivo condition of Lonicera caerulea L. Folia Pomer. Univ. Technol. Stetin. Agric. Aliment. Pisc. Zootech. 2017, 334, 79–88. [Google Scholar] [CrossRef]
- Wojtania, A.; Markiewicz, M.; Góraj-Koniarska, J. Ex vitro rooting, acclimatization and genetic stability of Lonicera caerulea var. kamtschatica. J. Hortic. Res. 2020, 28, 61–70. [Google Scholar] [CrossRef]
- Mihaljević, I.; Tomaš, V.; Vuković, D.; Dugalić, K. Propagation of the blue honeysuckle (Lonicera caerulea L.) cultivars in in vitro culture. Pomol. Croat. 2019, 23, 41–48. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Li, Y.; Nie, Q.; Li, J. An efficient procedure for regeneration from leaf-derived calluses of Lonicera macranthoides ‘Jincuilei’, an important medicinal plant. HortScience 2009, 44, 746–750. [Google Scholar] [CrossRef]
- Hui, J.X.; Wen, C.S.; Hua, Z.Y.; Ming, L.X. Comparative study on different methods for Lonicera japonica Thunb. Micropropagation and acclimatization. J. Med. Plants Res. 2012, 6, 4389–4393. [Google Scholar] [CrossRef]
- Debnath, S.C. Strategies to propagate Vaccinium nuclear stocks for the Canadian berry industry. Can. J. Plant Sci. 2007, 87, 911–922. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Driver, J.A.; Kuniyuki, A.H. In vitro propagation of Paradox Walnut root stock. Am. Soc. Hortic. Sci. 1984, 19, 506–509. [Google Scholar] [CrossRef]
- Karhu, S.T. Axillary shoot proliferation of blue honeysuckle. Plant Cell Tissue Organ Cult. 1997, 48, 195–201. [Google Scholar] [CrossRef]
- Karhu, S.T. Rooting of blue honeysuckle microshoots. Plant Cell Tissue Organ Cult. 1997, 48, 153–159. [Google Scholar] [CrossRef]
- Morrissey, J.; Guerinot, M.L. Iron uptake and transport in plants: The good, the bad, and the ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef]
- Piagnani, C.; De Nisi, P.; Espen, L.; Zocchi, G. Adaptive responses to iron-dificiency in callus cultures of two cultivars of Vitis spp. J. Plant Physiol. 2003, 160, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Hiroshi, H.; Azizur, R.M.; Saitou, K.; Kobayashi, M.; Chikako, O. Influence of chelating ligands on bioavailability and mobility of iron in plant growth media and their effect on radish growth. Environ. Exp. Bot. 2011, 71, 345–351. [Google Scholar] [CrossRef]
- Van der Salm, T.P.M.; Van der Toorn, C.J.G.; Hänisch ten Cate, C.H.; Dubois, L.A.M.; De Vries, D.P.; Dons, H.J.M. Importance of the iron chelate formula for micropropagation of Rosa hybrida L. ‘Moneyway’. Plant Cell Tissue Organ Cult. 1994, 37, 73–77. [Google Scholar] [CrossRef]
- Trejgell, A.; Libront, I.; Tretyn, A. The effect of Fe-EDDHA on shoot multiplication and in vitro rooting of Carlina onopordifolia Besser. Acta Physiol. Plant 2012, 34, 2051–2055. [Google Scholar] [CrossRef]
- Molassiotis, A.N.; Dimassi, K.; Therios, I.; Diamantidis, G. Fe-EDDHA promotes rooting of rootstock GF-677 (Prunus amygdalus × P. persica) explants in vitro. Biol. Plant. 2003, 46, 141–144. [Google Scholar] [CrossRef]
- Antonopoulou, C.; Dimassi, K.; Therios, I.; Chatzissavvidis, C.; Papadakis, I. The effect of Fe-EDDHA and ascorbic acid on in vitro rooting of the peach rootstock GF-677 explants. Acta Physiol. Plant. 2007, 29, 559–561. [Google Scholar] [CrossRef]
- Garrison, W.; Dale, A.; Saxena, P.K. Improved shoot multiplication and development in hybrid hazelnut nodal cultures by ethylenediamine di-2-hydroxy-phenylacetic acid (Fe-EDDHA). Can. J. Plant Sci. 2013, 93, 511–521. [Google Scholar] [CrossRef]
- Castillo, B.; Smith, M.A.L.; Madhavi, D.L. Effect of irradiance level and iron chelate source on the shoot tip culture of Carica papaya L. HortScience 1997, 32, 1120–1123. [Google Scholar] [CrossRef]
- Zawadzka, M.; Orlikowska, T. The influence of FeEDDHA in red raspberry cultures during shoot multiplication and adventitious regeneration from leaf explants. Plant Cell Tissue Organ Cult. 2006, 85, 145–149. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Shukla, M.R.; Sullivan, J.A.; Saxena, P.K. Iron supplementation promotes in vitro shoot induction and multiplication of Baptisia australis. Plant Cell Tissue Organ Cult. 2017, 129, 145–152. [Google Scholar] [CrossRef]
- Hunková, J.; Libiaková, G.; Fejér, J.; Vujović, T.; Gajdošová, A. Testing of different iron sources and concentrations on shoot multiplication of blackberry (Rubus fruticosus L.). Genetika 2018, 50, 351–356. [Google Scholar] [CrossRef]
- Nikulina, E.; Akimova, S.; Tsirulnikova, N.; Kirkach, V. Different Fe (II) and Fe (III) complexes in clonal micropropagation of Gooseberry. IOP Conf. Ser. Earth Environ. Sci. 2020, 578, 012015. [Google Scholar] [CrossRef]
- Nikulina, E.A.; Akimova, S.V.; Tsirulnikova, N.V.; Glinushkin, A.P.; Dolgoborodov, I.O.; Kirkach, V.V. Screening of different Fe (II) and Fe (III) complexes at the stage of rhizogenesis in vitro of gooseberry plants. J. Phys. Conf. Ser. 2021, 1942, 012075. [Google Scholar] [CrossRef]
- Kalinitchenko, V.P.; Glinushkin, A.P.; Kudeyarov, V.N.; Minkina, T.M.; Chernenko, V.V.; Sushkova, S.N.; Mandzhieva, S.S.; Makarenkov, D.A.; Ilyina, L.P. Biogeosystem Technique for sustsainable agriculture, water scarcity overcoming healthy soil and environment. In Proceedings of the ACS Fall Meeting, San Francisco, CA, USA, 17–20 August 2020; p. 3429424. [Google Scholar] [CrossRef]
- Isachkin, A.V. Fundamentals of Scientific Research in Horticulture: A Textbook for Universities; Isachkin, A.V., Kryuchkova, V.A., Eds.; Lan: St. Petersburg, Russia, 2020; p. 420. [Google Scholar]
- Rossall, S.; Qing, C.; Paneri, M.; Bennett, M.; Swarup, R. A ‘growing’ role for phosphites in promoting plant growth and development. Acta Hortic. 2016, 1148, 61–68. [Google Scholar] [CrossRef]
- Thao, H.T.B.; Yamakawa, T. Phosphite (phosphorous acid): Fungicide, fertilizer or biostimulator? Soil Sci. Plant Nutr. 2009, 55, 228–234. [Google Scholar] [CrossRef]
- Fritz, C.D.; Evans, W.F.; Cooke, A.R. Growth Regulation Process. U.S. Patent 3,879,188, 22 April 1975. [Google Scholar]
- Jolley, V.D.; Cook, K.A.; Nansen, N.C.; Stevens, W.B. Plant physiological responses for genotypic evaluation of iron efficiency in strategy I and strategy II plants—A review. J. Plant Nutr. 1996, 19, 1241–1255. [Google Scholar] [CrossRef]
- Hindt, M.N.; Guerinot, M.L. Getting a sense for signals: Regulation of the plant iron deficiency response. Biochim. Biophys. Acta (BBA)–Mol. Cell Res. 2012, 1823, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, V.; Grishina, E.; Kalinitchenko, V.; Glinushkin, A.; Kudeyarov, V.; Gudkov, S.; Savostyanov, A.; Minkina, T.; Ilyin, V.; Mandzhieva, S. Biogeosystem Tecnique methodology as a new chemical soil-biological engineering foundation for the safe expanded technological development in the Noosphere. In Proceedings of the EGU General Assembly 2021, online, 19–30 April 2021. [Google Scholar] [CrossRef]
- Diatlova, N.M.; Lavrova, O.Y.; Temkina, V.Y.; Kireeva, A.Y.; Seliverstova, I.A.; Rudakova, G.Y.; Cirulnikova, N.V.; Dobrikova, E.O. The Use of Chelating Agents in Agriculture; Overview of ser. ‘Reagents and highly purified substances’ Separate Edition; NIITEKHIM: Moscow, Russia, 1984; 31p. [Google Scholar]
- Kalinitchenko, V.P.; Glinushkin, A.P.; Sharshak, V.K.; Ladan, E.P.; Minkina, T.M.; Sushkova, S.N.; Mandzhieva, S.S.; Batukaev, A.A.; Chernenko, V.V.; Ilyina, L.P.; et al. Intra-Soil Milling for Stable Evolution and High Productivity of Kastanozem Soil. Processes 2021, 9, 1302. [Google Scholar] [CrossRef]
- Severina, V.; Proklin, V.; Rykhlik, A.; Kalinitchenko, V.; Glinushkin, A.; Dubenok, N.; Minkina, T.; Nesvat, A.; Deryabkina, I.; Zamulina, I. Biogeosystem technique water paradigm for prevention of the world water scarcity and cardinal transformation of current irrigation practice. In Proceedings of the EGU General Assembly 2021, online, 19–30 April 2021; p. EGU21-14922. [Google Scholar] [CrossRef]
- Kalinichenko, V.P.; Glinushkin, A.P.; Sokolov, M.S.; Zinchenko, V.E.; Minkina, T.M.; Mandzhieva, S.S.; Sushkova, S.N.; Makarenkov, D.A.; Bakoyev, S.Y.; Il’ina, L.P. Impact of soil organic matter on calcium carbonate equilibrium and forms of Pb in water extracts from Kastanozem complex. J. Soils Sediments 2019, 19, 2717–2728. [Google Scholar] [CrossRef]
- Konchekov, E.M.; Glinushkin, A.P.; Kalinitchenko, V.P.; Artem’ev, K.; Burmistrov, D.E.; Kozlov, V.A.; Kolik, L.V. Properties and Use of Water Activated by Plasma of Piezoelectric Direct Discharge. Front. Phys. 2021, 8, 616385. [Google Scholar] [CrossRef]
- Kalinitchenko, V.P.; Glinushkin, A.P.; Swidsinski, A.V.; Minkina, T.M.; Andreev, A.G.; Mandzhieva, S.S.; Sushkova, S.N.; Makarenkov, D.A.; Ilyina, L.P.; Chernenko, V.V.; et al. Thermodynamic mathematical model of the Kastanozem complex and new principles of sustainable semiarid protective silviculture management. Environ. Res. 2021, 194, 110605. [Google Scholar] [CrossRef]
- Tsirulnikova, N.V.; Nukulina, E.A.; Akimova, S.V.; Kirkach, V.V.; Glinushkin, A.P.; Podkovyrov, I.Y. In vitro effect of Replacement Mineral Salts of Trace Elements with P-containing Chelates to Improve Rooting of Cherry Rootstock (cv. VC-13). IOP Conf. Ser. Earth Environ. Sci. 2021, 663, 012042. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Molar Mass, g | Iron Content in Powder/in Solution, % | Concentration of Fe Solution, mgL−1 | |||
|---|---|---|---|---|---|---|
| Reduced (×0.5) Fe-2.79 | Standard (×1.0) Fe-5.58 | Increased (×1.5) Fe-8.37 | Doubled (×2.0) Fe-11.16 | |||
| Fe(III)-EDDHA (mL L−1) | 439 | In solution 0.015 | 18.6 | 37.2 | 55.8 | 74.4 |
| Fe(III)-EDTA (mg L−1) | 368 | In powder 13.4 | 0.02 | 0.04 | 0.06 | 0.08 |
| Fe(III)-DTPA (mg L−1) | 470 | In powder 9.81 | 0.03 | 0.06 | 0.09 | 0.12 |
| Fe(III)-HEDP (mL L−1) | 259 | In solution 0.65 | 0.43 | 0.86 | 1.29 | 1.72 |
| Fe(II)-HEDP (mL L−1) | 260 | In solution 0.52 | 0.54 | 1.07 | 1.61 | 2.15 |
| Parameter | Multiplication | Rooting | Acclimation |
|---|---|---|---|
| Multiplication factor | + | ||
| Rooting rate, % | + | ||
| Survival rate, % | + | ||
| Average number of shoots, pcs. | + | ||
| Average shoots length, cm | + | + | + |
| Leaf surface area, cm2 | + | ||
| Average number of roots, pcs. | + | + | |
| Average roots length, cm | + | + | |
| The total length of the roots, cm | + | + |
| Concentrations of Iron Chelates in Modified Nutrient Media (Factor A) | Iron Chelates (Factor B) | Factor A Mean | |||||
|---|---|---|---|---|---|---|---|
| FeSO4 × 7H2O + Na2EDTA + SD * (Control) | Fe(III)-EDDHA + SD | Fe(III)-EDTA + SD | Fe(III)-DTPA + SD | Fe(III)-HEDP + SD | Fe(II)-HEDP + SD | ||
| Rooting rate, % | LSD05 a = 7.49 | ||||||
| ×0.5 | 85.0 ± 3.03 | 80.0 ± 2.47 a** | 88.0 ± 4.56 | 87.0 ± 2.77 | 47.0 ± 5.55 | 87.0 ± 2.25 | 79.0 |
| ×1.0 | 87.0 ± 2.52 | 60.0 ± 6.63 | 87.0 ± 2.33 | 100 ± 0.00 b | 0 | 87.0 ± 3.76 | 70.2 |
| ×1.5 | 88.0 ± 3.10 | 86.0 ± 0.47 a | 93.0 ± 2.33 | 100 ± 0.00 b | 0 | 67.0 ± 5.57 | 72.3 |
| ×2.0 | 87.0 ± 3.62 | 60.0 ± 1.36 | 67.0 ± 6.25 | 100 ± 0.00 b | 0 | 67.0 ± 4.65 | 63.5 |
| Factor B mean LSD05 b = 8.41 | 86.8 | 71.5 | 83.8 | 96.8 | 11.8 | 77.0 | |
| LSD05 ab = 10.40 to compare private averages | |||||||
| Average number of shoots, pcs. | LSD05 a = 0.40 | ||||||
| ×0.5 | 1.4 ± 0.83 | 1.3 ± 0.44 | 1.4 ± 0.49 | 3.1 ± 0.44 a,b | 1.8 ± 0.83 a,b | 0.9 ± 1.36 | 1.7 |
| ×1.0 | 1.5 ± 0.72 | 1.7 ± 0.50 | 1.5 ± 0.50 | 2.5 ± 0.60 b | 0.9 ± 0.68 | 1.1 ± 1.36 | 1.5 |
| ×1.5 | 1.5 ± 0.79 | 3.3 ± 0.79 a,b,ab | 1.3 ± 0.47 | 2.5 ± 1.12 a,b | 1.1 ± 0.85 | 1.7 ± 1.67 a | 1.9 |
| ×2.0 | 1.5 ± 0.81 | 1.3 ± 0.71 | 1.7 ± 0.70 | 2.2 ± 0.44 b | 1.2 ± 0.83 | 1.6 ± 1.47 a | 1.6 |
| Factor B mean LSD05 b = 0.34 | 1.5 | 1.9 | 1.5 | 2.6 | 1.3 | 1.3 | |
| LSD05 ab = 1.14 to compare private averages | |||||||
| Average shoot length, cm | LSD05 a = 0.39 | ||||||
| ×0.5 | 1.6 ± 0.85 | 2.4 ± 0.90 a,b | 2.2 ± 1.38 a,b | 1.0 ± 1.20 | 0.9 ± 0.39 | 0.9 ± 0.43 | 1.5 |
| ×1.0 | 1.8 ± 0.72 | 2.1 ± 1.49 b | 2.1 ± 1.09 b | 0.9 ± 1.01 | 0.4 ± 0.34 | 1.7 ± 0.36 | 1.5 |
| ×1.5 | 1.5 ± 0.84 | 0.8 ± 0.98 | 1.8 ± 0.94 b | 0.9 ± 0.50 | 0.4 ± 0.40 | 1.4 ± 0.44 | 1.1 |
| ×2.0 | 1.4 ± 0.95 | 1.5 ± 1.08 | 1.3 ± 0.85 | 0.8 ± 0.63 | 0.6 ± 0.47 | 1.5 ± 0.30 | 1.2 |
| Factor B mean LSD05 b = 0.33 | 1.6 | 1.7 | 1.9 | 0.9 | 0.6 | 1.4 | |
| LSD05 ab = 1.13 to compare private averages | |||||||
| Average number of roots, pcs. | LSD05 a = 0.97 | ||||||
| ×0.5 | 2.7 ± 1.59 | 3.7 ± 2.60 b | 4.2 ± 2.48 b | 3.1 ± 2.24 | 1.1 ± 1.54 a | 3.6 ± 2.38 b | 3.1 |
| ×1.0 | 2.5 ± 1.45 | 3.9 ± 3.07 b | 4.4 ± 2.55 b | 3.3 ± 1.77 b | 0 | 3.1 ± 1.89 | 2.9 |
| ×1.5 | 2.6 ± 1.77 | 3.3 ± 3.19 | 5.5 ± 2.60 a b | 2.3 ± 1.12 | 0 | 4.9 ± 2.30 a,b | 3.1 |
| ×2.0 | 2.9 ± 1.91 | 4.4 ± 2.06 b | 4.2 ± 3.56 b | 3.0 ± 1.74 | 0 | 2.1 ± 2.76 | 2.8 |
| Factor B mean LSD05 b = 0.81 | 2.7 | 3.8 | 4.6 | 2.9 | 0.3 | 3.4 | |
| LSD05 ab = 2.77 to compare private averages | |||||||
| Average root length, cm | LSD05 a = 0.49 | ||||||
| ×0.5 | 1.1 ± 0.69 | 1.3 ± 0.62 | 1.7 ± 0.86 b | 0.7 ± 0.77 | 0.4 ± 0.53 | 1.0 ± 0.39 | 1.0 |
| ×1.0 | 1.2 ± 0.70 | 1.7 ± 0.83 b | 1.4 ± 0.71 | 0.7 ± 0.96 | 0 | 0.9 ± 0.38 | 1.0 |
| ×1.5 | 0.9 ± 0.68 | 1.3 ± 4.50 | 1.6 ± 0.67 b | 0.5 ± 0.97 | 0 | 2.2 ± 0.43 a,b | 1.1 |
| ×2.0 | 0.9 ± 0.69 | 1.6 ± 0.48 b | 1.2 ± 0.95 | 0.4 ± 0.50 | 0 | 0.5 ± 0.44 | 0.8 |
| Factor B mean LSD05 b = 0.41 | 1.0 | 1.5 | 1.5 | 0.6 | 0.1 | 1.2 | |
| LSD05 ab = 1.39 to compare private averages | |||||||
| Average total root length, cm | LSD05 a = 1.57 | ||||||
| ×0.5 | 3.3 ± 2.69 | 6.2 ± 3.51 b | 7.9 ± 4.56 b | 2.5 ± 5.19 | 1.1 ± 1.55 | 4.5 ± 2.11 | 4.3 |
| ×1.0 | 3.5 ± 2.37 | 6.7 ± 4.43 | 6.9 ± 3.83 | 2.6 ± 4.24 | 0 | 4.2 ± 1.80 | 5.9 |
| ×1.5 | 3.4 ± 2.70 | 3.9 ± 5.26 | 8.9 ± 5.44 a,b | 1.9 ± 2.04 | 0 | 6.1 ± 2.42 a,b | 4.0 |
| ×2.0 | 3.3 ± 3.01 | 7.0 ± 1.86 b | 7.9 ± 7.99 b | 1.7 ± 2.93 | 0 | 1.7 ± 1.66 | 3.6 |
| Factor B mean LSD05 b = 1.33 | 3.4 | 6.9 | 7.9 | 2.2 | 0.3 | 4.1 | |
| LSD05 ab = 4.51 to compare private averages | |||||||
| Concentrations of Iron Chelates in Modified Nutrient Media (Factor A) | Iron Chelates (Factor B) | Factor A Mean | |||||
|---|---|---|---|---|---|---|---|
| FeSO4 × 7H2O + Na2EDTA + SD * (Control) | Fe(III)-EDDHA+ SD | Fe(III)-EDTA + SD | Fe(III)-DTPA + SD | Fe(III)-HEDP + SD | Fe(II)-HEDP + SD | ||
| Survival rate, % | LSD05 a = 6.32 | ||||||
| ×0.5 | 81.0 ± 5.89 | 57.0 ± 4.42 | 64.0 ± 3.45 | 57.0 ± 4.25 | 64.0 ± 3.15 a | 93.0 ± 4.45 b | 69.3 |
| ×1.0 | 79.0 ± 5.56 | 79.0 ± 6.66 | 78.0 ± 2.56 | 78.0 ± 4.23 | 14.0 ± 2.42 | 93.0 ± 4.12 b | 70.2 |
| ×1.5 | 83.0 ± 5.91 | 78.0 ± 7.77 | 64.0 ± 6.66 | 93.0 ± 4.12 a,b | 7.0 ± 3.34 | 79.0 ± 5.54 | 67.8 |
| ×2.0 | 86.0 ± 6.33 a** | 79.0 ± 5.52 | 71.0 ± 5.26 | 78.0 ± 2.16 | 14.0 ± 2.25 | 71.0 ± 5.12 | 66.5 |
| Factor B mean LSD05 b = 6.27 | 82.3 | 73.3 | 69.3 | 76.5 | 24.8 | 84.0 | |
| LSD05 ab = 12.23 to compare private averages | |||||||
| Average shoot length, cm | LSD05 a = 2.84 | ||||||
| ×0.5 | 7.9 ± 5.12 | 12.3 ± 4.82 a | 9.2 ± 5.62 | 9.9 ± 5.46 | 8.3 ± 5.50 b | 10.6 ± 5.42 a | 9.7 |
| ×1.0 | 6.7 ± 4.68 | 10.4 ± 4.84 a | 12.1 ± 4.61 a | 12.4 ± 4.25 a | 2.7 ± 0.25 | 10.9 ± 4.86 a | 9.2 |
| ×1.5 | 8.7 ± 5.11 | 15.0 ± 1.28 a,b | 13.6 ± 3.18 a | 13.4 ± 3.77 a | 2.6 ± 0.00 | 9.3 ± 6.06 | 10.4 |
| ×2.0 | 9.9 ± 5.41 b | 13.2 ± 4.08 a,b | 13.0 ± 4.99 a | 12.2 ± 4.71 | 2.5 ± 0.25 a | 8.7 ± 6.26 | 9.9 |
| Factor B mean LSD05 b = 2.39 | 8.3 | 12.7 | 12.0 | 15.3, b | 4.0 | 9.9 | |
| LSD05 ab = 8.13 to compare private averages | |||||||
| Sheet surface area, cm2 | LSD05 a = 5.52 | ||||||
| ×0.5 | 17.9 ± 10.66 | 25.3 ± 9.51 b | 20.4 ± 9.96 | 21.8 ± 11.49 | 18.4 ± 11.99 a | 22.6 ± 11.46 b | 21.1 |
| ×1.0 | 16.5 ± 11.01 | 23.8 ± 9.23 b | 26.6 ± 11.78 b | 26.6 ± 7.76 b | 2.2 ± 0.65 | 25.0 ± 10.35 b | 20.1 |
| ×1.5 | 19.5 ± 8.64 | 31.6 ± 3.28 a,b | 29.8 ± 5.60 b | 28.7 ± 6.61 b | 4.5 ± 0.00 | 19.8 ± 13.24 | 22.3 |
| ×2.0 | 20.8 ± 8.28 | 28.8 ± 7.07 b | 27.7 ± 10.03 b | 26.9 ± 8.32 b | 1.9 ± 0.25 | 18.4 ± 12.69 | 20.8 |
| Factor B mean LSD05 b = 4.65 | 18.7 | 27.4 | 26.1 | 26.0 | 6.8 | 21.5 | |
| LSD05 ab = 15.83 to compare private averages | |||||||
| Average number of roots, pcs. | LSD05 a = 0.53 | ||||||
| ×0.5 | 3.5 ± 0.89 | 4.1 ± 1.05 a,b | 3.7 ± 0.94 | 3.8 ± 1.09 | 3.4 ± 1.07 | 4.0 ± 1.04 b | 3.8 |
| ×1.0 | 3.4 ± 0.88 | 4.1 ± 0.90 a | 4.1 ± 0.79 a | 4.5 ± 0.78 a | 2.0 ± 0.00 | 4.0 ± 0.78 a | 3.7 |
| ×1.5 | 3.6 ± 0.93 | 4.5 ± 0.50 a,b | 4.4 ± 0.68 a,b | 4.3 ± 0.72 a,b | 3.0 ± 0.00 | 3.6 ± 1.23 | 3.9 |
| ×2.0 | 3.7 ± 0.94 | 4.5 ± 0.66 a,b | 4.2 ± 0.98 a,b | 4.3 ± 0.75 a,b | 2.5 ± 0.50 | 3.4 ± 1.28 | 3.8 |
| Factor B mean LSD05 b = 0.44 | 3.6 | 4.3 | 4.1 | 4.2 | 2.7 | 3.8 | |
| LSD05 ab = 1.51 to compare private averages | |||||||
| Average root length, cm | LSD05 a = 1.08 | ||||||
| ×0.5 | 4.7 ± 1.97 | 6.1 ± 1.76 a | 5.1 ± 2.11 | 4.3 ± 1.72 | 4.5 ± 2.17 b | 5.5 ± 1.94 | 5.0 |
| ×1.0 | 4.4 ± 2.10 | 5.2 ± 1.32 | 6.0 ± 1.49 a | 5.8 ± 1.69 a | 2.0 ± 0.15 | 5.4 ± 1.96 | 4.8 |
| ×1.5 | 5.2 ± 2.10 | 7.2 ± 0.43 a,b | 6.3 ± 1.33 a | 5.9 ± 1.49 | 2.1 ± 0.00 | 4.9 ± 2.58 | 5.3 |
| ×2.0 | 5.5 ± 1.93 b | 6.5 ± 0.93 b | 6.1 ± 1.62 a | 5.9 ± 1.35 | 2.1 ± 0.10 | 4.5 ± 2.44 | 5.1 |
| Factor B mean LSD05 b = 0.91 | 5.0 | 6.3 | 5.9 | 5.5 | 2.7 | 5.1 | |
| LSD05 ab = 3.09 to compare private averages | |||||||
| Average total root length, cm | LSD05 a = 5.61 | ||||||
| ×0.5 | 17.0 ± 9.41 | 26.1 ± 11.04 b | 23.9 ± 11.67 b | 22.7 ± 10.21 b | 19.6 ± 11.83 a | 23.3 ± 11.75 b | 22.1 |
| ×1.0 | 17.2 ± 9.64 | 24.0 ± 9.32 b | 26.7 ± 9.34 b | 28.5 ± 9.12 b | 6.5 ± 0.50 | 25.9 ± 11.40 b | 21.5 |
| ×1.5 | 23.6 ± 9.61 a | 34.3 ± 3.49 a | 32.2 ± 6.37 | 29.9 ± 6.13 | 7.5 ± 0.00 | 20.1 ± 12.71 | 24.6 |
| ×2.0 | 23.5 ± 9.38 a | 30.0 ± 5.85 a,b | 27.9 ± 8.83 | 28.5 ± 9.26 b | 5.5 ± 0.50 | 20.2 ± 13.67 | 22.6 |
| Factor B mean LSD05 b = 4.73 | 20.3 | 28.6 | 27.7 | 27.4 | 9.8 | 22.4 | |
| LSD05 ab = 16.08 to compare private averages | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glinushkin, A.; Akimova, S.; Nikulina, E.; Tsirulnikova, N.; Kirkach, V.; Kalinitchenko, V.; Radzhabov, A.; Radkevich, E.; Marchenko, L.; Solovyov, A.; et al. Preliminary Study: Micropropagation Using Five Types of Chelated Iron and the Subsequent Acclimation of Blue Honeysuckle (Lonicera caerulea var. kamtschatica Sevast.). Forests 2023, 14, 821. https://doi.org/10.3390/f14040821
Glinushkin A, Akimova S, Nikulina E, Tsirulnikova N, Kirkach V, Kalinitchenko V, Radzhabov A, Radkevich E, Marchenko L, Solovyov A, et al. Preliminary Study: Micropropagation Using Five Types of Chelated Iron and the Subsequent Acclimation of Blue Honeysuckle (Lonicera caerulea var. kamtschatica Sevast.). Forests. 2023; 14(4):821. https://doi.org/10.3390/f14040821
Chicago/Turabian StyleGlinushkin, Alexey, Svetlana Akimova, Elena Nikulina, Nina Tsirulnikova, Vadim Kirkach, Valery Kalinitchenko, Agamagomed Radzhabov, Elena Radkevich, Liudmila Marchenko, Alexandr Solovyov, and et al. 2023. "Preliminary Study: Micropropagation Using Five Types of Chelated Iron and the Subsequent Acclimation of Blue Honeysuckle (Lonicera caerulea var. kamtschatica Sevast.)" Forests 14, no. 4: 821. https://doi.org/10.3390/f14040821