
Altitudinal Gradients Modify the Ecological Stoichiometry and Enzyme Activities of Soil in a Natural Secondary Quercus spp. Forest of the Dabie Mountains
Abstract
:1. Introduction
2. Materials and Methods
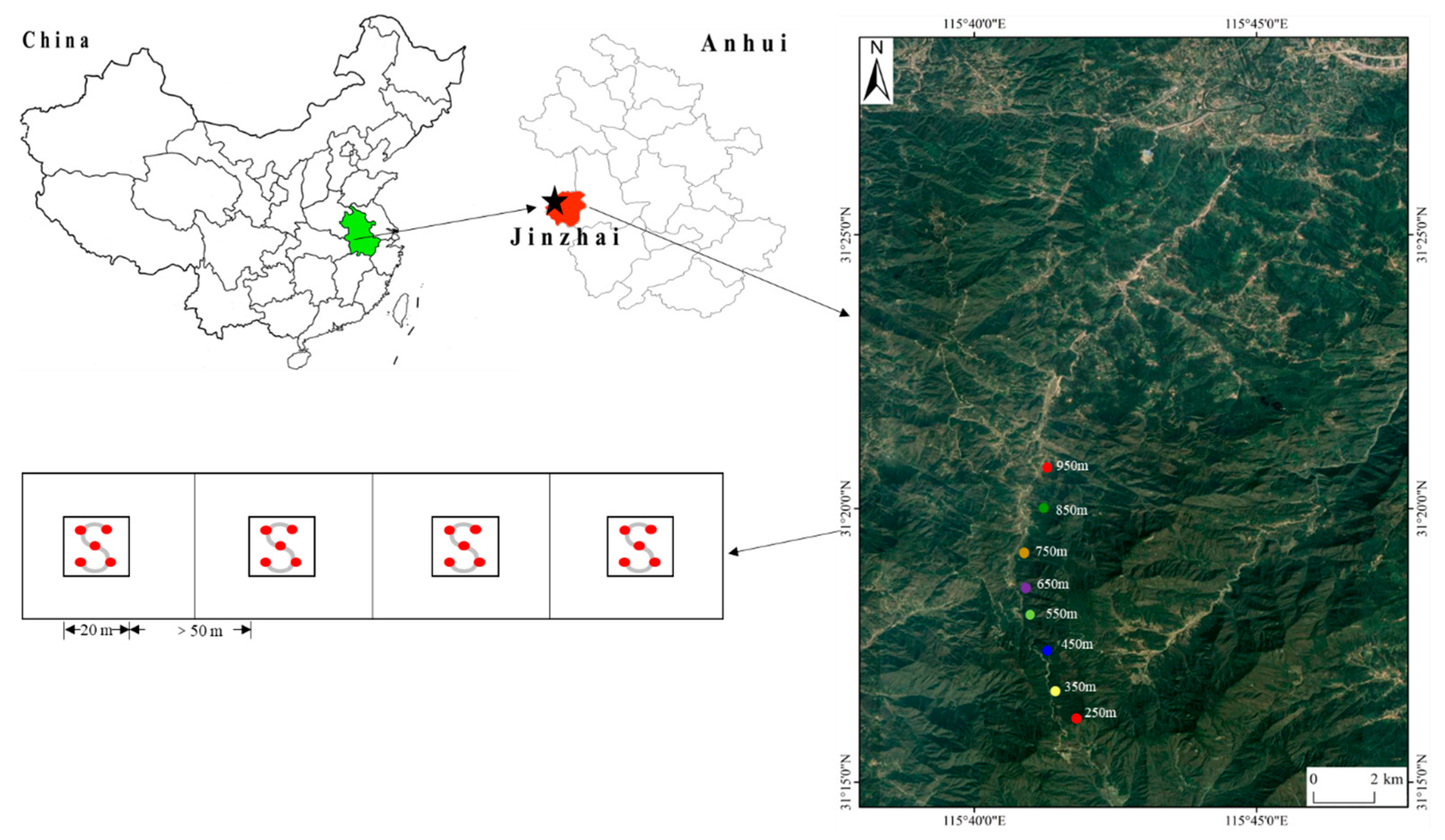
2.1. Study Area
2.2. Sampling Design
2.3. Laboratory Analysis
2.4. Data Analysis
3. Results and Discussion
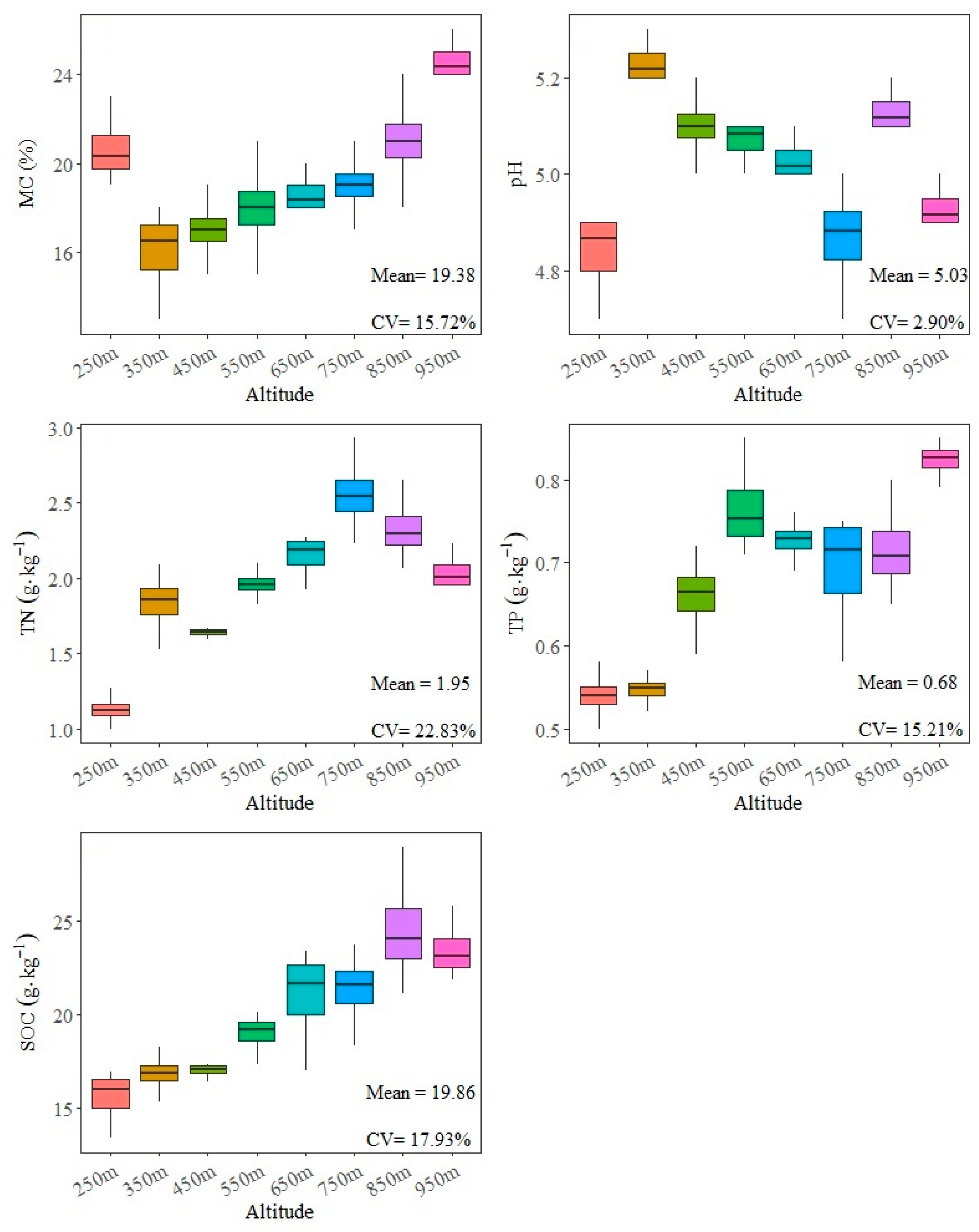
3.1. Impacts of Altitude on Soil Physicochemical Properties
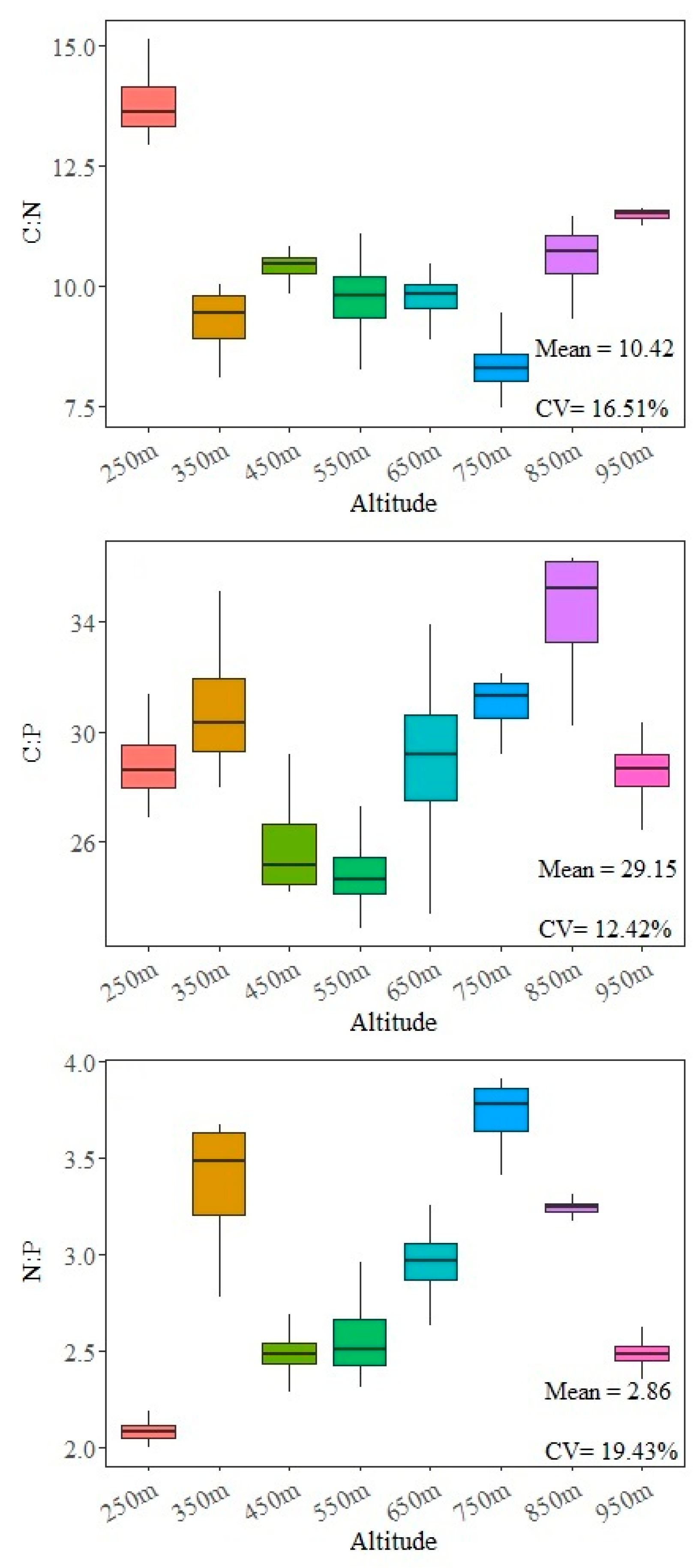
3.2. Elevation Characteristics of Soil C, N, and P Ecological Stoichiometry
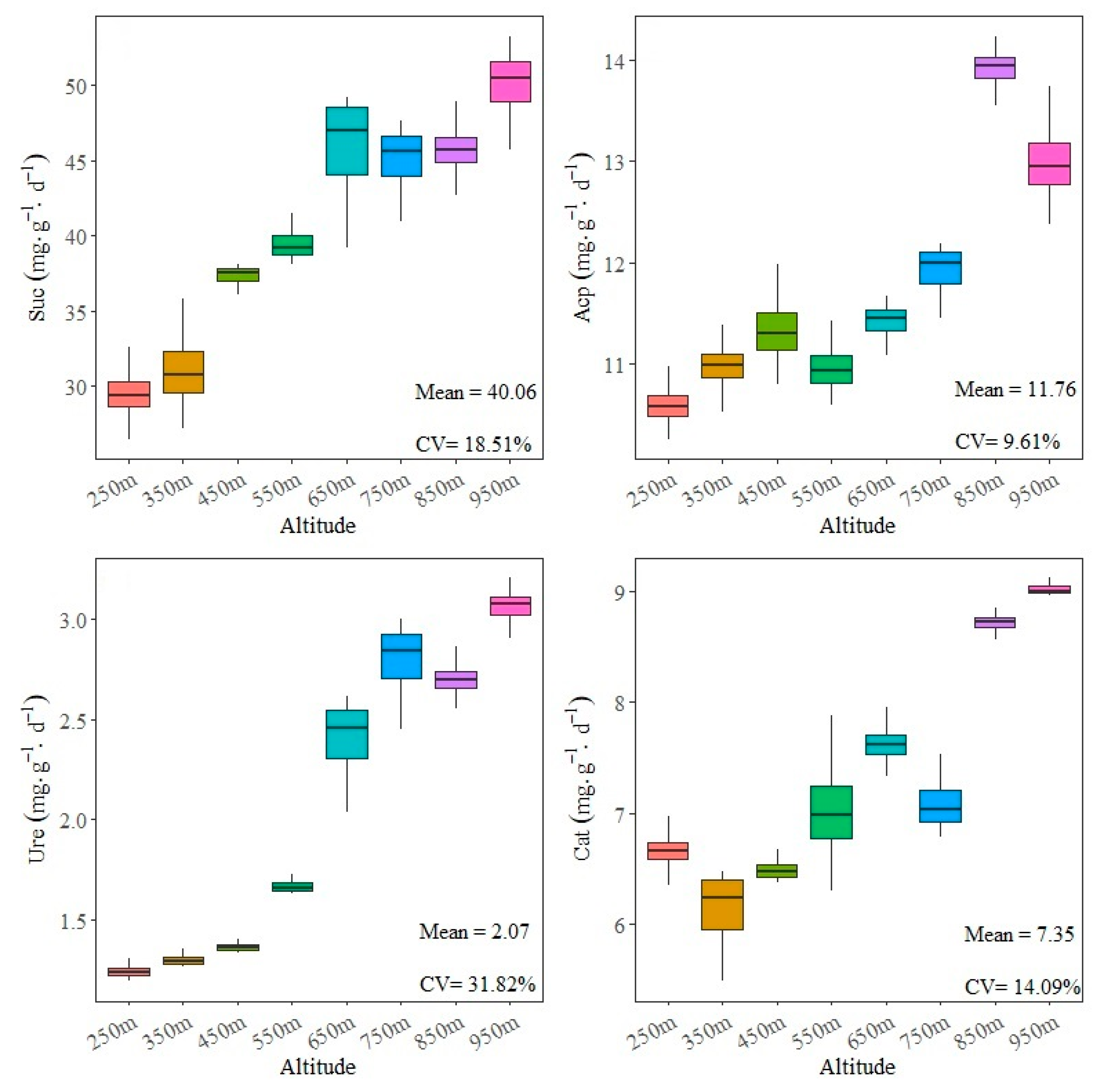
3.3. Impacts of Elevation on Soil Enzyme Activities
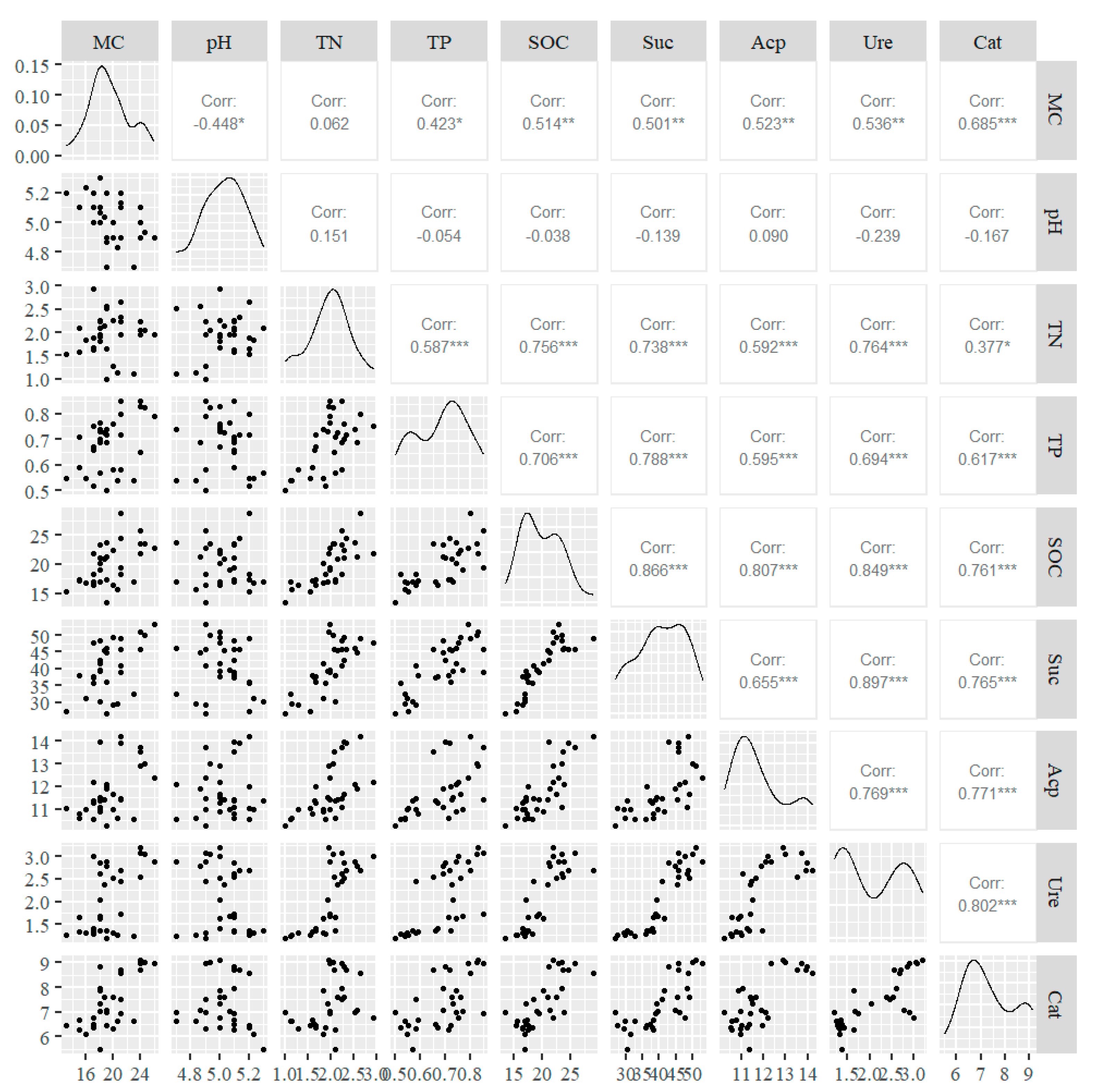
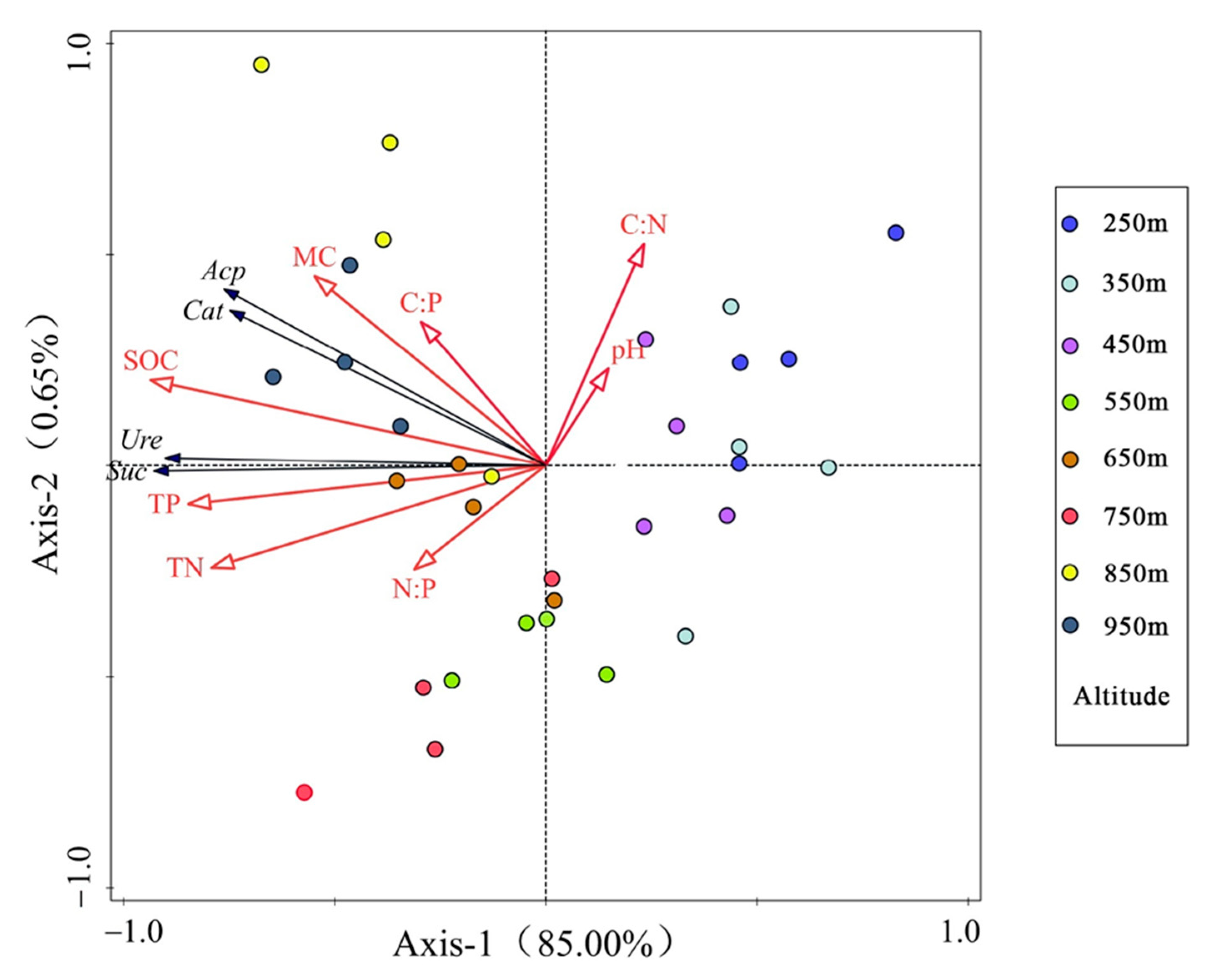
3.4. Redundancy Analysis of Soil Enzyme Activities and Environmental Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Pan, Y.; Zhou, Z.; Deng, J.; Zhao, F.; Guo, Y.; Han, X.; Yang, G.; Feng, Y.; Ren, G.; et al. Resource limitation and modeled microbial metabolism along an elevation gradient. Catena 2022, 209, 105807. [Google Scholar] [CrossRef]
- Elser, J.J. Biological stoichiometry from genes to ecosystems: Ideas, plans, and realities. Integr. Comp. Biol. 2002, 42, 1226. [Google Scholar]
- Liu, J.; Chen, J.; Chen, G.; Guo, J.; Li, Y. Enzyme stoichiometry indicates the variation of microbial nutrient requirements at different soil depths in subtropical forests. PLoS One 2020, 15, e0220599. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, M.; Huang, Z.; Lin, T.C.; Vadeboncoeur, M.A.; Searle, E.B.; Chen, H.Y.H. Temporal changes in soil C-N-P stoichiometry over the past 60 years across subtropical China. Glob. Change Biol. 2018, 24, 1308–1320. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.J.; Li, B.Y.; Gao, D.X.; Fu, S.Y.; Lu, Y.F.; Xu, M.P.; Ren, C.J.; Han, X.H. Response of Ecoenzymatic Stoichiometry to Soil Physicochemical Properties after Afforestation on Loess Hilly Region. Eurasian Soil Sci. 2020, 53, 1669–1675. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Feng, C.; Ma, Y.; Wang, W.; Huang, C.; Qi, C.; Fu, S.; Chen, H.Y.H. Transition from N to P limited soil nutrients over time since restoration in degraded subtropical broadleaved mixed forests. For. Ecol. Manag. 2021, 494, 119298. [Google Scholar] [CrossRef]
- Jiang, L.; He, Z.S.; Liu, J.F.; Xing, C.; Gu, X.G.; Wei, C.S.; Zhu, J.; Wang, X.L. Elevation Gradient Altered Soil C, N, and P Stoichiometry of Pinus taiwanensis Forest on Daiyun Mountain. Forests 2019, 10, 1089. [Google Scholar] [CrossRef] [Green Version]
- De Feudis, M.; Cardelli, V.; Massaccesi, L.; Lagomarsino, A.; Fornasier, F.; Westphalen, D.; Cocco, S.; Corti, G.; Agnelli, A. Influence of Altitude on Biochemical Properties of European Beech (Fagus sylvatica L.) Forest Soils. Forests 2017, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, C.; Wang, M.L. Linkages of C: N: P stoichiometry between soil and leaf and their response to climatic factors along altitudinal gradients. J. Soils Sed. 2019, 19, 1820–1829. [Google Scholar] [CrossRef]
- Aerts, R.; van Bodegom, P.M.; Cornelissen, J.H.C. Litter stoichiometric traits of plant species of high-latitude ecosystems show high responsiveness to global change without causing strong variation in litter decomposition. New Phytol. 2012, 196, 181–188. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Whitaker, J.; Ostle, N.J.; McNamara, N.P.; Bardgett, R.D.; Salinas, N.; Meir, P. Soil microbial nutrient constraints along a tropical forest elevation gradient: A belowground test of a biogeochemical paradigm. Biogeosciences 2015, 12, 6071–6083. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Li, D.D.; Jiao, F.; Yao, J.; Du, H.T. The Latitudinal Patterns of Leaf and Soil C:N:P Stoichiometry in the Loess Plateau of China. Front. Plant Sci. 2019, 10, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.X.; Dong, S.K.; Gao, Q.Z.; Zhou, H.K.; Liu, S.L.; Su, X.K.; Li, Y.Y. Effects of short-term and long-term warming on soil nutrients, microbial biomass and enzyme activities in an alpine meadow on the Qinghai-Tibet Plateau of China. Soil Biol. Biochem. 2014, 76, 140–142. [Google Scholar] [CrossRef]
- Hu, C.; Li, F.; Xie, Y.H.; Deng, Z.M.; Chen, X.S. Soil carbon, nitrogen, and phosphorus stoichiometry of three dominant plant communities distributed along a small-;scale elevation gradient in the East Dongting Lake. Phys. Chem. Earth 2018, 103, 28–34. [Google Scholar] [CrossRef]
- He, X.; Hou, E.; Liu, Y.; Wen, D. Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China. Sci. Rep. 2016, 6, 24261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Wang, J.J.; Wang, H.L.; Deng, P.F.; Li, A.Q.; Zhang, S.S.; Xu, X.N. Fine-root chemical traits rather than morphological traits of Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) plantations vary along an altitudinal gradient in Eastern China. For. Syst. 2022, 31, 4. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Z.C.; Ren, X.L.; Ma, X.M.; Zhou, M.Y.; Ge, X.; Liu, H.; Fu, S.L. Evaluation of Soil Quality in a Composite Pecan Orchard Agroforestry System Based on the Smallest Data Set. Sustainability 2022, 14, 10665. [Google Scholar] [CrossRef]
- Dominguez, M.T.; Holthof, E.; Smith, A.R.; Koller, E.; Emmett, B.A. Contrasting response of summer soil respiration and enzyme activities to long-term warming and drought in a wet shrubland (NE Wales, UK). Appl. Soil Ecol. 2017, 110, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wu, J.; Li, G.; Yan, L. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Su, Y.Q.; Wu, Z.L.; Xie, P.Y.; Zhang, L.; Chen, H. Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape. Forests 2020, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; An, S.S. Responses of soil nitrogen, phosphorous and organic matter to vegetation succession on the Loess Plateau of China. J. Arid Land 2015, 7, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Tessier, J.T. Vernal photosynthesis and nutrient retranslocation in Dryopteris intermedia. Am. Fern J. 2001, 91, 187–196. [Google Scholar] [CrossRef]
- Lie, Z.; Huang, W.; Liu, X.; Zhou, G.; Yan, J.; Li, Y.; Huang, C.; Wu, T.; Fang, X.; Zhao, M.; et al. Warming leads to more closed nitrogen cycling in nitrogen-rich tropical forests. Glob. Change Biol. 2021, 27, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Hui, D.F.; Yang, X.T.; Deng, Q.; Liu, Q.; Wang, X.; Yang, H.; Ren, H. Soil C:N:P stoichiometry in tropical forests on Hainan Island of China: Spatial and vertical variations. Catena 2021, 201, 105228. [Google Scholar] [CrossRef]
- McClaran, M.P.; Moore-Kucera, J.; Martens, D.A.; van Haren, J.; Marsh, S.E. Soil carbon and nitrogen in relation to shrub size and death in a semi-arid grassland. Geoderma 2008, 145, 60–68. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Jiang, L.Z.; Ushio, M.; Kitayama, K. Changes of soil chemical properties, microbial biomass and enzymatic activities along a gradient of forest degradation in logged over tropical rain forests, Borneo. Plant Soil 2022, 12. [Google Scholar] [CrossRef]
- Kooch, Y.; Mehr, M.A.; Hosseini, S.M. The effect of forest degradation intensity on soil function indicators in northern Iran. Ecol. Indic. 2020, 114, 106324. [Google Scholar] [CrossRef]
- Ndossi, E.M.; Becker, J.N.; Hemp, A.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Effects of land use and elevation on the functional characteristics of soil enzymes at Mt. Kilimanjaro. Eur. J. Soil Biol. 2020, 97, 103167. [Google Scholar] [CrossRef]
- Cao, R.; Yang, W.Q.; Chang, C.H.; Wang, Z.; Wang, Q.; Li, H.; Tan, B. Differential seasonal changes in soil enzyme activity along an altitudinal gradient in an alpine-gorge region. Appl. Soil Ecol. 2021, 166, 104078. [Google Scholar] [CrossRef]
- Wang, X.F.; Li, J.L.; Xing, G.T.; Mai, S.W.; Liu, W.J.; Jiang, Y.M.; Xu, W.X.; Yang, Q.; Yang, H.; Lu, J.L.; et al. Soil Organic Carbon Distribution, Enzyme Activities, and the Temperature Sensitivity of a Tropical Rainforest in Wuzhishan, Hainan Island. Forests 2022, 13, 1943. [Google Scholar] [CrossRef]
- Jin, K.; Sleutel, S.; Buchan, D.; De Neve, S.; Cai, D.X.; Gabriels, D.; Jin, J.Y. Changes of soil enzyme activities under different tillage practices in the Chinese Loess Plateau. Soil Tillage Res. 2009, 104, 115–120. [Google Scholar] [CrossRef]
- Cao, Y.E.; Wang, Y.; Xu, Z.L. Soil C:P ratio along elevational gradients in Picea schrenkiana forest of Tianshan Mountains. Pol. J. Ecol. 2018, 66, 325–336. [Google Scholar] [CrossRef]
- Zuccarini, P.; Asensio, D.; Ogaya, R.; Sardans, J.; Penuelas, J. Effects of seasonal and decadal warming on soil enzymatic activity in a P-deficient Mediterranean shrubland. Glob. Change Biol. 2020, 26, 3698–3714. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistic | Eigenvalues | Cumulative Explained Variation (%) | Pseudo-Canonical Correlation | Cumulative Explained Fitted Variation (%) |
|---|---|---|---|---|
| Axis 1 | 0.850 | 85.00 | 0.932 | 99.00 |
| Axis 2 | 0.006 | 85.65 | 0.637 | 99.75 |
| Axis 3 | 0.002 | 85.82 | 0.623 | 99.94 |
| Axis 4 | 0.001 | 85.87 | 0.561 | 100.00 |
| Name | Order of Importance | Explains (%) | Contribution (%) | pseudo-F | p-Value |
|---|---|---|---|---|---|
| SOC | 1 | 74.5 | 86.7 | 87.5 | 0.002 |
| TP | 2 | 5.9 | 6.9 | 8.8 | 0.002 |
| pH | 3 | 2.0 | 2.4 | 3.4 | 0.066 |
| N:P | 4 | 1.2 | 1.4 | 1.8 | 0.208 |
| C:N | 5 | 0.8 | 0.9 | 1.3 | 0.284 |
| MC | 6 | 0.6 | 0.7 | 1.0 | 0.316 |
| C:P | 7 | 0.6 | 0.7 | 1.0 | 0.308 |
| TN | 8 | 0.3 | 0.3 | 0.5 | 0.558 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Zhang, M.; Zuo, R.; Pu, F.; Feng, C.; Fu, S.; Liu, H. Altitudinal Gradients Modify the Ecological Stoichiometry and Enzyme Activities of Soil in a Natural Secondary Quercus spp. Forest of the Dabie Mountains. Forests 2023, 14, 774. https://doi.org/10.3390/f14040774
Huang C, Zhang M, Zuo R, Pu F, Feng C, Fu S, Liu H. Altitudinal Gradients Modify the Ecological Stoichiometry and Enzyme Activities of Soil in a Natural Secondary Quercus spp. Forest of the Dabie Mountains. Forests. 2023; 14(4):774. https://doi.org/10.3390/f14040774
Chicago/Turabian StyleHuang, Cheng, Manru Zhang, Ruitao Zuo, Faguang Pu, Chun Feng, Songling Fu, and Hua Liu. 2023. "Altitudinal Gradients Modify the Ecological Stoichiometry and Enzyme Activities of Soil in a Natural Secondary Quercus spp. Forest of the Dabie Mountains" Forests 14, no. 4: 774. https://doi.org/10.3390/f14040774