
Thinning Levels of Laurel Natural Regeneration to Establish Traditional Agroforestry Systems, Ecuadorian Amazon Upper Basin
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
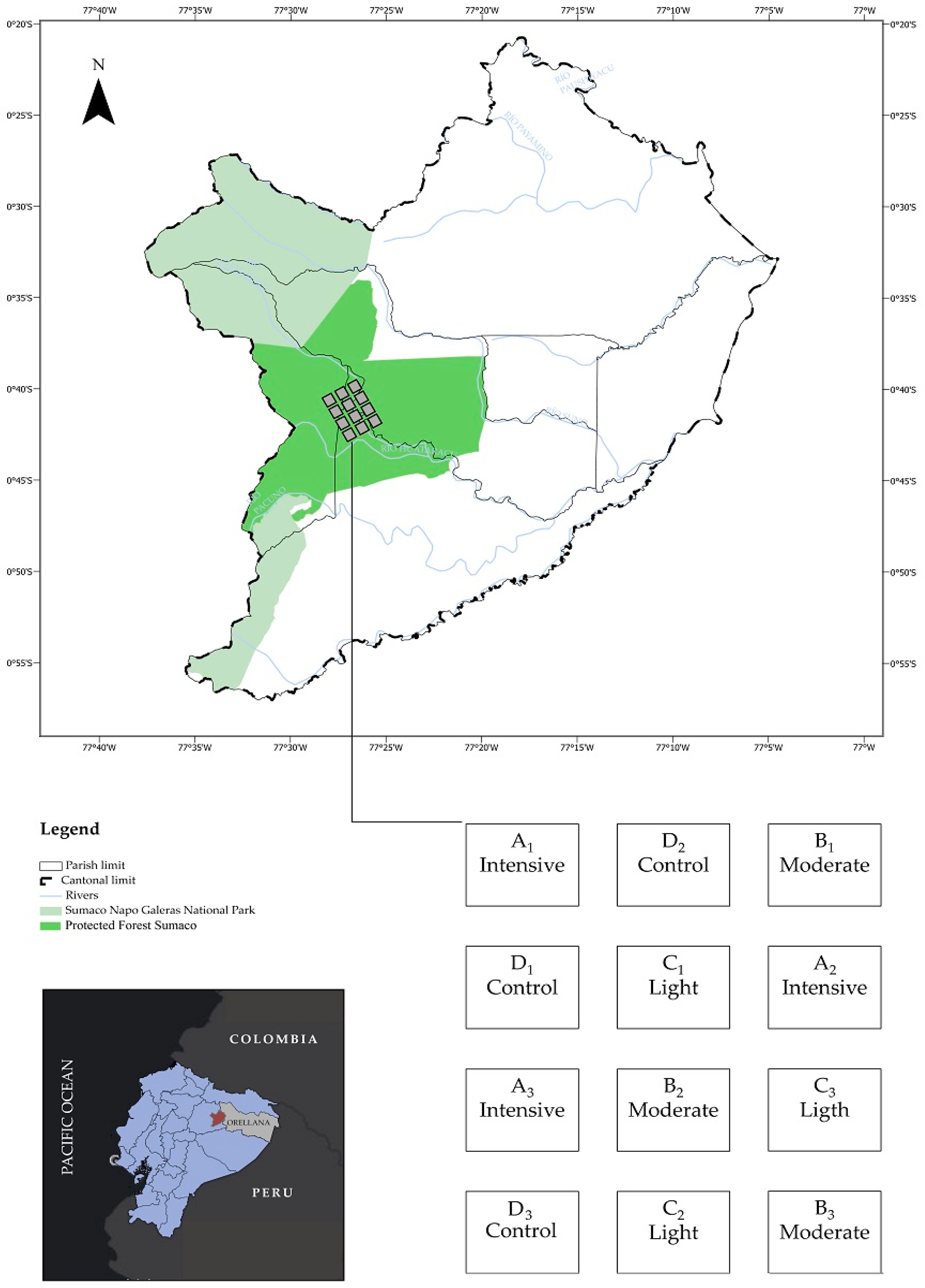
2.1. Location and Conditions of the Experimental Area
2.2. Statistical Design
2.3. Quantitative and Qualitative Thinning Indices
2.4. Information and Data Collection
2.5. Statistical Analyses
3. Results
3.1. Mean Plot Data by Thinnings Treatments
3.2. Quantitative and Qualitative of Thinnings Levels
3.3. Effects of Thinning Levels on Tree Parameters
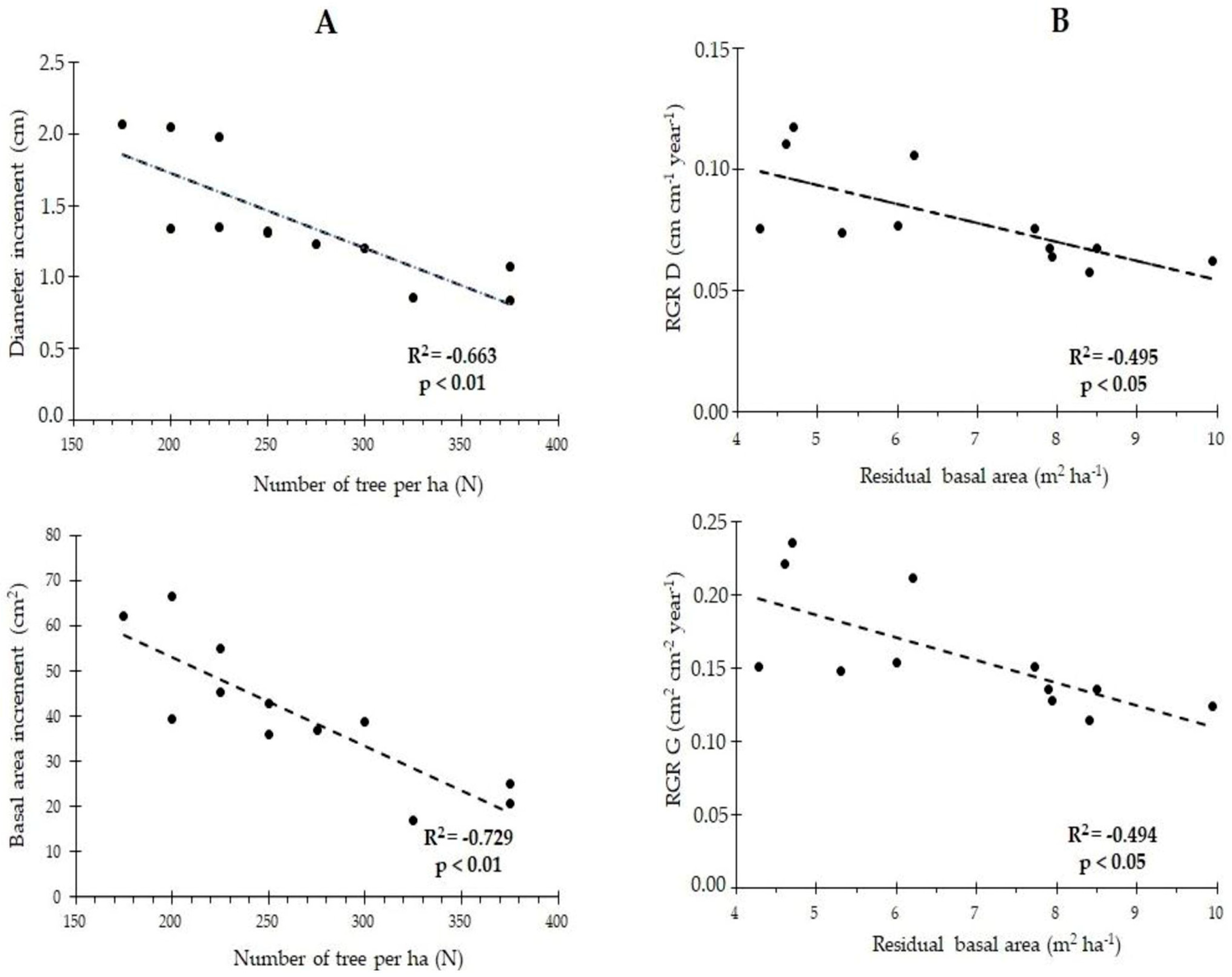
3.4. Relationship between Growth Rate and Plot Density in Terms of N and G
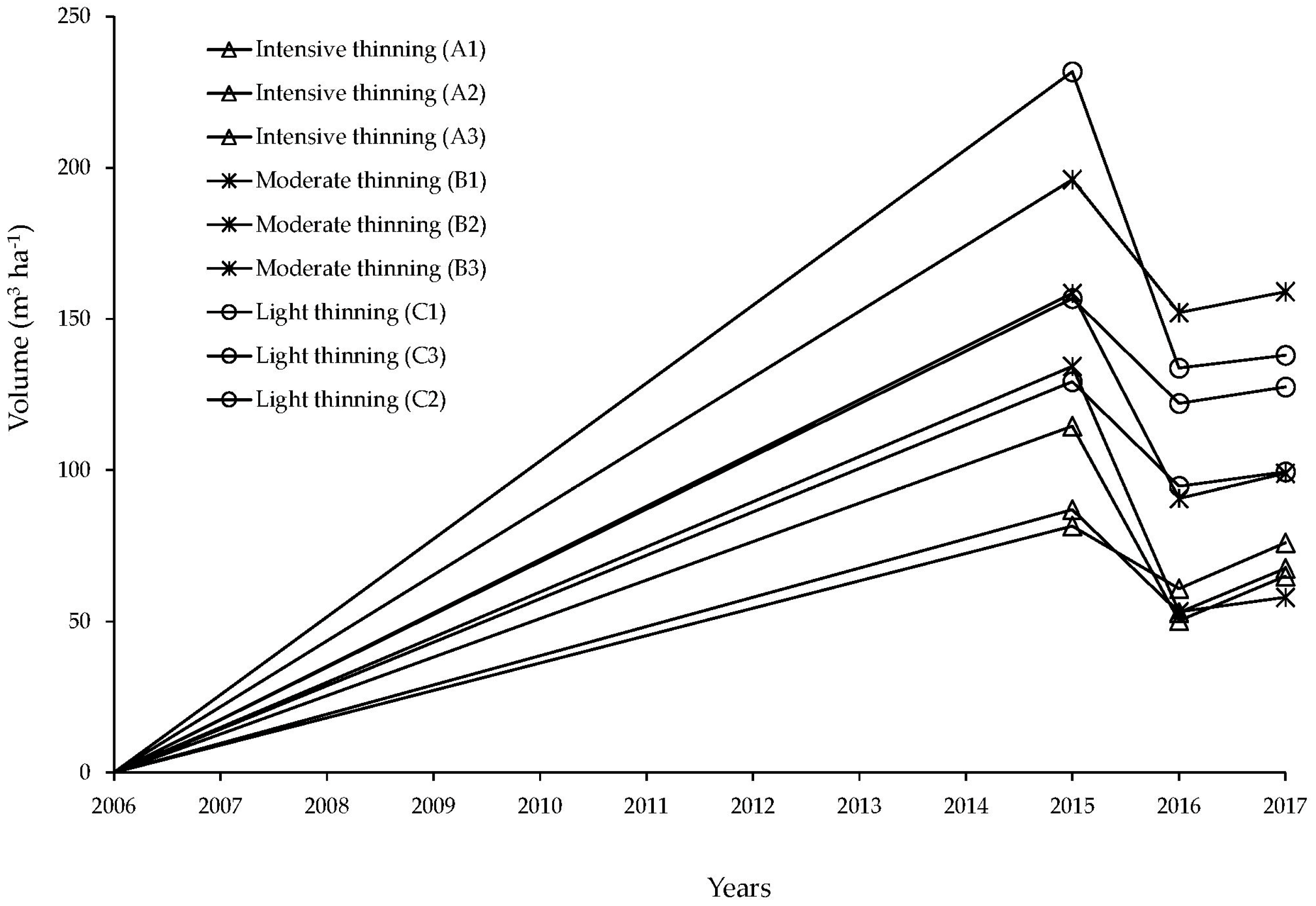
3.5. Growth after Thinning Treatments
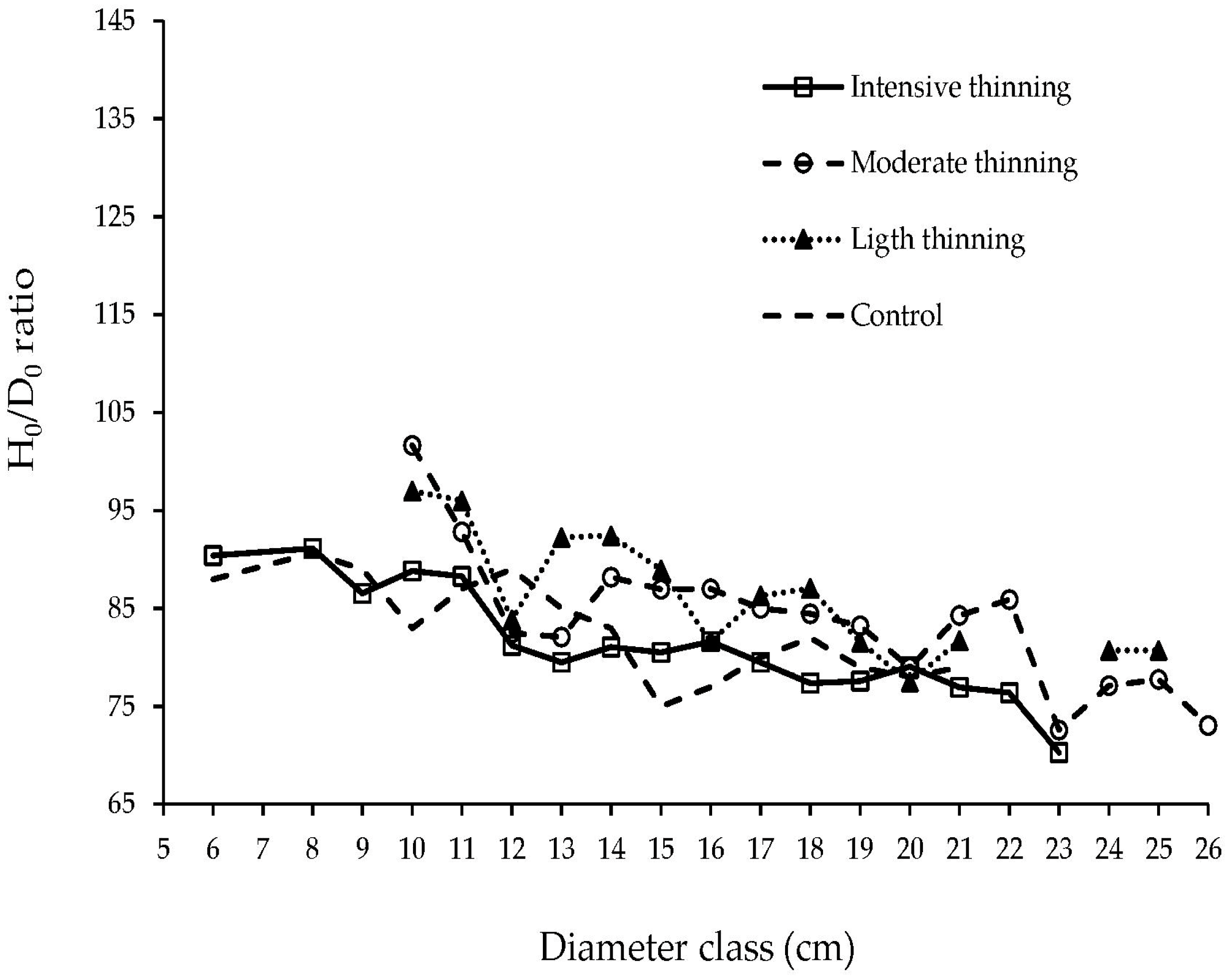
3.6. Height Diameter Relation
4. Discussion
4.1. Overall Production Observen in Thinning Plots
4.2. Quantitative and Qualitative Thinning Weights
4.3. Effects of Thinning Level on Tree Growth
4.4. Linear Relation of Growth Rate in DBH and Basal Area with Stand Density in Terms of N and G
4.5. Growth and Tree Density
4.6. Height Diameter Relation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. State of the World’s Forests 2011; FAO: Roma, Italy, 2011; ISBN 978-92-5-106750-5. [Google Scholar]
- MAE. Deforestación del Ecuador Continental Periodo 2014–2016; Ministry of Environment: Quito, Ecuador, 2017; pp. 1–37.
- Cañadas-López, A.; Arias, P.; Rade-Loor, D. Land Use/Cover Change (1997–2007) in the Protected Forest Sumaco, Northern Ecuadorian Amazon. In Biophysical and Socio-Economic Frame Conditions for the Sustainable Management of Natural Resources: International Research on Food Security, Natural Resource Management and Rural Development, 1st ed.; Tielkes, E., Ed.; DIRSL Gmbh Verlag: Hamburg, Germany, 2014; p. 225. [Google Scholar]
- Cañadas-López, A. Volumen y perfil de fuste para Cordia alliodora en sistemas agroforestales del bosque protector Sumaco. Bosques Latid. Cero 2007, 3, 15–20. [Google Scholar]
- Cañadas-López, Á.; Korol, M.; Zambrano, C.; Camacho, B. Modelos descriptivos de corona para Laurel (Cordia alliodora) bajo sistemas agroforestales en el Bosque Protector Sumaco, Ecuador. Av. Cien. Ing. 2012, 4, 31–38. [Google Scholar] [CrossRef]
- Hoch, L.; Pokorny, B.; De Jong, W. Financial attractiveness of smallholder tree plantations in the Amazon: Bridging external expectations and local realities. Agrofor. Syst. 2012, 84, 361–375. [Google Scholar] [CrossRef]
- Cañadas-López, Á.; Vilcko, F.; Rade-Loor, D.; Zambrano, C.; Molina, C. Hacia una descripción de fuste para el Laurel Cordia alliodora en sistemas agroforestales en el Bosque Protector Sumaco, Ecuador. Av. Cien. Ing. 2014, 6, 33–43. [Google Scholar] [CrossRef]
- Cañadas-López, Á.; Rade-Loor, D.; Siegmund-Schultze, M.; Moreira-Muñoz, G.; Vargas-Hernández, J.J.; Wehenkel, C. Growth and yield models for balsa wood plantations in the coastal lowlands of Ecuador. Forests 2019, 10, 733. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Making tropical succession and landscape reforestation successful. J. Sustain. Forest. 2013, 32, 649–658. [Google Scholar] [CrossRef]
- Wehenkel, C.; Corral-Rivas, J.J.; Hernández-Díaz, J.C. Genetic diversity in relation to secondary succession of forest tree communities. Pol. J. Ecol. 2011, 59, 45–54. [Google Scholar]
- Poorter, L.; Bongers, F.; Aide, T.M.; Almeyda Zambrano, A.M.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass resilience of Neotropical secondary forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef]
- Hérault, B.; Piponiot, C. Key drivers of ecosystem recovery after disturbance in a neotropical forest. For. Ecosyst. 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Rocha, G.P.; Vieira, D.L.; Simon, M.F. Fast natural regeneration in abandoned pastures in southern Amazonia. For. Ecol. Manag. 2016, 370, 93–101. [Google Scholar] [CrossRef]
- Cañadas, L. El Mapa Ecológico y Bioclimático del Ecuador; Editores Asociados Cia. Ltd.: Quito, Ecuador, 1983; pp. 50–120. ISBN 978-9942-09360-8. [Google Scholar]
- Cañadas-López, Á.; Rade-Loor, D.; Andrade-Candell, J.; Hernández-Díaz, J.C.; Molina Hidrovo, C.; Zambrano-Zambrano, M.; Wehenkel, C. Gap edge attributes in Neotropical rainforest, Ecuador. Rev. Biol. Trop. 2018, 66, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Parresol, B.R.; Devall, M.S. Patterns of diametric growth in stem-analyzed laurel trees (Cordia alliodora) in a Panamanian forest. Southwest Nat. 2013, 58, 170–178. [Google Scholar] [CrossRef]
- Schlönvoigt, A.; Beer, J. Initial growth of pioneer timber tree species in a Taungya system in the humid lowlands of Costa Rica. Agrofor. Syst. 2001, 51, 97–108. [Google Scholar] [CrossRef]
- Somarriba, E. Sustainable timber production from uneven-aged shade stands of Cordia alliodora in small coffee farms. Agrofor. Syst. 1990, 10, 253–263. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; Delgado Monsanto, L.; de Oliveira Neto, S.N.; Mota Cancio, N. La agroforestería como estrategia para la recuperación y conservación de reservas de carbono en bosques de la Amazonía. Bosque 2015, 36, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Hummel, S. Height, diameter, and crown dimensions of Cordia alliodora associated with tree density. For. Ecol. Manag. 2000, 127, 31–40. [Google Scholar] [CrossRef]
- Somarriba, E.; Suárez-Islas, A.; Calero-Borge, W.; Villota, A.; Castillo, C.; Vílchez, S.; Deheuvels, O.; Cerda, R. Cocoa–timber agroforestry systems: Theobroma cacao–Cordia alliodora in Central America. Agrofor. Syst. 2014, 88, 1001–1019. [Google Scholar] [CrossRef] [Green Version]
- Cañadas-López, A. Providing Information about Natural Resources as a Base to Support the Decentralization of the Forest Sector in Canton Loreto; Research Centre of Forest Ecology, Edition 192, Line A: Göttingen, Germany, 2005; pp. 65–75. [Google Scholar]
- Stromgaard, P. Biomass estimation equations for miombo woodland, Zambia. Agrofor. Syst. 1985, 3, 3–13. [Google Scholar] [CrossRef]
- Schmitt-Harsh, M.; Evans, T.P.; Castellanos, E.; Randolph, J.C. Carbon stocks in coffee agroforests and mixed dry tropical forests in the western highlands of Guatemala. Agrofor. Syst. 2012, 86, 141–157. [Google Scholar] [CrossRef]
- Cañadas, A.; Rade, D.; Domínguez, J.M.; Murillo, I.; Molina, C. Modelación Forestal Como Innovación Tecnológica Para el Manejo Silvicultural y Aprovechamiento Económico de la Balsa, Región Costa-Ecuador; Ed. Abya-Yala: Quito, Ecuador, 2016; pp. 20–40. ISBN 978-9942-09360-8. [Google Scholar]
- Birhane, E.; Ahmed, S.; Hailemariam, M.; Negash, M.; Rannestad, M.M.; Norgrove, L. Carbon stock and woody species diversity in homegarden agroforestry along an elevation gradient in southern Ethiopia. Agrofor. Syst. 2020, 94, 1099–1110. [Google Scholar] [CrossRef]
- Staupendahl, K.; Puumalainen, J. Modellierung des Einflusses von Durchforstungen auf die Durchmesserverteilung von gleichaltrigen Fichtenreibeständen. Forstwiss. Cent. Bl. 2000, 116, 249–262. [Google Scholar]
- Alder, D.; Montenegro, F.A. A yield model for Cordia alliodora plantation in Ecuador. Int. Forest. Rev. 1999, 1, 242–250. [Google Scholar]
- Heuveldop, J.; Fassbender, H.W.; Alpizar, L.; Enriquez, G.; Fölster, H. Modelling agroforestry systems of cacao (Theobroma cacao) with laurel (Cordia alliodora) and poro (Erythrina poeppigiana) in Costa Rica II. Cacao and wood production, litter production and decomposition. Agrofor. Syst. 1988, 6, 37–48. [Google Scholar] [CrossRef]
- Somarriba, E.J.; Beer, J.W. Dimensions, volumes and growth of Cordia alliodora in agroforestry systems. For. Ecol. Manag. 1987, 18, 113–126. [Google Scholar] [CrossRef]
- Peck, R.B.; Bishop, J.P. Management of secondary tree species in agroforestry systems to improve production sustainability in Amazonian Ecuador. Agrofor. Syst. 1992, 17, 53–63. [Google Scholar] [CrossRef]
- Cañadas-López, A.; Gamboa-Trujillo, P.; Buitrón-Garrido, S.; Medina-Torres, B.; Hernández-Vargas, J.J.; Wehenkel, C. Laurel regeneration management by smallholders to generate agroforestry systems in the Ecuadorian Amazon upper basin: Growth and yield models. Forests, 2023; summited. [Google Scholar]
- Summers, P.M.; Browder, J.O.; Pedlowski, M.A. Tropical forest management and silvicultural practices by small farmers in the Brazilian Amazon: Recent farm-level evidence from Rondonia. For. Ecol. Manag. 2004, 192, 161–177. [Google Scholar] [CrossRef]
- Simmons, C.S.; Perz, S.; Pedlowski, M.A.; Silva, L.G.T. The changing dynamics of land conflict in the Brazilian Amazon: The rural-urban complex and its environmental implications. Urban Ecosys. 2002, 6, 99–121. [Google Scholar] [CrossRef]
- Beer, J.W.; Clarkin, K.L.; De las Salas, G.; Glover, N.L. A case study of traditional agroforestry practices in a wet tropical zone: The ‘La Suiza’ project. In Las Ciencias Forestales y su Contribución al Desarrollo de la America Tropical, 1st ed.; Chavarria, M., Ed.; Conicit-Interciencia-Scitec: San José, Costa Rica, 1981; pp. 191–209. [Google Scholar]
- Kapp, G.B.; Beer, J. A comparison of agrosilvicultural systems with plantation forestry in the Atlantic lowlands of Costa Rica. Agrofor. Syst. 1995, 32, 207–223. [Google Scholar] [CrossRef]
- Somarriba, E.; Valdivieso, R.; Vásquez, W.; Galloway, G. Survival, growth, timber productivity and site index of Cordia alliodora in forestry and agroforestry systems. Agrofor. Syst. 2001, 51, 111–118. [Google Scholar] [CrossRef]
- Abetz, P. Eine Entscheidungshilfe für die Durchforstung von Fichtenbeständen. AFZ 1975, 30, 666–667. [Google Scholar]
- Kramer, H. Waldwachstumlehre, 1st ed.; Kessel Verlag: Hamburg/Berlin, Germany, 1988; pp. 35–80. [Google Scholar]
- Hunt, R. Basic Growth Analysis. Plant Growth Analysis for Beginners, 1st ed.; Unwin Hyman: London, UK, 1990; pp. 20–60. [Google Scholar]
- Pommerening, A.; Muszta, A. Methods of modelling relative growth rate. For. Ecosyst. 2015, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Kramer, H.; Röös, M. Druchforstungversuch in einem weitständig begründeten Kiefernbestand. Forst. Holz. 1989, 44, 139–144. [Google Scholar]
- Montero, G.; Cañellas, I.; Ortega, C.; Del Rio, M. Results from a thinning experiment in a Scots pine (Pinus sylvestris L.) natural regeneration stand in the Sistema Ibérico Mountain Range (Spain). For. Ecol. Manag. 2001, 145, 151–161. [Google Scholar] [CrossRef]
- Vega, L. Plantaciones de Cordia alliodora en Combinación con Cultivos Agrícolas: Una Alternativa de Manejo en Surinam; IFLAIC Bol.53: Mérida, Venezuela, 1978; pp. 21–38. [Google Scholar]
- Piotto, D.; Montagnini, F.; Ugalde, L.; Kanninen, M. Performance of forest plantations in small and medium-sized farms in the Atlantic lowlands of Costa Rica. For. Ecol. Manag. 2003, 175, 195–204. [Google Scholar] [CrossRef]
- Drew, T.J.; Flewelling, J.W. Stand density management: An alternative approach and its application to Douglas-fir plantations. For. Sci. 1979, 25, 518–532. [Google Scholar] [CrossRef]
- Kohyama, T.S.; Potts, M.D.; Kohyama, T.I.; Kassim, A.R.; Ashton, P.S. Demographic properties shape tree size distribution in a Malaysian rain forest. Am. Nat. 2015, 185, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Weiner, J. Asymmetric competition in plant populations. Trends Evol. Evol. 1990, 5, 360–364. [Google Scholar] [CrossRef]
- Kohyama, T.; Takada, T. The stratification theory for plant coexistence promoted by one-sided competition. J. Ecol. 2009, 97, 463–471. [Google Scholar] [CrossRef]
- Hummel, S.S. Stand Development of Cordia alliodora (Boraginaceae), a Neotropical Secondary Forest Tree in Northern Costa Rica (1992–1996). Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1998. [Google Scholar]
- Dawkins, H.C. Crown diameters: Their relation to bole diameter in tropical forest trees. Commonw. For. Rev. 1963, 42, 318–333. [Google Scholar]
- Grau, O.; Peñuelas, J.; Ferry, B.; Freycon, V.; Blanc, L.; Desprez, M.; Baraloto, C.; Chave, J.; Descroix, L.; Dourdain, A.; et al. Nutrient-cycling mechanisms other than the direct absorption from soil may control forest structure and dynamics in poor Amazonian soils. Sci. Rep. 2017, 7, 45017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negrón-Juárez, R.I.; Holm, J.A.; Marra, D.M.; Rifai, S.W.; Riley, W.J.; Chambers, J.Q.; Koven, C.D.; Knox, R.G.; McGroddy, M.E.; Di Vittorio, A.V. Vulnerability of Amazon forests to storm-driven tree mortality. Environ. Res. Lett. 2018, 13, 054021. [Google Scholar] [CrossRef]
- Marchi, M.; Paletto, A.; Cantiani, P.; Bianchetto, E.; De Meo, I. Comparing thinning system effects on ecosystem services provision in artificial black pine (Pinus nigra JF Arnold) forests. Forests 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Fortin, M.; Van Couwenberghe, R.; Perez, V.; Piedallu, C. Evidence of climate effects on the height-diameter relationships of tree species. Ann. For. Sci. 2019, 76, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Balandier, P.; Dupraz, C. Growth of widely spaced trees. A case study from young agroforestry plantations in France. Agrofor. Syst. 1998, 43, 151. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand before Thinning | Tree Removed | Stand after Thinning | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tr | N | Do | Ho | Dg | Hg | G | V | N | Do | Ho | Dg | Hg | G | V | N | Do | Ho | Dg | Hg | G | V |
| A1 | 450 | 24.0 | 18.4 | 17.3 | 14.4 | 11.2 | 114.6 | 275 | 24.0 | 18.4 | 16.8 | 14.0 | 6.6 | 64.4 | 175 | 22.4 | 18.2 | 18.1 | 15.1 | 4.6 | 50.2 |
| A2 | 450 | 22.7 | 19.5 | 15.5 | 12.6 | 9.0 | 87.0 | 225 | 14.2 | 12.0 | 14.2 | 12.0 | 4.3 | 34.1 | 225 | 22.7 | 19.5 | 16.7 | 13.2 | 4.7 | 52.9 |
| A3 | 425 | 24.3 | 18.5 | 15.5 | 10.9 | 10.0 | 81.5 | 225 | 17.0 | 13.0 | 11.9 | 9.1 | 3.8 | 20.7 | 200 | 24.3 | 18.5 | 19.5 | 12.8 | 6.2 | 60.8 |
| B1 | 500 | 23.2 | 18.5 | 16.0 | 13.8 | 11.3 | 158.5 | 250 | 19.5 | 18.0 | 15.0 | 13.5 | 5.3 | 67.9 | 250 | 23.2 | 18.5 | 17.0 | 14.1 | 6.0 | 90.6 |
| B2 | 450 | 26.7 | 21.0 | 20.6 | 16.9 | 12.3 | 196.1 | 225 | 17.7 | 20.1 | 15.1 | 15.0 | 4.4 | 44.0 | 225 | 26.7 | 21.0 | 21.0 | 16.5 | 7.9 | 152.1 |
| B3 | 450 | 24.8 | 18.0 | 18.4 | 14.5 | 18.0 | 134.3 | 250 | 24.8 | 18.0 | 19.1 | 14.5 | 7.3 | 81.1 | 200 | 23.7 | 17.5 | 18.1 | 14.4 | 5.3 | 53.2 |
| C1 | 450 | 26.4 | 20.0 | 18.6 | 16.2 | 13.0 | 156.8 | 150 | 26.4 | 20.0 | 15.8 | 15.0 | 3.1 | 34.7 | 300 | 26.4 | 20.0 | 20.0 | 16.8 | 9.9 | 122.1 |
| C2 | 425 | 28.5 | 22.0 | 19.5 | 16.0 | 13.4 | 231.8 | 150 | 28.5 | 22.0 | 21.1 | 16.2 | 5.5 | 98.0 | 275 | 25.8 | 20.1 | 18.7 | 15.9 | 7.9 | 133.8 |
| C3 | 400 | 29.4 | 19.0 | 19.0 | 13.8 | 12.2 | 129.3 | 150 | 25.0 | 15.5 | 17.2 | 12.0 | 3.7 | 34.6 | 250 | 29.4 | 19.0 | 20.1 | 14.9 | 8.5 | 94.7 |
| D1 | 375 | 25.9 | 18.9 | 16.4 | 12.8 | 7.6 | 84.2 | ||||||||||||||
| D2 | 350 | 21.1 | 17.8 | 15.7 | 12.5 | 6.5 | 73.7 | ||||||||||||||
| D3 | 300 | 21.2 | 19 | 12.7 | 10.4 | 3.7 | 55.5 | ||||||||||||||
| Plot | Quantitative Weights (%) | Qualitative Weight | ||
|---|---|---|---|---|
| TN | TG | TV | NG | |
| A1 | 61.11 | 58.93 | 57.46 | 1.04 |
| A2 | 50.00 | 47.83 | 45.69 | 1.05 |
| A3 | 52.94 | 37.80 | 32.77 | 1.40 |
| Average: | 54.68 | 48.19 | 45.31 | 1.16 |
| B1 | 50.00 | 46.90 | 46.98 | 1.07 |
| B2 | 50.00 | 35.77 | 32.74 | 1.40 |
| B3 | 55.56 | 58.02 | 57.92 | 0.96 |
| Average: | 51.85 | 46.90 | 39.86 | 1.14 |
| C1 | 33.33 | 23.92 | 25.35 | 1.39 |
| C2 | 35.29 | 41.19 | 39.30 | 0.86 |
| C3 | 37.50 | 30.33 | 27.91 | 1.24 |
| Average: | 35.37 | 31.81 | 30.85 | 1.16 |
| Thinning Weights | D2016 (cm) | AGR (cm year−1) | RGR (cm cm−1 year−1) | ||
|---|---|---|---|---|---|
| DBH (cm) | Basal Area (cm2) | DBH (cm) | Basal Area (cm2) | ||
| A | 18.12 a | 2.03 a | 61.37 a | 0.111 a | 0.223 a |
| B | 18.71 a | 1.34 b | 40.26 b | 0.072 b | 0.143 b |
| C | 19.58 a | 1.25 b | 39.45 b | 0.066 b | 0.131 b |
| D (Control) | 14.28 b | 0.93 c | 20.95 c | 0.070 b | 0.139 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cañadas-López, Á.; Gamboa-Trujillo, P.; Buitrón-Garrido, S.; Medina-Torres, B.; Vargas-Hernández, J.J.; Wehenkel, C. Thinning Levels of Laurel Natural Regeneration to Establish Traditional Agroforestry Systems, Ecuadorian Amazon Upper Basin. Forests 2023, 14, 667. https://doi.org/10.3390/f14040667
Cañadas-López Á, Gamboa-Trujillo P, Buitrón-Garrido S, Medina-Torres B, Vargas-Hernández JJ, Wehenkel C. Thinning Levels of Laurel Natural Regeneration to Establish Traditional Agroforestry Systems, Ecuadorian Amazon Upper Basin. Forests. 2023; 14(4):667. https://doi.org/10.3390/f14040667
Chicago/Turabian StyleCañadas-López, Álvaro, Paul Gamboa-Trujillo, Santiago Buitrón-Garrido, Byron Medina-Torres, J. Jesús Vargas-Hernández, and Christian Wehenkel. 2023. "Thinning Levels of Laurel Natural Regeneration to Establish Traditional Agroforestry Systems, Ecuadorian Amazon Upper Basin" Forests 14, no. 4: 667. https://doi.org/10.3390/f14040667