
Influence of Earlier Snowmelt on the Seedling Growth of Six Subboreal Tree Species in the Spring


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plant Cultivation
2.3. Snowmelt Treatment
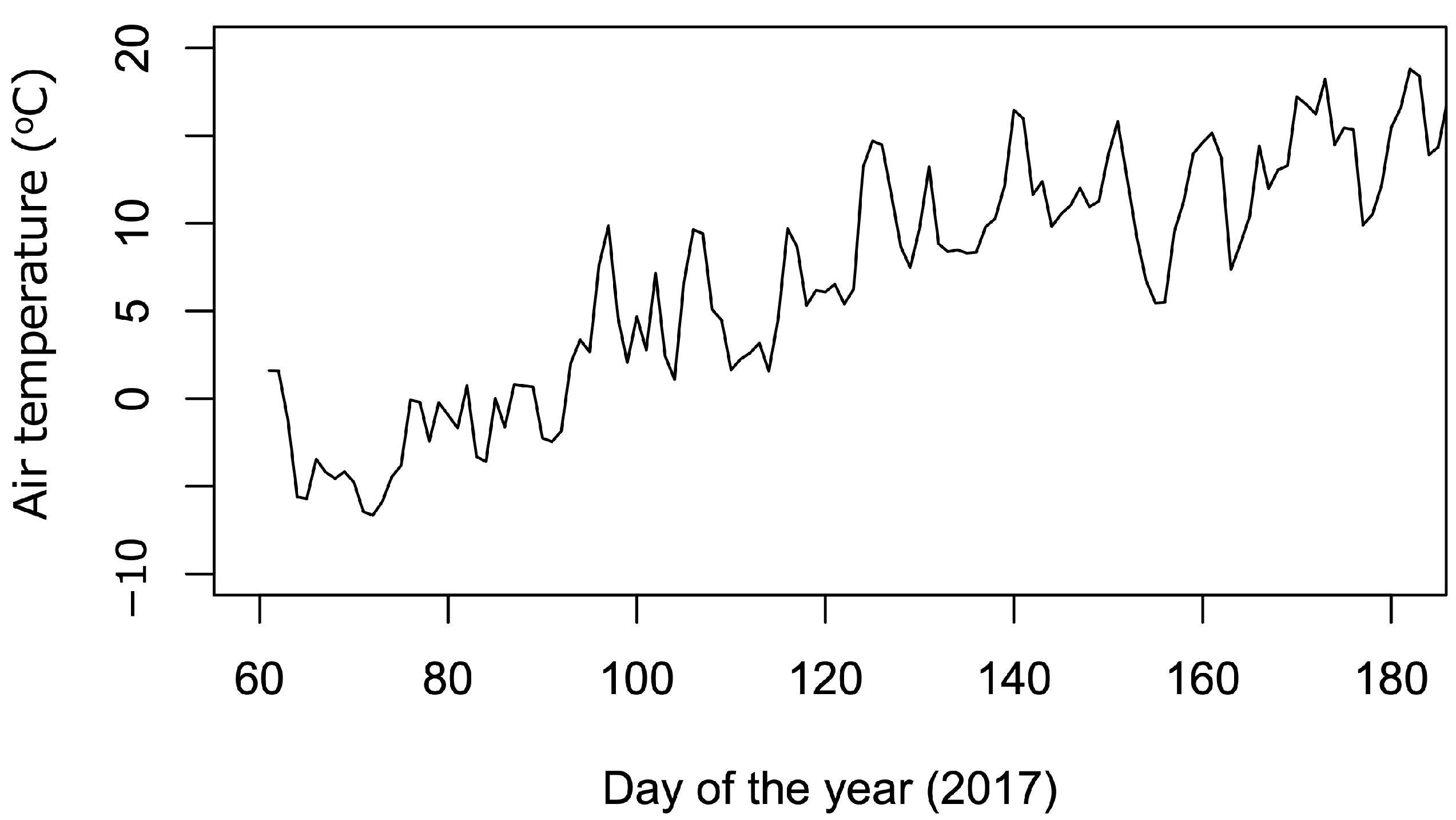
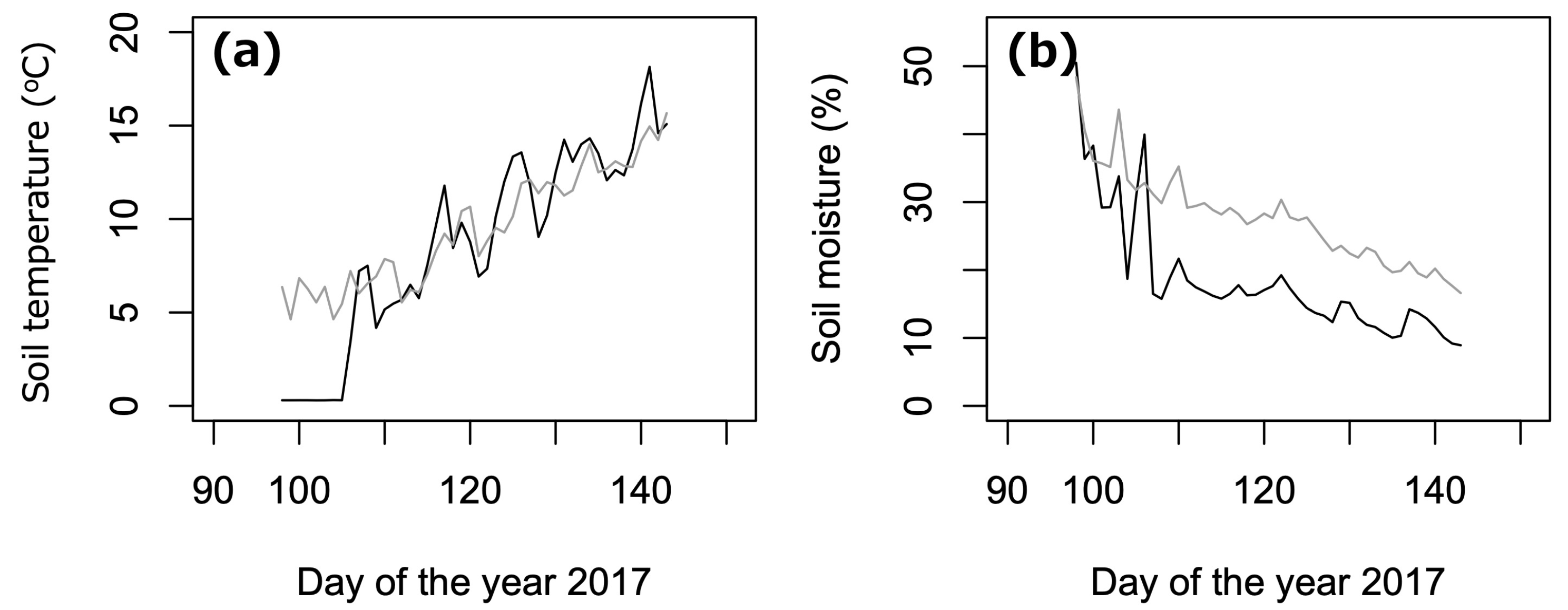
2.4. Environmental Measurements
2.5. Plant Growth
2.6. Phenology
2.7. Leaf Traits
2.8. Statistical Analysis
3. Results
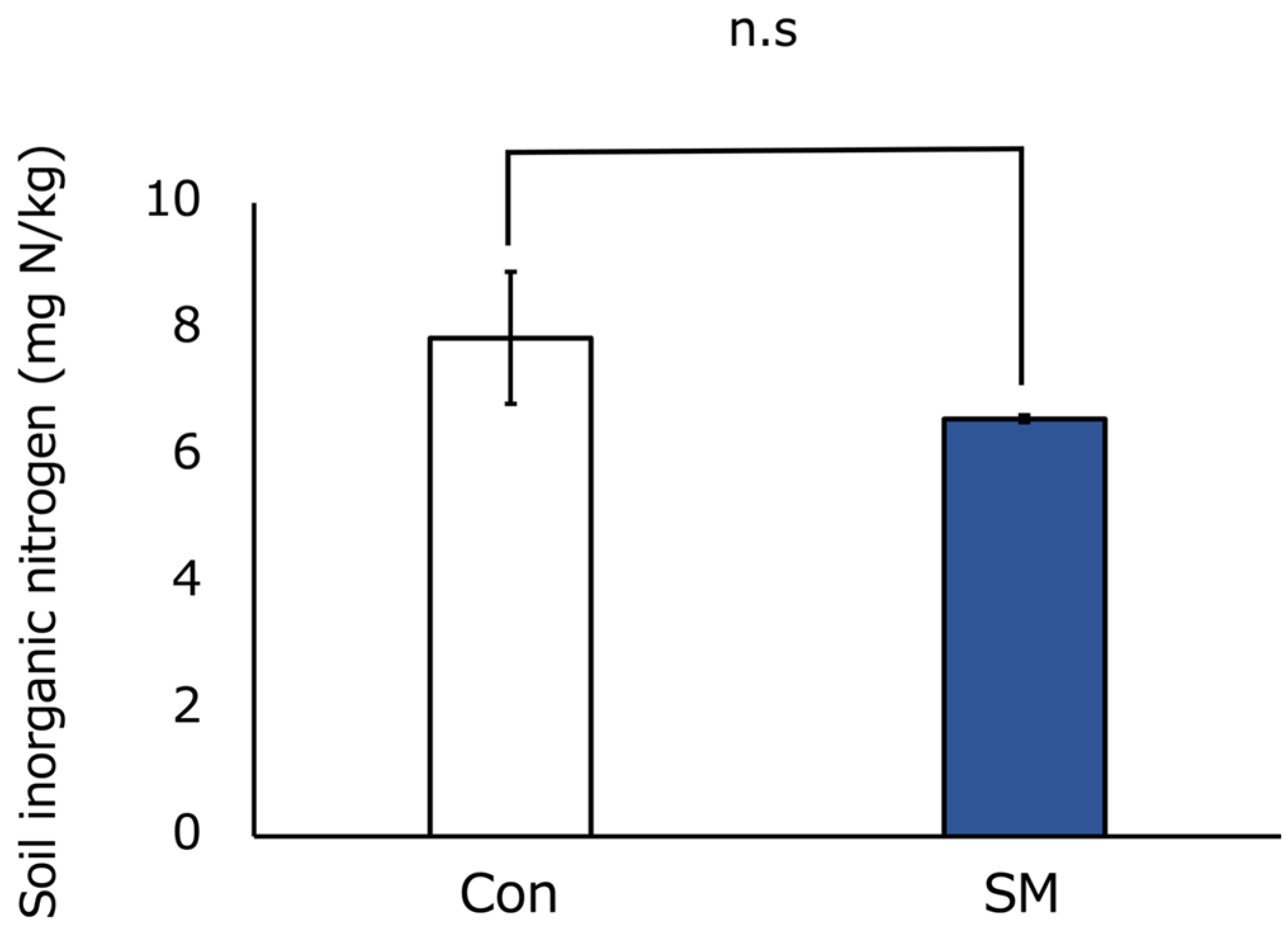
3.1. Abiotic Environment
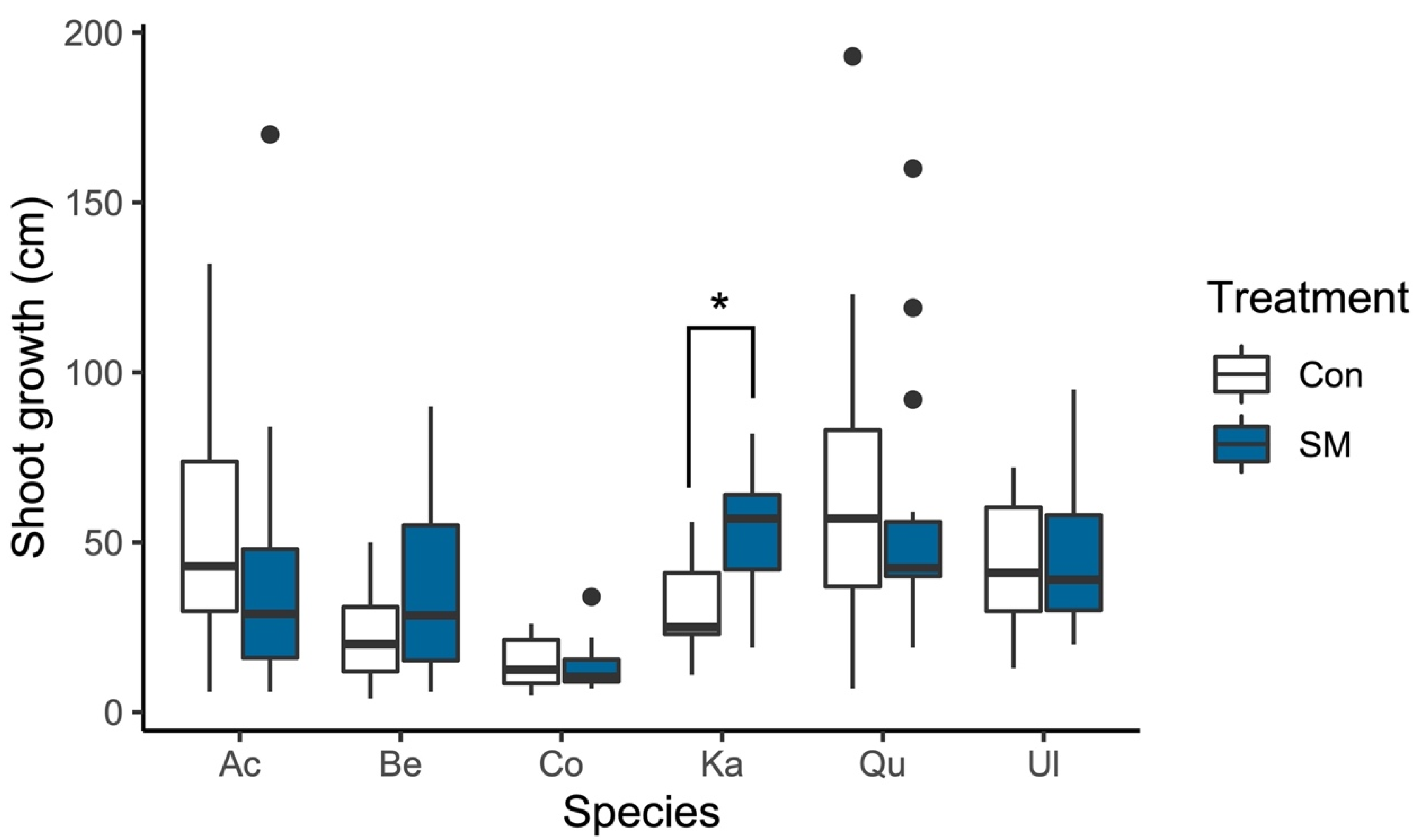
3.2. Shoot Growth
3.3. Phenology and Leaf Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Hayhoe, K.; Wake, C.P.; Huntington, T.G.; Luo, L.; Schwartz, M.D.; Sheffield, J.; Wood, E.; Anderson, B.; Bradbury, J.; DeGaetano, A.; et al. Past and Future Changes in Climate and Hydrological Indicators in the US Northeast. Clim. Dyn. 2007, 28, 381–407. [Google Scholar] [CrossRef]
- Liu, J.; Curry, J.A.; Dai, Y.; Horton, R. Causes of the Northern High-Latitude Land Surface Winter Climate Change. Geophys. Res. Lett. 2007, 34, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Makoto, K.; Kajimoto, T.; Koyama, L.; Kudo, G.; Shibata, H.; Yanai, Y.; Cornelissen, J.H.C. Winter Climate Change in Plant–Soil Systems: Summary of Recent Findings and Future Perspectives. Ecol. Res. 2014, 29, 593–606. [Google Scholar] [CrossRef]
- Makoto, K.; Templer, P.H.; Katayama, A.; Seki, O.; Takagi, K. Early Snowmelt by an Extreme Warming Event Affects Understory More than Overstory Trees in Japanese Temperate Forests. Ecosphere 2022, 13, e4182. [Google Scholar] [CrossRef]
- Chen, X.; Liang, S.; Cao, Y.; He, T.; Wang, D. Observed Contrast Changes in Snow Cover Phenology in Northern Middle and High Latitudes from 2001–2014. Sci. Rep. 2015, 5, 16820. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Larcher, W. Frost Survival of Plants; Springer: Berlin/Heidelberg, Germany, 1987; ISBN 9783642717475. [Google Scholar]
- Marumo, E.; Takagi, K.; Makoto, K. Timing of Bud Burst of Smaller Individuals Is Not Always Earlier than That of Larger Trees in a Cool-Temperate Forest with Heavy Snow. J. For. Res. 2020, 25, 285–290. [Google Scholar] [CrossRef]
- Makoto, K. Does Ontogeny Matter for the Spring Temperature Requirement for Bud Burst of Two Coniferous Species in Cool-Temperate Forests? Can. J. For. Res. 2022, 53, 203–209. [Google Scholar] [CrossRef]
- Weigel, R.; Henry, H.A.L.; Beil, I.; Gebauer, G.; Jurasinski, G.; Klisz, M.; van der Maaten, E.; Muffler, L.; Kreyling, J. Ecosystem Processes Show Uniform Sensitivity to Winter Soil Temperature Change Across a Gradient from Central to Cold Marginal Stands of a Major Temperate Forest Tree. Ecosystems 2021, 24, 1545–1560. [Google Scholar] [CrossRef]
- Little, C.J.; Wheeler, J.A.; Sedlacek, J.; Cortés, A.J.; Rixen, C. Small-Scale Drivers: The Importance of Nutrient Availability and Snowmelt Timing on Performance of the Alpine Shrub Salix herbacea. Oecologia 2016, 180, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Wipf, S.; Rixen, C. A Review of Snow Manipulation Experiments in Arctic and Alpine Tundra Ecosystems. Polar Res. 2010, 29, 95–109. [Google Scholar] [CrossRef]
- Bokhorst, S.; Bjerke, J.W.; Bowles, F.W.; Melillo, J.; Callaghan, T.V.; Phoenix, G.K. Impacts of Extreme Winter Warming in the Sub-Arctic: Growing Season Responses of Dwarf Shrub Heathland. Glob. Change Biol. 2008, 14, 2603–2612. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Meadows, M.W.; Lucas, R.G.; Hart, S.C. Snowmelt Timing Alters Shallow but Not Deep Soil Moisture in the Sierra Nevada. Water Resour. Res. 2014, 50, 1448–1456. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Makoto, K. Winter Climate Change, Plant Traits and Nutrient and Carbon Cycling in Cold Biomes. Ecol. Res. 2014, 29, 517–527. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A Handbook of Protocols for Standardised and Easy Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Frenette-Dussault, C.; Shipley, B.; Meziane, D.; Hingrat, Y. Trait-Based Climate Change Predictions of Plant Community Structure in Arid Steppes. J. Ecol. 2013, 101, 484–492. [Google Scholar] [CrossRef]
- Ladwig, L.M.; Chandler, J.L.; Guiden, P.W.; Henn, J.J. Extreme Winter Warm Event Causes Exceptionally Early Bud Break for Many Woody Species. Ecosphere 2019, 10, e02542. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and Consequences of Variation in Leaf Mass per Area (LMA): A Meta-Analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Kreyling, J.; Gebauer, G.; Klisz, M.; Weigel, R. 15N Tracer Enrichment in Response to Winter Soil Temperature Manipulation Differs between Canopy Trees and Juveniles. Trees-Struct. Funct. 2021, 35, 325–331. [Google Scholar] [CrossRef]
- Rosbakh, S.; Leingärtner, A.; Hoiss, B.; Krauss, J.; Steffan-Dewenter, I.; Poschlod, P. Contrasting Effects of Extreme Drought and Snowmelt Patterns on Mountain Plants along an Elevation Gradient. Front. Plant Sci. 2017, 8, 1478. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.T.; Gezon, Z.J. Plasticity in Functional Traits in the Context of Climate Change: A Case Study of the Subalpine Forb Boechera stricta (Brassicaceae). Glob. Change Biol. 2015, 21, 1689–1703. [Google Scholar] [CrossRef]
- Navarro, J.; Powers, J.M.; Paul, A.; Campbell, D.R. Phenotypic Plasticity and Selection on Leaf Traits in Response to Snowmelt Timing and Summer Precipitation. New Phytol. 2022, 234, 1477–1490. [Google Scholar] [CrossRef]
- Rixen, C.; Schwoerer, C.; Wipf, S. Winter Climate Change at Different Temporal Scales in Vaccinium myrtillus, an Arctic and Alpine Dwarf Shrub. Polar Res. 2010, 29, 85–94. [Google Scholar] [CrossRef]
- Livensperger, C.; Steltzer, H.; Darrouzet-Nardi, A.; Sullivan, P.F.; Wallenstein, M.; Weintraub, M.N. Earlier Snowmelt and Warming Lead to Earlier but Not Necessarily More Plant Growth. AoB Plants 2016, 8, plw021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiwa, K. Changes in Leaf Phenology Are Dependent on Tree Height in Acer Mono, a Deciduous Broad-Leaved Tree. Ann. Bot. 1999, 83, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Uemura, S. Patterns of Leaf Phenology in Forest Understory. Can. J. Bot. 1994, 72, 409–414. [Google Scholar] [CrossRef]
- Makoto, K.; Bryanin, S.V.; Takagi, K. The Effect of Snow Reduction and Eisenia japonica Earthworm Traits on Soil Nitrogen Dynamics in Spring in a Cool-Temperate Forest. Appl. Soil Ecol. 2019, 144, 1–7. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Satoh, F.; Koike, T.; Sasa, K. Vertical Distribution and Seasonal Pattern of Fine-Root Dynamics in a Cool-Temperate Forest in Northern Japan: Implication of the Understory Vegetation, Sasa Dwarf Bamboo. Ecol. Res. 2007, 22, 485–495. [Google Scholar] [CrossRef]
- Augspurger, C.K. Spring 2007 Warmth and Frost: Phenology, Damage and Refoliation in a Temperate Deciduous Forest. Funct. Ecol. 2009, 23, 1031–1039. [Google Scholar] [CrossRef]
- Hanba, Y.T.; Mori, S.; Lei, T.T.; Koike, T.; Wada, E. Variations in Leaf 13 C along a Vertical Profile of Irradiance in a Temperate Japanese Forest. Oecologia 1997, 110, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Peppe, D.J.; Royer, D.L.; Cariglino, B.; Oliver, S.Y.; Newman, S.; Leight, E.; Enikolopov, G.; Fernandez-Burgos, M.; Herrera, F.; Adams, J.M.; et al. Sensitivity of Leaf Size and Shape to Climate: Global Patterns and Paleoclimatic Applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Tateno, R.; Imada, S.; Fukuzawa, K.; Isobe, K.; Urakawa, R.; Oda, T.; Hosokawa, N.; Sasai, T.; Inagaki, Y.; et al. The Effect of a Freeze–Thaw Cycle on Dissolved Nitrogen Dynamics and Its Relation to Dissolved Organic Matter and Soil Microbial Biomass in the Soil of a Northern Hardwood Forest. Biogeochemistry 2019, 142, 319–338. [Google Scholar] [CrossRef]
- Zhang, X.; Manzanedo, R.D.; D’Orangeville, L.; Rademacher, T.T.; Li, J.; Bai, X.; Hou, M.; Chen, Z.; Zou, F.; Song, F.; et al. Snowmelt and Early to Mid-Growing Season Water Availability Augment Tree Growth during Rapid Warming in Southern Asian Boreal Forests. Glob. Change Biol. 2019, 25, 3462–3471. [Google Scholar] [CrossRef]
- Wang, X.; Ye, T.; Ata-Ul-Karim, S.T.; Zhu, Y.; Liu, L.; Cao, W.; Tang, L. Development of a Critical Nitrogen Dilution Curve Based on Leaf Area Duration in Wheat. Front. Plant Sci. 2017, 8, 1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kļaviņa, D.; Gaitnieks, T.; Menkis, A. Survival, Growth and Ectomycorrhizal Community Development of Container- and Bare-Root Grown Pinus Sylvestris and Picea Abies Seedlings Outplanted on a Forest Clear-Cut. Balt. For. 2013, 19, 39–49. [Google Scholar]
- Díaz, S.; Purvis, A.; Cornelissen, J.H.C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional Traits, the Phylogeny of Function, and Ecosystem Service Vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Wood Structure | Bud-Burst Day (DOY) | Leaf Area (m2) | LMA (g/m2) | LDMC | Leaf N (g/m2) | δ13C (‰) |
|---|---|---|---|---|---|---|---|
| Acer mono | Diffuse | 140.2 ± 4.5 | 0.12 ± 0.05 | 33.9 ± 14.4 | 0.37 ± 0.05 | 1.40 ± 0.56 | −27.5 ± 0.2 |
| Betula platyphylla | Diffuse | 131.3 ± 2.1 | 0.09 ± 0.02 | 49.9 ± 6.2 | 0.34 ± 0.08 | 2.05 ± 0.22 | −29.6 ± 0.3 |
| Cornus controversa | Diffuse | 136.0 ± 3.7 | 0.07 ± 0.05 | 44.0 ± 16.8 | 0.45 ± 0.11 | 1.90 ± 0.29 | −28.1 ± 1.8 |
| Kalopanax septemlobus | Ring | 140.1 ± 1.8 | 0.20 ± 0.12 | 56.7 ± 22.0 | 0.34 ± 0.07 | 2.57 ± 0.37 | −25.9 ± 0.4 |
| Quercus crispula | Ring | 142.2 ± 3.9 | 0.08 ± 0.04 | 40.1 ± 6.3 | 0.32 ± 0.08 | 1.75 ± 0.23 | −26.0 ± 0.8 |
| Ulmus davidiana | Ring | 134.5 ± 3.6 | 0.04 ± 0.01 | 43.5 ± 9.4 | 0.32 ± 0.07 | 2.06 ± 0.05 | −28.5 ± 0.5 |
| Species | Shoot Length | Bud-Burst Day | Leaf Area | LMA | LDMC | Leaf N | Leaf δ13C | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | p | Estimate | p | Estimate | p | Estimate | p | Estimate | p | Estimate | p | Estimate | p | |
| Acer mono | n.s. | −0.021 | p < 0.01 | n.s. | n.s. | n.s. | n.s. | n.s. | ||||||
| Betula platyphylla | n.s. | −0.021 | p < 0.01 | n.s. | n.s. | n.s. | n.s. | n.s. | ||||||
| Cornus controversa | n.s. | −0.025 | p < 0.01 | n.s. | n.s. | n.s. | n.s. | n.s. | ||||||
| Kalopanax septemlobus | 0.57 | p < 0.05 | −0.015 | p < 0.01 | 0.99 | p < 0.01 | −0.26 | p < 0.05 | n.s. | −0.37 | p < 0.05 | n.s. | ||
| Quercus crispula | n.s. | n.s. | n.s. | −0.15 | p < 0.05 | n.s. | n.s. | n.s. | ||||||
| Ulmus davidiana | n.s. | −0.015 | p < 0.01 | n.s. | n.s. | n.s. | n.s. | n.s. | ||||||
| Response Variable | Explanatory Variables | |||||
|---|---|---|---|---|---|---|
| Bud-Burst Day | Leaf Area | LMA | LDMC | Leaf N | Leaf δ¹3C | |
| Shoot length | Estimate: +46.1 | |||||
| n.s. | p < 0.05 | n.s. | n.s. | n.s. | n.s. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marumo, E.; Ueda, M.U.; Seki, O.; Takagi, K.; Makoto, K. Influence of Earlier Snowmelt on the Seedling Growth of Six Subboreal Tree Species in the Spring. Forests 2023, 14, 600. https://doi.org/10.3390/f14030600
Marumo E, Ueda MU, Seki O, Takagi K, Makoto K. Influence of Earlier Snowmelt on the Seedling Growth of Six Subboreal Tree Species in the Spring. Forests. 2023; 14(3):600. https://doi.org/10.3390/f14030600
Chicago/Turabian StyleMarumo, Erica, Miki U. Ueda, Osamu Seki, Kentaro Takagi, and Kobayashi Makoto. 2023. "Influence of Earlier Snowmelt on the Seedling Growth of Six Subboreal Tree Species in the Spring" Forests 14, no. 3: 600. https://doi.org/10.3390/f14030600