
Long-Term Climate Sensitivity of Resin-Tapped and Non-Resin-Tapped Scots Pine Trees Based on Tree Ring Width and Blue Intensity

Abstract
:1. Introduction
2. Materials and Methods
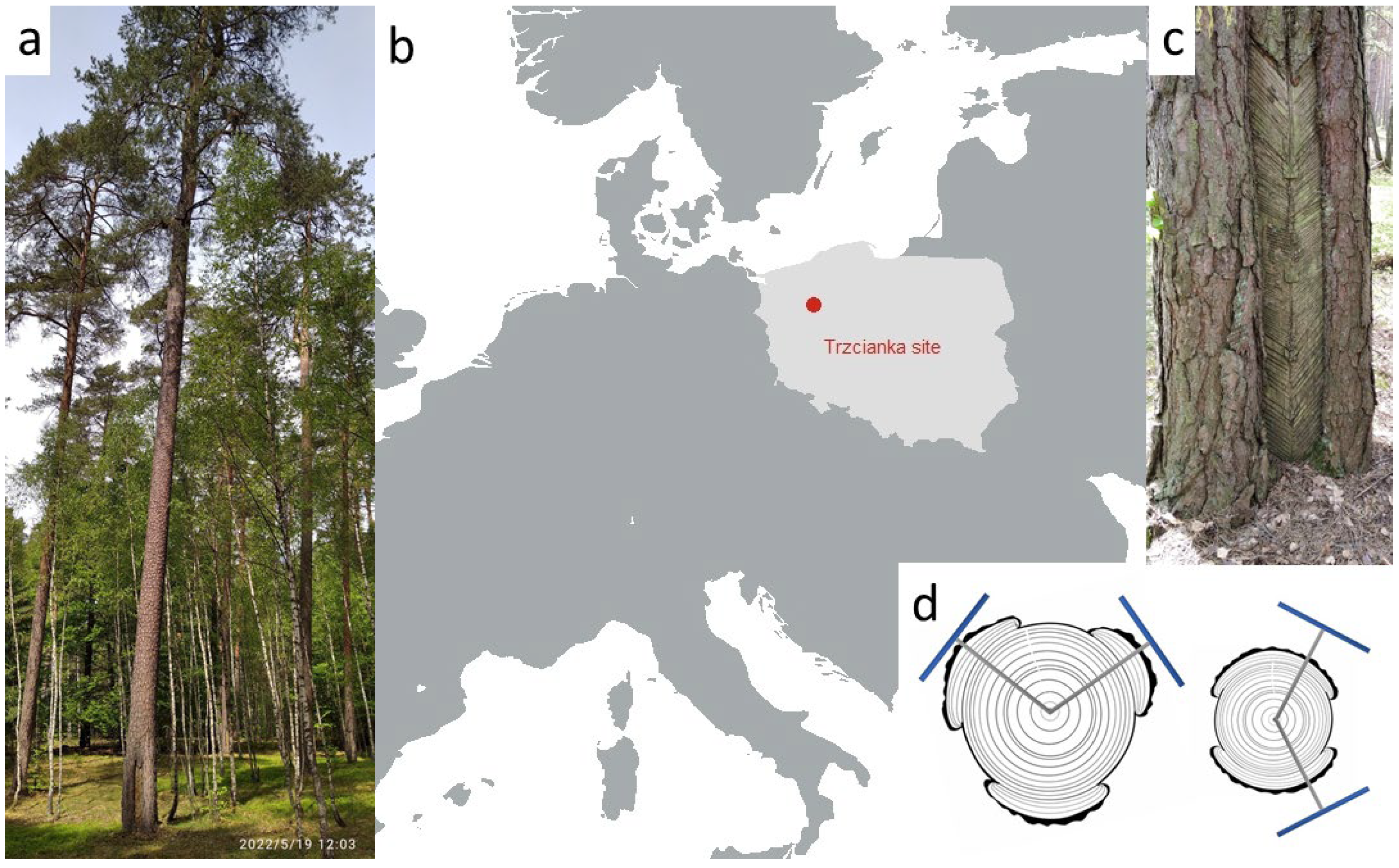
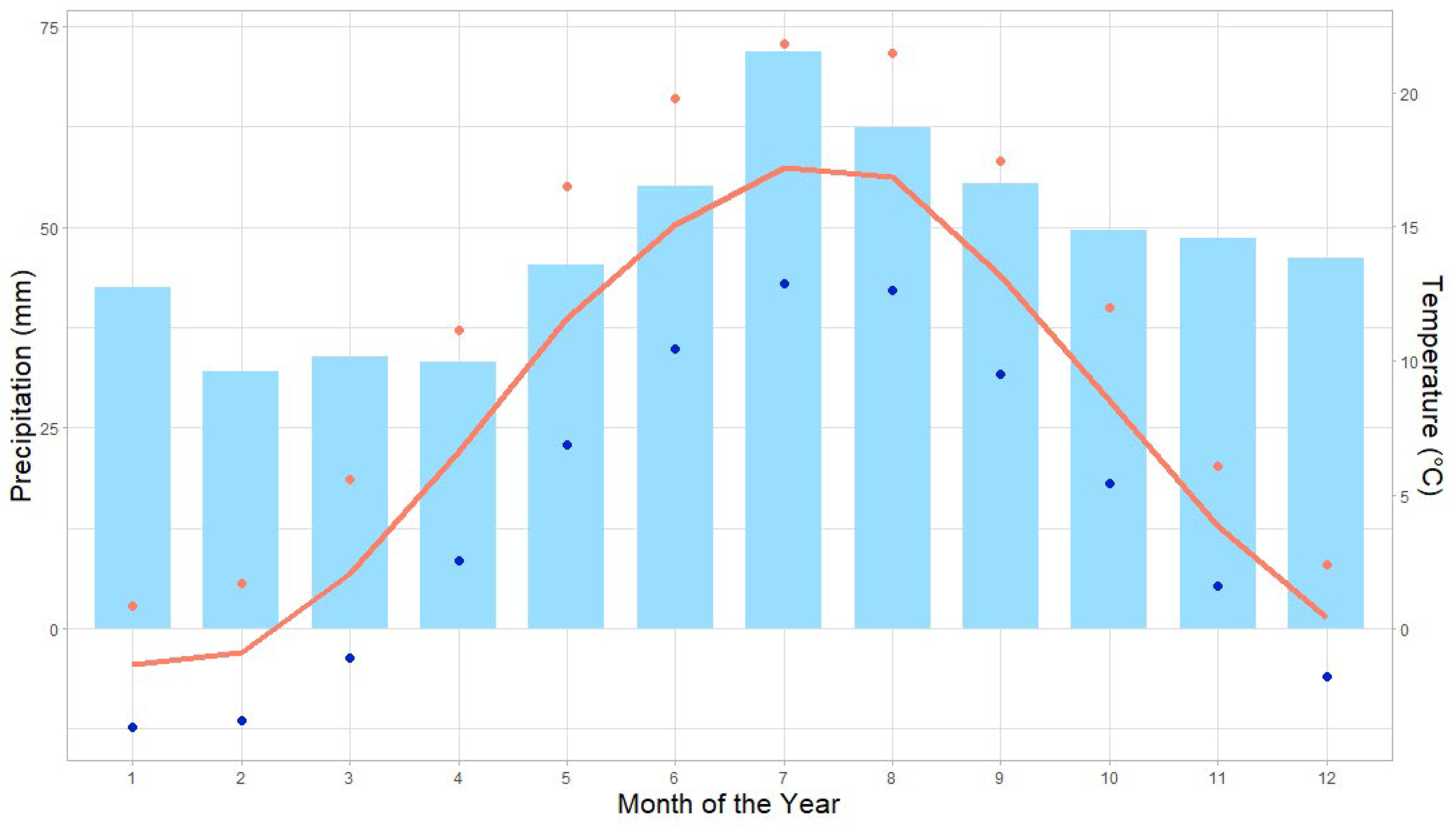
2.1. Research Area
2.2. Tree Coring and Sample Preparation
2.3. Climate–Signal Correlations
3. Results
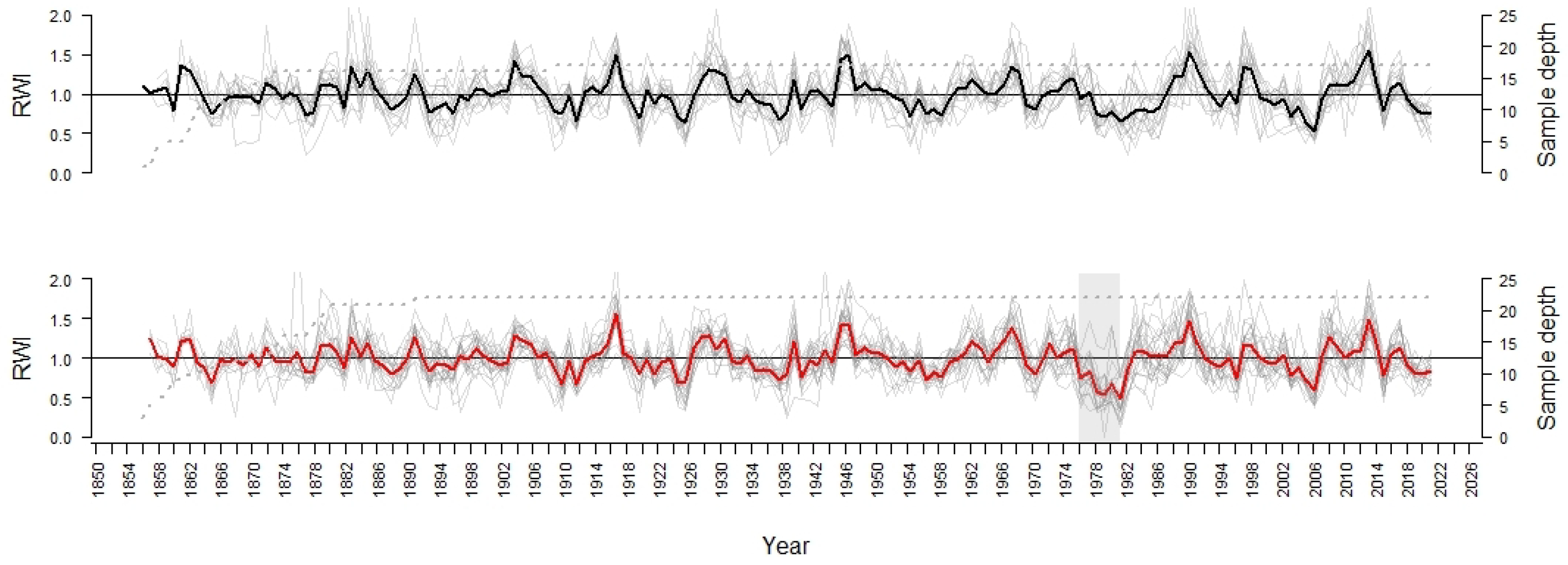
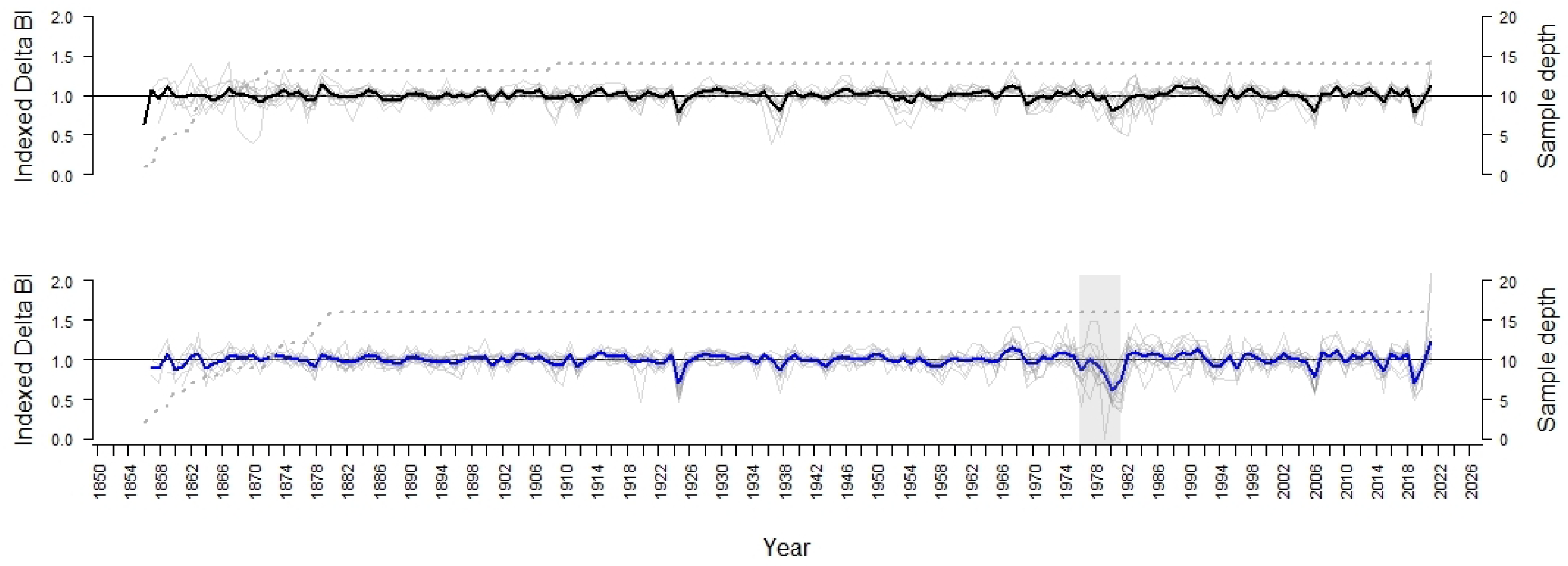
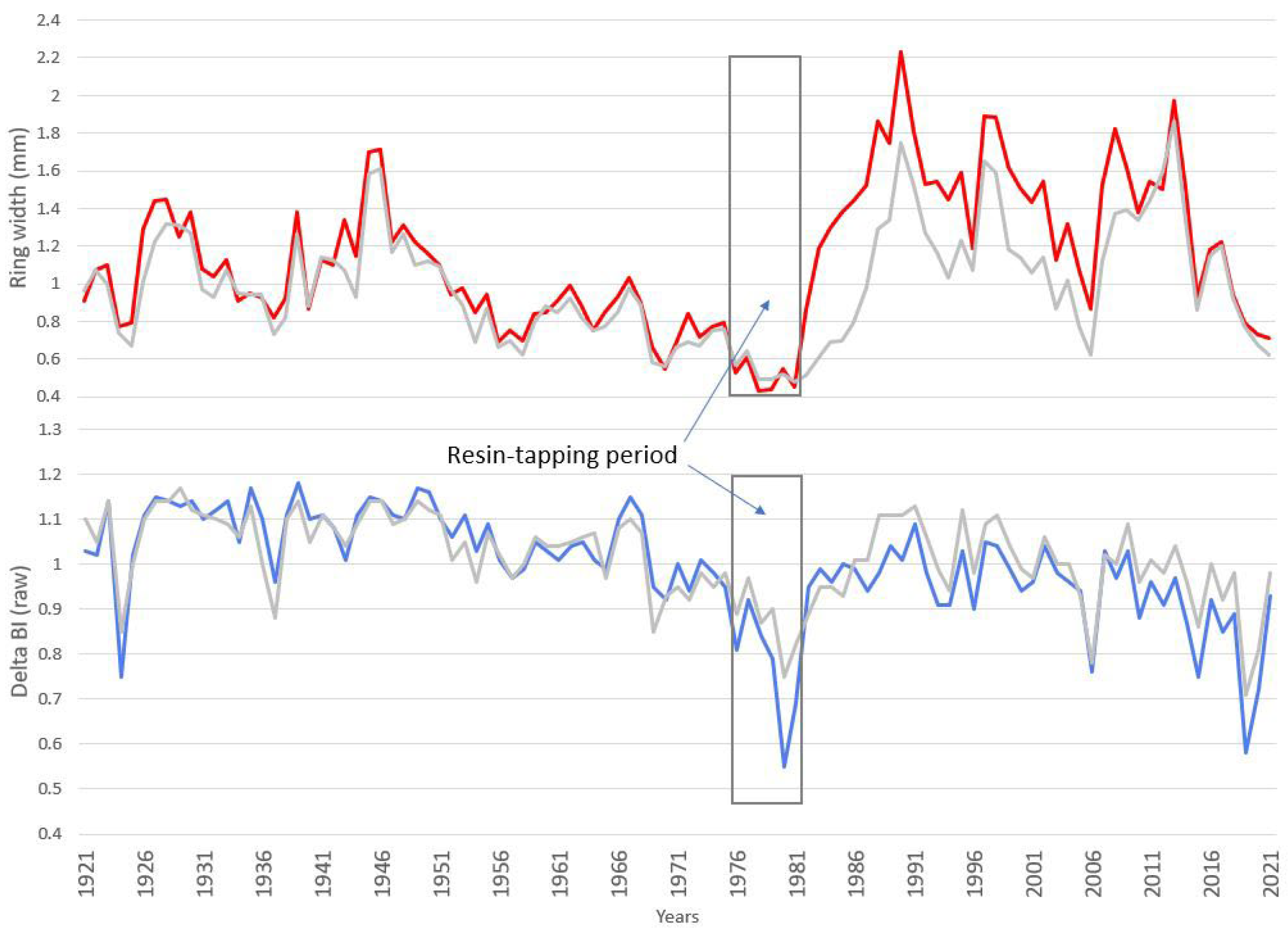
3.1. Comparison of the Chronologies
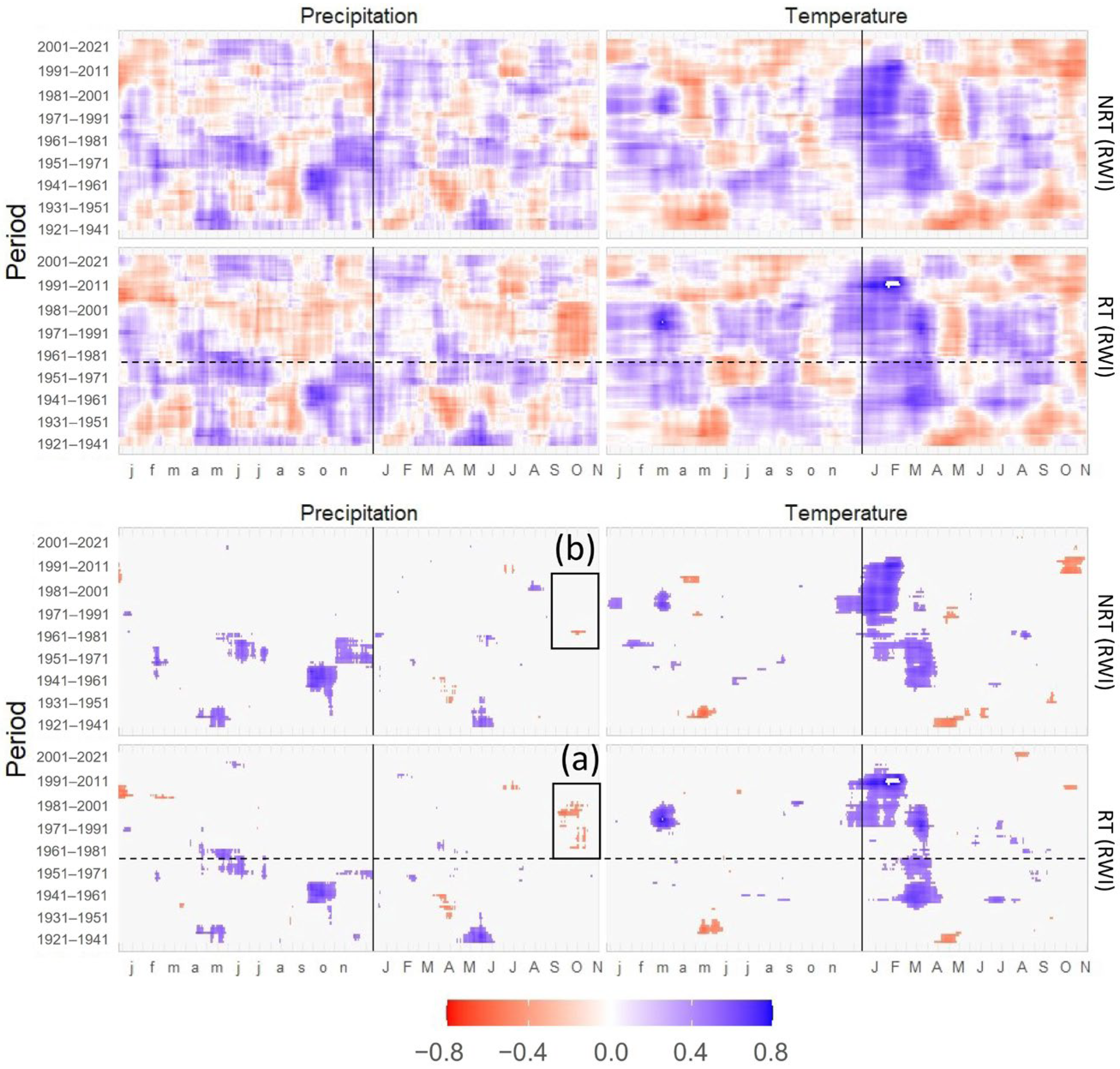
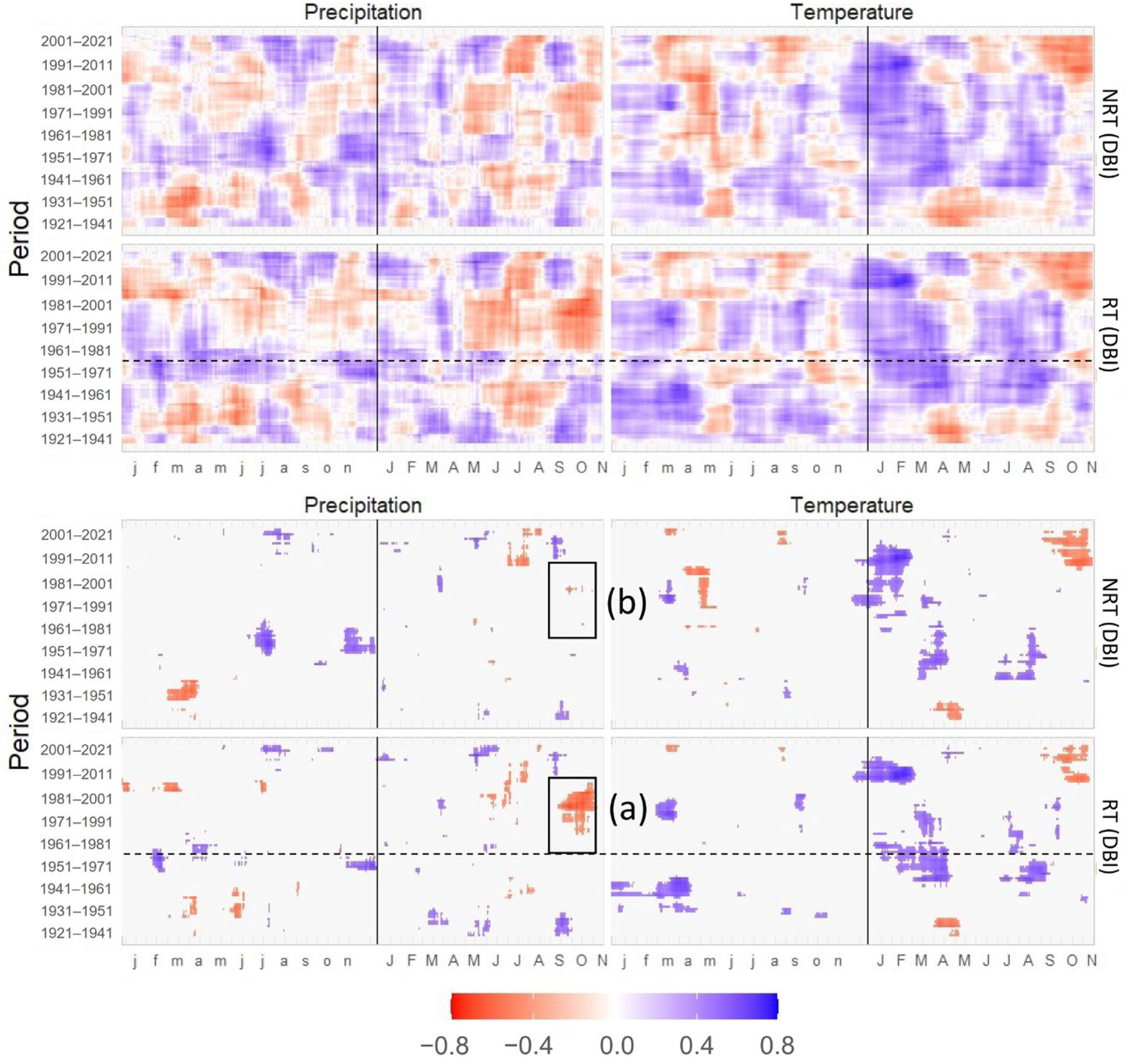
3.2. Statistical Analysis of Signals between the Periods
3.3. Correlations of TRW and DBI with Mean Temperature and Precipitation
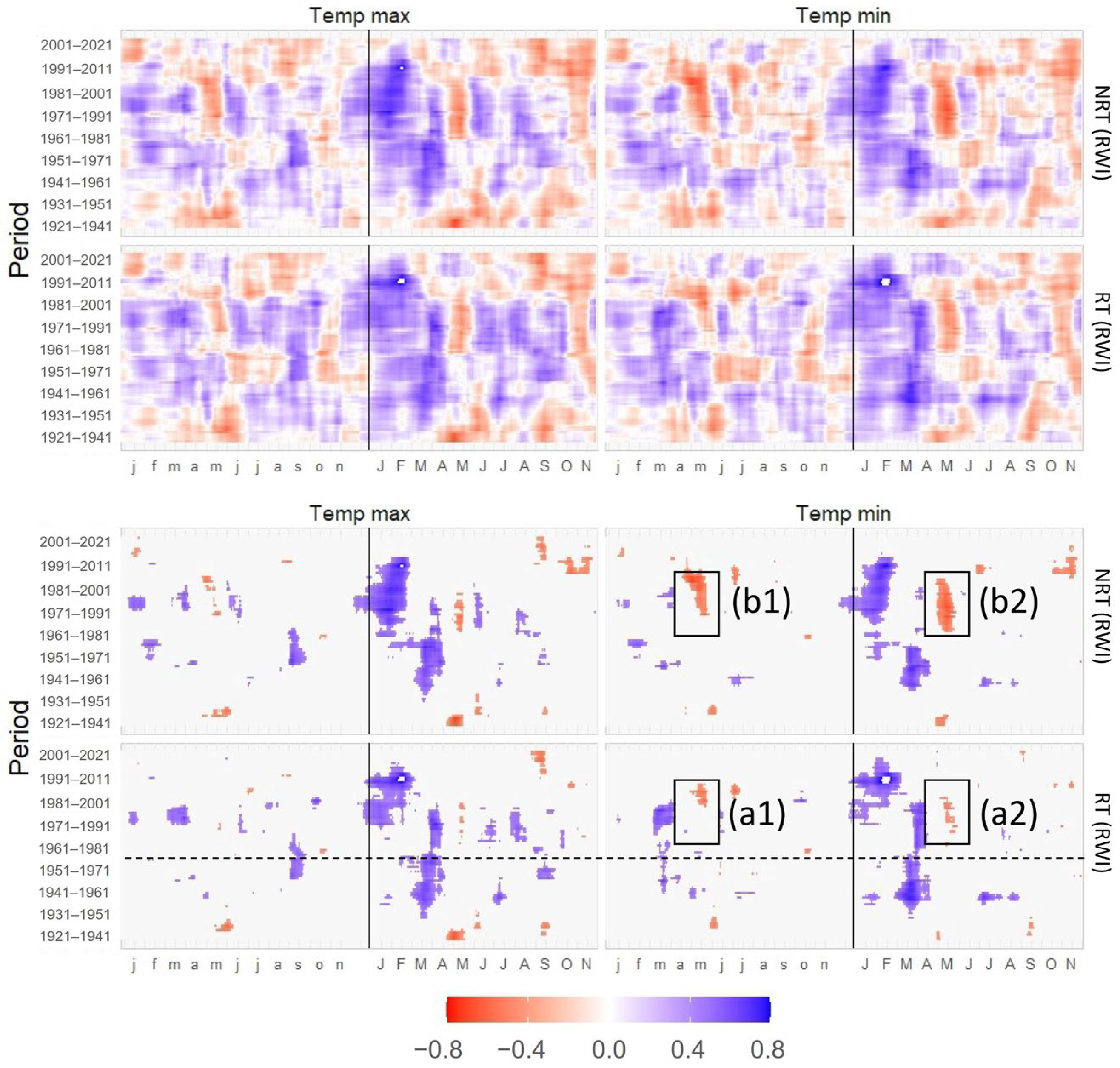
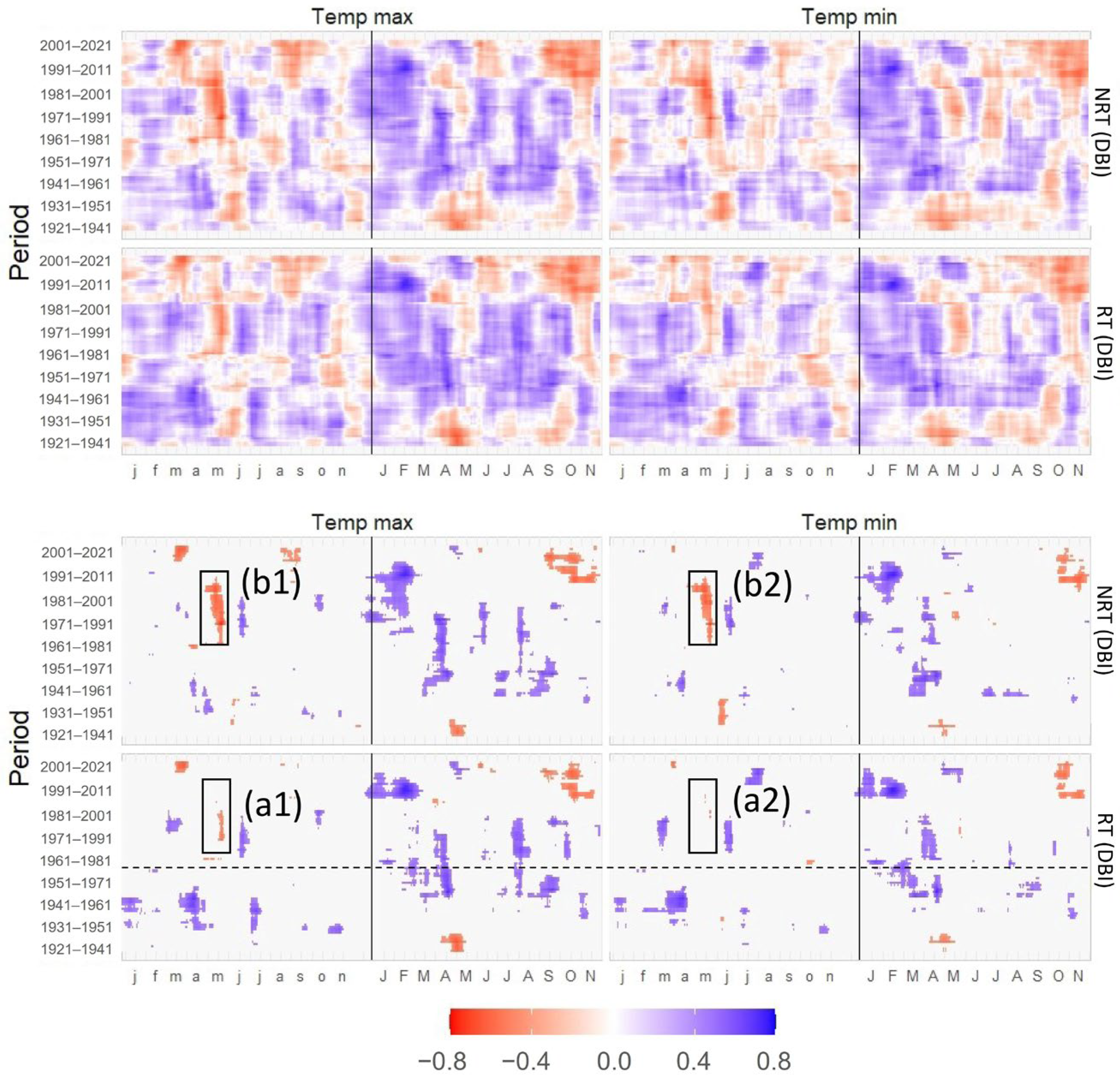
3.4. Correlations of TRW and DBI with Maximum and Minimum Temperatures
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- van der Maaten, E.; Mehl, A.; Wilmking, M.; van der Maaten-Theunissen, M. Tapping the Tree-Ring Archive for Studying Effects of Resin Extraction on the Growth and Climate Sensitivity of Scots Pine. For. Ecosyst. 2017, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Michalec, K.; Wąsik, R.; Barszcz, A. Shaping of the Width of Tree rings in Scots pine as a Result of Resin Tapping. Acta Silvestria 2020, LVI, 49–60. [Google Scholar] [CrossRef]
- Magnuszewski, M.; Tomusiak, R. Effect of Resin-Tapping on the Radial Increment of Scots pine (Pinus sylvestris L.)—Case Study of a Stand from Lidzbark Forest District. For. Res. Pap. 2013, 74, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Zaluma, A.; Strike, Z.; Rieksts-Riekstiņš, R.; Gaitnieks, T.; Vasaitis, R. Long-Term Pathological Consequences of Resin Tapping Wounds on Stems of Scots Pine (Pinus sylvestris L.). Trees 2022, 36, 1507–1514. [Google Scholar] [CrossRef]
- Garcia-Forner, N.; Campelo, F.; Carvalho, A.; Vieira, J.; Rodríguez-Pereiras, A.; Ribeiro, M.; Salgueiro, A.; Silva, M.E.; Louzada, J.L. Growth-Defence Trade-Offs in Tapped Pines on Anatomical and Resin Production. For. Ecol. Manag. 2021, 496, 119406. [Google Scholar] [CrossRef]
- Misi, D.; Puchałka, R.; Pearson, C.; Robertson, I.; Koprowski, M. Differences in the Climate-Growth Relationship of Scots Pine: A Case Study from Poland and Hungary. Forests 2019, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Czyżyk, K. Radial Growth Response of Scots Pine (Pinus sylvestris L.) after Increasing the Availability of Water Resources. Forests 2021, 12, 1053. [Google Scholar] [CrossRef]
- Debel, A.; Meier, W.J.-H.; Bräuning, A. Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years. Forests 2021, 12, 1433. [Google Scholar] [CrossRef]
- Popa, A.; Popa, I.; Roibu, C.-C.; Badea, O.N. Do Different Tree-Ring Proxies Contain Different Temperature Signals? A Case Study of Norway Spruce (Picea abies (L.) Karst) in the Eastern Carpathians. Plants 2022, 11, 2428. [Google Scholar] [CrossRef]
- Gunnarson, B.E.; Linderholm, H.W.; Moberg, A. Improving a Tree-Ring Reconstruction from West-Central Scandinavia: 900 Years of Warm-Season Temperatures. Clim. Dyn. 2011, 36, 97–108. [Google Scholar] [CrossRef]
- Rydval, M.; Larsson, L.-Å.; McGlynn, L.; Gunnarson, B.E.; Loader, N.J.; Young, G.H.F.; Wilson, R. Blue Intensity for Dendroclimatology: Should We Have the Blues? Experiments from Scotland. Dendrochronologia 2014, 32, 191–204. [Google Scholar] [CrossRef]
- Wilson, R.; Wilson, D.; Rydval, M.; Crone, A.; Büntgen, U.; Clark, S.; Ehmer, J.; Forbes, E.; Fuentes, M.; Gunnarson, B.E.; et al. Facilitating Tree-Ring Dating of Historic Conifer Timbers Using Blue Intensity. J. Archaeol. Sci. 2017, 78, 99–111. [Google Scholar] [CrossRef] [Green Version]
- López, R.; Cano, F.J.; Rodríguez-Calcerrada, J.; Sangüesa-Barreda, G.; Gazol, A.; Camarero, J.J.; Rozenberg, P.; Gil, L. Tree-Ring Density and Carbon Isotope Composition Are Early-Warning Signals of Drought-Induced Mortality in the Drought Tolerant Canary Island Pine. Agric. For. Meteorol. 2021, 310, 108634. [Google Scholar] [CrossRef]
- Pompa-García, M.; Hevia, A.; Camarero, J.J. Minimum and Maximum Wood Density as Proxies of Water Availability in Two Mexican Pine Species Coexisting in a Seasonally Dry Area. Trees 2021, 35, 597–607. [Google Scholar] [CrossRef]
- Camarero, J.J.; Hevia, A. Links between Climate, Drought and Minimum Wood Density in Conifers. IAWA J. 2020, 41, 236–255. [Google Scholar] [CrossRef]
- Björklund, J.; von Arx, G.; Nievergelt, D.; Wilson, R.; Van den Bulcke, J.; Günther, B.; Loader, N.J.; Rydval, M.; Fonti, P.; Scharnweber, T.; et al. Scientific Merits and Analytical Challenges of Tree-Ring Densitometry. Rev. Geophys. 2019, 57, 1224–1264. [Google Scholar] [CrossRef] [Green Version]
- Babst, F.; Frank, D.; Büntgen, U.; Nievergelt, D.; Esper, J. Effect of Sample Preparation and Scanning Resolution on the Blue Reflectance of Picea abies. TRACE Proc. 2009, 7, 188–195. [Google Scholar]
- McCarroll, D.; Pettigrew, E.; Luckman, A.; Guibal, F.; Edouard, J.-L. Blue Reflectance Provides a Surrogate for Latewood Density of High-Latitude Pine Tree Rings. Arct. Antarct. Alp. Res. 2002, 34, 450–453. [Google Scholar] [CrossRef]
- Campbell, R.; McCarroll, D.; Loader, N.J.; Grudd, H.; Robertson, I.; Jalkanen, R. Blue Intensity in Pinus sylvestris Tree-Rings: Developing a New Palaeoclimate Proxy. Holocene 2007, 17, 821–828. [Google Scholar] [CrossRef] [Green Version]
- Björklund, J.A.; Gunnarson, B.E.; Seftigen, K.; Esper, J.; Linderholm, H.W. Blue Intensity and Density from Northern Fennoscandian Tree Rings, Exploring the Potential to Improve Summer Temperature Reconstructions with Earlywood Information. Clim. Past 2014, 10, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Rao, R.; Rydval, M.; Wood, C.; Larsson, L.-Å.; Luckman, B.H. Blue Intensity for Dendroclimatology: The BC Blues: A Case Study from British Columbia, Canada. Holocene 2014, 24, 1428–1438. [Google Scholar] [CrossRef]
- Kaczka, R.J.; Spyt, B.; Janecka, K.; Beil, I.; Büntgen, U.; Scharnweber, T.; Nievergelt, D.; Wilmking, M. Different Maximum Latewood Density and Blue Intensity Measurements Techniques Reveal Similar Results. Dendrochronologia 2018, 49, 94–101. [Google Scholar] [CrossRef]
- Reid, E.; Wilson, R. Delta Blue Intensity vs. Maximum Density: A Case Study Using Pinus uncinata in the Pyrenees. Dendrochronologia 2020, 61, 125706. [Google Scholar] [CrossRef]
- Heeter, K.J.; Harley, G.L.; Van De Gevel, S.L.; White, P.B. Blue Intensity as a Temperature Proxy in the Eastern United States: A Pilot Study from a Southern Disjunct Population of Picea rubens (Sarg.). Dendrochronologia 2019, 55, 105–109. [Google Scholar] [CrossRef]
- Blake, S.A.P.; Palmer, J.G.; Björklund, J.; Harper, J.B.; Turney, C.S.M. Palaeoclimate Potential of New Zealand Manoao colensoi (Silver Pine) Tree Rings Using Blue-Intensity (BI). Dendrochronologia 2020, 60, 125664. [Google Scholar] [CrossRef]
- Janecka, K.; Harvey, J.E.; Trouillier, M.; Kaczka, R.J.; Metslaid, S.; Metslaid, M.; Buras, A.; Wilmking, M. Higher Winter-Spring Temperature and Winter-Spring/Summer Moisture Availability Increase Scots Pine Growth on Coastal Dune Microsites Around the South Baltic Sea. Front. For. Glob. Chang. 2020, 3, 578912. [Google Scholar] [CrossRef]
- Wilson, R.; D’Arrigo, R.; Andreu-Hayles, L.; Oelkers, R.; Wiles, G.; Anchukaitis, K.; Davi, N. Experiments Based on Blue Intensity for Reconstructing North Pacific Temperatures along the Gulf of Alaska. Clim. Past 2017, 13, 1007–1022. [Google Scholar] [CrossRef] [Green Version]
- Buckley, B.M.; Hansen, K.G.; Griffin, K.L.; Schmiege, S.; Oelkers, R.; D’Arrigo, R.D.; Stahle, D.K.; Davi, N.; Nguyen, T.Q.T.; Le, C.N.; et al. Blue Intensity from a Tropical Conifer’s Annual Rings for Climate Reconstruction: An Ecophysiological Perspective. Dendrochronologia 2018, 50, 10–22. [Google Scholar] [CrossRef]
- Fuentes, M.; Salo, R.; Björklund, J.; Seftigen, K.; Zhang, P.; Gunnarson, B.; Aravena, J.-C.; Linderholm, H.W. A 970-Year-Long Summer Temperature Reconstruction from Rogen, West-Central Sweden, Based on Blue Intensity from Tree Rings. Holocene 2018, 28, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Babst, F.; Wright, W.E.; Szejner, P.; Wells, L.; Belmecheri, S.; Monson, R.K. Blue Intensity Parameters Derived from Ponderosa Pine Tree Rings Characterize Intra-Annual Density Fluctuations and Reveal Seasonally Divergent Water Limitations. Trees 2016, 30, 1403–1415. [Google Scholar] [CrossRef]
- Seftigen, K.; Fuentes, M.; Ljungqvist, F.C.; Björklund, J. Using Blue Intensity from Drought-Sensitive Pinus sylvestris in Fennoscandia to Improve Reconstruction of Past Hydroclimate Variability. Clim. Dyn. 2020, 55, 579–594. [Google Scholar] [CrossRef]
- Akhmetzyanov, L.; Sánchez-Salguero, R.; García-González, I.; Domínguez-Delmás, M.; Sass-Klaassen, U. Blue Is the Fashion in Mediterranean Pines: New Drought Signals from Tree-Ring Density in Southern Europe. Sci. Total Environ. 2023, 856, 159291. [Google Scholar] [CrossRef]
- Mills, C.M.; Crone, A.; Wood, C.; Wilson, R. Dendrochronologically Dated Pine Buildings from Scotland: The SCOT2K Native Pine Dendrochronology Project. Vernac. Archit. 2017, 48, 23–43. [Google Scholar] [CrossRef]
- Akhmetzyanov, L.; Sánchez-Salguero, R.; García-González, I.; Buras, A.; Dominguez-Delmás, M.; Mohren, F.; den Ouden, J.; Sass-Klaassen, U. Towards a New Approach for Dendroprovenancing Pines in the Mediterranean Iberian Peninsula. Dendrochronologia 2020, 60, 125688. [Google Scholar] [CrossRef]
- Génova, M.; Caminero, L.; Dochao, J. Resin Tapping in Pinus pinaster: Effects on Growth and Response Function to Climate. Eur. J. For. Res. 2014, 133, 323–333. [Google Scholar] [CrossRef]
- Soliño, M.; Yu, T.; Alía, R.; Auñón, F.; Bravo-Oviedo, A.; Chambel, M.R.; de Miguel, J.; del Río, M.; Justes, A.; Martínez-Jauregui, M.; et al. Resin-Tapped Pine Forests in Spain: Ecological Diversity and Economic Valuation. Sci. Total Environ. 2018, 625, 1146–1155. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Sun, S.; Wang, Y.; Chang, Y.; Tao, X.; Hou, M.; Wang, W.; Liu, X.; Zhang, L. Does Resin Tapping Affect the Tree-Ring Growth and Climate Sensitivity of the Chinese Pine (Pinus tabuliformis) in the Loess Plateau, China? Dendrochronologia 2021, 65, 125800. [Google Scholar] [CrossRef]
- Hao-ran, W.U.; Yan-ru, F.a.N.; Xiao-yun, N.I.U.; Qi-fu, L.; Yan-jie, L.I.; Jing-min, J.; Jian-er, J.I.N. Effects of Resin-Tapping Year on Wood Properties of Living Pinus elliottii. For. Res. 2022, 35, 31–39. [Google Scholar] [CrossRef]
- Maxwell, R.S.; Larsson, L.-A. Measuring Tree-Ring Widths Using the CooRecorder Software Application. Dendrochronologia 2021, 67, 125841. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 March 2023).
- Jevšenak, J.; Levanič, T. DendroTools: R Package for Studying Linear and Nonlinear Responses between Tree-Rings and Daily Environmental Data. Dendrochronologia 2018, 48, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Jevšenak, J. New Features in the DendroTools R Package: Bootstrapped and Partial Correlation Coefficients for Monthly and Daily Climate Data. Dendrochronologia 2020, 63, 125753. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef] [Green Version]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Speer, J.H. Fundamentals of Tree-Ring Research; The University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Barbaroux, C.; Bréda, N. Contrasting Distribution and Seasonal Dynamics of Carbohydrate Reserves in Stem Wood of Adult Ring-Porous Sessile Oak and Diffuse-Porous Beech Trees. Tree Physiol. 2002, 22, 1201–1210. [Google Scholar] [CrossRef]
- Kilbride, J. New Dendrochronological Approaches to Identifying Climate Drivers in Northern Conifers. UVM Honors College Senior Thesis, The University of Vermont, Burlington, VT, USA, 2016. [Google Scholar]
- Cao, X.; Hu, H.; Kao, P.; Buckley, B.M.; Dong, Z.; Chen, X.; Zhou, F.; Fang, K. Improved Spring Temperature Reconstruction Using Earlywood Blue Intensity in Southeastern China. Int. J. Climatol. 2022, 42, 6204–6220. [Google Scholar] [CrossRef]
- Waszak, N.; Robertson, I.; Puchałka, R.; Przybylak, R.; Pospieszyńska, A.; Koprowski, M. Investigating the Climate-Growth Response of Scots Pine (Pinus sylvestris L.) in Northern Poland. Atmosphere 2021, 12, 1690. [Google Scholar] [CrossRef]
- Matisons, R.; Jansone, D.; Elferts, D.; Adamovičs, A.; Schneck, V.; Jansons, Ā. Plasticity of Response of Tree-Ring Width of Scots Pine Provenances to Weather Extremes in Latvia. Dendrochronologia 2019, 54, 1–10. [Google Scholar] [CrossRef]
- Kalbarczyk, R.; Ziemiańska, M.; Nieróbca, A.; Dobrzańska, J. The Impact of Climate Change and Strong Anthropopressure on the Annual Growth of Scots Pine (Pinus sylvestris L.) Wood Growing in Eastern Poland. Forests 2018, 9, 661. [Google Scholar] [CrossRef] [Green Version]
- Janecka, K.; Metslaid, S.; Metslaid, M.; Harvey, J.E.; Wilmking, M. Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea. Forests 2022, 13, 477. [Google Scholar] [CrossRef]
- Candel-Pérez, D.; Lo, Y.-H.; Blanco, J.A.; Chiu, C.-M.; Camarero, J.J.; González de Andrés, E.; Imbert, J.B.; Castillo, F.J. Drought-Induced Changes in Wood Density Are Not Prevented by Thinning in Scots Pine Stands. Forests 2018, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Björklund, J.; Gunnarson, B.E.; Seftigen, K.; Zhang, P.; Linderholm, H.W. Using Adjusted Blue Intensity Data to Attain High-Quality Summer Temperature Information: A Case Study from Central Scandinavia. Holocene 2015, 25, 547–556. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | NRT | RT | ||
|---|---|---|---|---|
| TRW | DBI | TRW | DBI | |
| No. of trees | 17 | 14 | 22 | 16 |
| DBH (cm) | 47.4 | 49.7 | ||
| Height (m) | 30.7 | |||
| GLK | 0.659 | 0.645 | 0.664 | 0.626 |
| AR1 | 0.747 | 0.628 | 0.762 | 0.691 |
| RBAR | 0.506 | 0.344 | 0.472 | 0.332 |
| EPS | 0.942 | 0.873 | 0.948 | 0.881 |
| SNR | 16.36 | 6.90 | 18.29 | 7.42 |
| Statistic | Period | NRT | RT | ||
|---|---|---|---|---|---|
| TRW (mm) | DBI | TRW (mm) | DBI | ||
| Mean | Before tapping | 0.91 | 1.04 | 0.97 | 1.06 |
| SD | 1935–1974 | 0.239 | 0.07 | 0.254 | 0.069 |
| CV | 26.1% | 7.05% | 26.1% | 6.6% | |
| Mean | Tapping period | 0.57 | 0.89 | 0.55 | 0.81 |
| SD | 1976–1980 | 0.096 | 0.075 | 0.120 | 0.129 |
| CV | 16.7% | 8.5% | 21.6% | 16% | |
| Mean | After tapping | 1.1 | 0.98 | 1.38 | 0.93 |
| SD | 1981–2021 | 0.354 | 0.099 | 0.398 | 0.118 |
| CV | 32.1% | 10.1% | 28.8% | 12.7% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubowski, M.; Dobroczyński, M. Long-Term Climate Sensitivity of Resin-Tapped and Non-Resin-Tapped Scots Pine Trees Based on Tree Ring Width and Blue Intensity. Forests 2023, 14, 593. https://doi.org/10.3390/f14030593
Jakubowski M, Dobroczyński M. Long-Term Climate Sensitivity of Resin-Tapped and Non-Resin-Tapped Scots Pine Trees Based on Tree Ring Width and Blue Intensity. Forests. 2023; 14(3):593. https://doi.org/10.3390/f14030593
Chicago/Turabian StyleJakubowski, Marcin, and Marek Dobroczyński. 2023. "Long-Term Climate Sensitivity of Resin-Tapped and Non-Resin-Tapped Scots Pine Trees Based on Tree Ring Width and Blue Intensity" Forests 14, no. 3: 593. https://doi.org/10.3390/f14030593