
Improvement Cuttings in the Conversion of Pyrenean Oak (Quercus pyrenaica Willd.) Coppice
Department of Forest Sciences and Landscape Architecture, University of Tras-os-Montes and Alto Douro, Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), P.O. Box 1013, 5000-501 Vila Real, Portugal
Forests 2023, 14(3), 575; https://doi.org/10.3390/f14030575
Submission received: 14 February 2023
/
Revised: 10 March 2023
/
Accepted: 11 March 2023
/
Published: 14 March 2023
(This article belongs to the Special Issue Innovative Management and Multifunctional Utilization of Traditional Coppice Forests)
Abstract
:An appropriate management and valuation of Pyrenean oak (Quercus pyrenaica Willd.) forests are essential in order to ensure their multiple functions and uses. Many coppice forests of Pyrenean oak have been converted into high forests mostly due to changing demands. This work presents a 20-year study conducted with Pyrenean oak coppices, providing results from an experimental trial where silvicultural improvement interventions for coppice conversion were applied. In addition to control, selective thinning and 3-strip thinning with 2 × 2 m, 3 × 3 m, and 3 × 6 m (the latter also included selective thinning) were examined. The study analyzed tree growth, vertical stratification dynamics, and epicormic branching. It was found that the silvicultural interventions promoted tree development differently according to social crown class. The results have shown a significantly different response in tree diameter growth and live crown length, while height growth has been less accentuated. The study also examined the temporal effect of the interventions on diameter growth. Changes in tree crown position and structural characteristics varied among the cutting interventions. Regarding epicormic branching, the results have revealed that the number of shoots does not differ significantly among the treatments but shows differences in length. Management issues regarding Pyrenean oak coppice conversion have also been addressed.
1. Introduction
Pyrenean oak (Quercus pyrenaica Willd.) is a sub-Mediterranean species that occurs mostly in the Iberian Peninsula and, to a lesser extent, in the southwest of France and North Africa, with a total surface of 1.19 million hectares. In Portugal, Pyrenean oak covers a forest area of 62,000 ha [1] (NFI 2015). Pyrenean oak forests are a natural resource of great value providing important ecological, social, and economic functions.
Considering its bio-ecological characteristics, Pyrenean oak plays an important role in the conservation of biodiversity (e.g., [2]). Pyrenean oak woodlands are also relevant in soil and water conservation as well as in promoting water quality. In addition, they are important in the preservation of the natural landscape, providing an excellent environment for recreation and leisure. Another relevant function relies on the prevention of forest fires because of its lower combustibility compared with other fire-prone forest species (e.g., [3,4]).
Pyrenean oak wood may have multiple uses, such as carpentry, construction, furniture, veneer, flooring, cooperage, charcoal production, and firewood. Pyrenean oak still represents a fundamental wood species and valuable resource for people living in rural areas. These oak woodlands also offer a variety of non-wood goods (e.g., wild mushrooms, herbs, fruits, and game), altogether contributing to economic and social functions ([5,6] Carvalho, 2012; Santos et al., 2012).
Some Pyrenean oak forests have been exploited as coppices for the production of small-size material, mostly for firewood [5,7]. The species is considered to have a good ability to regenerate either by stool or root sprouts [8]. Usual rotations vary between 15 to 30 years, depending on the site quality. Human practices have significantly modified the distribution, structure, and composition of these forests.
Pyrenean oak forests have a long history of coppicing; however, several previously managed coppices are currently under conversion into high forests. In the past, the coppice had a greater use, but during the last decades, several coppices have been abandoned and converted. Some coppices are still maintained exclusively for firewood production, mostly on poor sites. Pyrenean oak coppices are characterized by having small-size trees and, compared with the high forest, present a lesser interest in biodiversity conservation and recreation. They represent a lower stand development stage as a result of repeated clear-cuts or other disturbances (e.g., wildfires). In some particular cases, specific biologic conservation is required in applying canopy gaps or landscape mosaics where coppice may take place (e.g., [9,10,11]).
Coppice conversion might be a management option depending on ecological and social context and goals [12]. Economic and social changes in the last decades have supported the conversion of many coppices into high forests, a common situation in other parts of European countries, while in some areas coppices have also been abandoned ([5,13,14,15,16,17,18]).
The management of Pyrenean oak is nowadays favorably oriented in pursuing multiple functions [5,19,20]. The conversion of many coppice forests of this species has been a management option to improve and valuate the oaks for their multiple functions, including the enhancement of biodiversity, protection, and aesthetics, as well as timber production for more valuable uses. This improves oak forests’ maintenance and active management with positive consequences for the environment (e.g., [21,22,23]). Through proper management, it is possible to get goods and services in a sustainable way that considers not only the socioeconomic needs but also conservation issues and ecosystem functioning. An advanced stand development stage also presents higher resistance to fire [5]. There are also additional benefits from the improvement and valuation of the wood to higher end-use values (e.g., [21,24,25,26]) and carbon storage (e.g., [27,28,29]). New industrial technologies also allow the use and valuation of diverse wood applications [30,31]. Silviculture should, therefore, seek a valuation of its products, allowing greater profitability and other ecological and social functions [12,32,33,34,35]. The high forest provides a higher yield per unit area and higher value compared with a coppice (e.g., [36] Hubert, 1999). From a silvicultural point of view, the production of firewood can be compatible with timber production in oak high forests. Through proper planning and management, high forests may supply firewood as well as round timber. Small-size woody material resulting from thinning and harvesting may meet firewood needs.
The conversion is an intermediate silvicultural intervention for the change of a coppice into a high forest of the same species [23,37,38]. This involves the change of a silvicultural system by a new stand originated from sexual reproduction [39,40]. A common way of conversion is by aging the coppice and obtaining well-developed trees to initiate the natural seed regeneration process. It depends on the site quality, initial density, and intervention intensity (e.g., [13,41]). Normally, this conversion involves an intermediate stage where selective cuttings are applied favoring the best promising trees, with the first thinning applied close to the end of the coppice rotation [5]. Techniques dealing with the conversion of forest stands have been tested in different forest ecosystems and socio-ecological environments [14,42,43,44], and a few studies cover the methods tested here for coppice conversion [45]. Conversion is a long process and a management option is required to define appropriate conversion perspectives.
In 1995 an experimental trial was installed to study the conversion of Pyrenean oak coppices. This study was conducted for 20 years aiming to analyze tree response to different silvicultural conversion practices. More specifically, the study aimed to analyze the effects on the growth of tree parameters, stand structural dynamics, and tree epicormic branching. This is the first study on the species regarding this particular silvicultural intervention of conversion. This study was initially part of the EU-AIR Research Project ‘Improvement of Coppice Forests in the Mediterranean Region’ (1994-98).
2. Material and Methods
2.1. Study Site
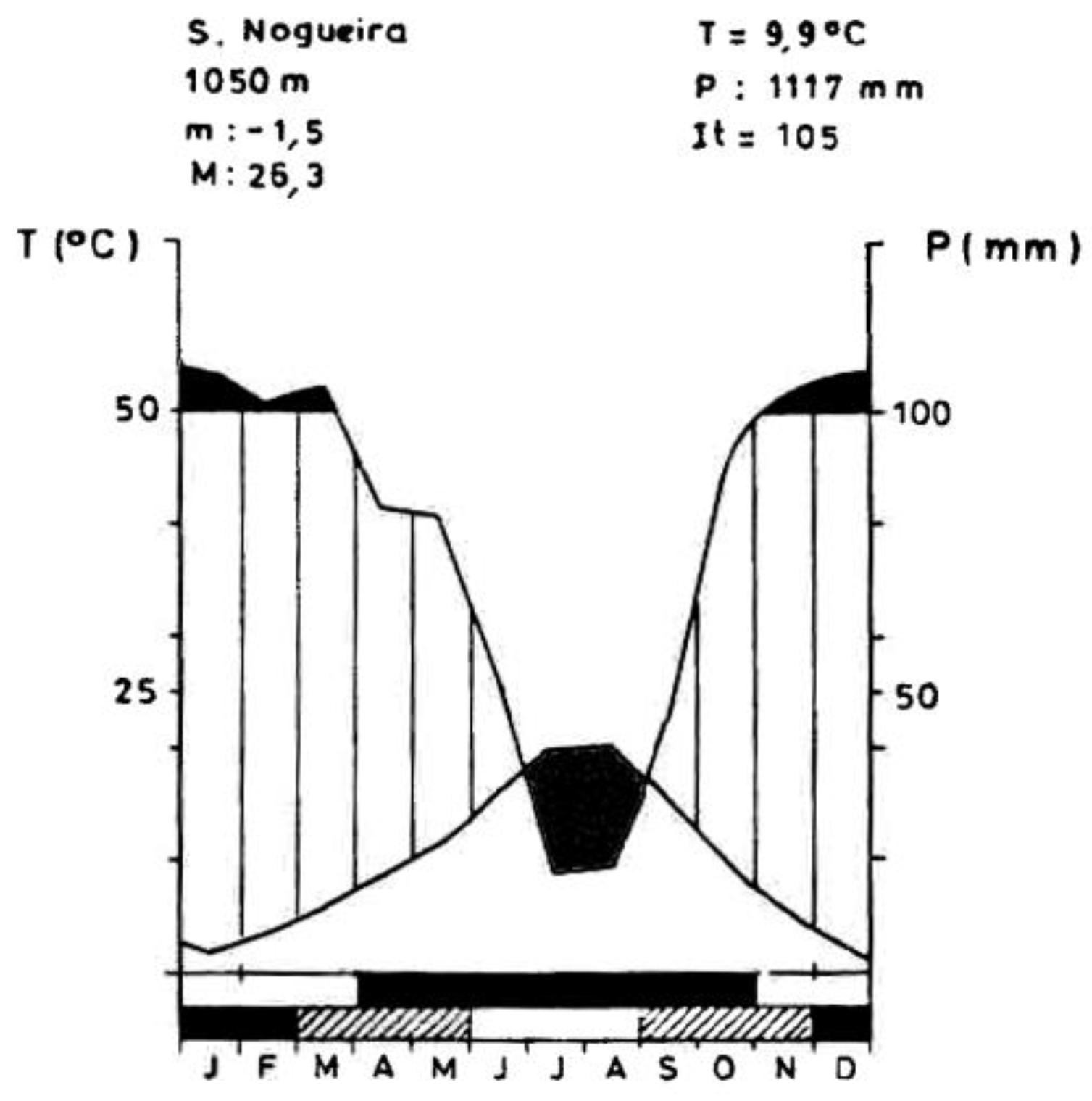
The experimental area is located in the Nogueira mountain, NE Portugal (41°44′ N, 6°51′ W). The site is situated at an elevation of 1140 m asl, SE exposure, and a slope ranging between 7 and 12°. The climate is Mediterranean with an annual precipitation of 1117 mm and a mean temperature of 9.9 °C, belonging to the supra-Mediterranean bioclimatic zone and humid ombroclimate (Figure 1). Geology is based on blastomilonites from the Precambrian polymetamorphic series [46,47]. The soil is a Cambissol with a loam texture, an average depth of 70 cm, and soil organic matter of 3.5%. The soil is slightly acidic with a pH (H2O) of 6.0. The area is dominated by oak woodlands belonging to the climatophilous vegetation of Holco mollis—Quercetum pyrenaicae Br-Bl P Silva & Rozeira, 1956. The study site is located in the Nogueira Mountain Forest Area under the management of the Forest Service (ICNF—Institute for Nature Conservation and Forests).
2.2. Experimental Design and Data Analysis
The experimental trial was installed in 1995 on a Pyrenean oak coppice, aged 28 years, with a mean stand height of 6 m. Table 1 summarizes the general biometric data of the coppice. Estimates on the stand biomass (W) and site quality (SI50) were based on studies performed by [48,49]. No prior thinning was performed in the coppice area. Typically, these coppices present high stand density, and conversion starts as a first thinning close to the end of coppice rotation.
The trial consisted of 5 treatments where improvement cuttings were applied. It included control (C), selective thinning (S), and three modalities of strip thinning. The strip thinning involved alternate rows of 2 × 2 m, 3 × 3 m, and 3 × 6 m. In the last one, an additional selective thinning was applied in the standing strip. The strip thinning means that is made up of successive alternate strips of clear-cuts and coppice. Each plot had a surface area of 800 m2 and 2 replications were installed for each treatment. The general approach was to apply to all treatments a similar cutting intensity of about 50% relative stocking reduction (number of trees). On the control plots, no trees were felled.
On each plot, 45 sample trees were chosen and marked, equally distributed over three crown classes (dominant, intermediate, and suppressed). These trees were measured at the installation age (1995) and periodically afterward, in 2000, 2007, and 2015. Additionally, sample plots were installed on each treatment to measure and follow the stand number of trees, diameter, and height. Measurements were taken for DBH, total tree height (h), the base of live crown (hlc), and 4 crossed crown radii. Tree live crown length (lcl) was computed from h and hlc, and crown diameter (dc) from crown radius. DBH was measured with a diameter tape and heights with a telescopic measuring pole (Figure 2). The number and length of epicormic branches were measured up to 3 m of the stem height. Tree crown class position was also evaluated. A vertical structure index (VI) was computed based on the Shannon–Weaver index [50,51] which evaluates the relative proportion of each tree layer. A mortality rate (Mr) was calculated as follows, according to [52], with N representing the number of live individuals at time intervals t1 and t2,
The purpose was to analyze the effect of the different applied cutting practices on the growth of tree parameters, structural dynamics, and epicormic branching. Another aim of the study was related to the time effect on tree growth of the applied cuttings.
3. Results
3.1. Tree Growth
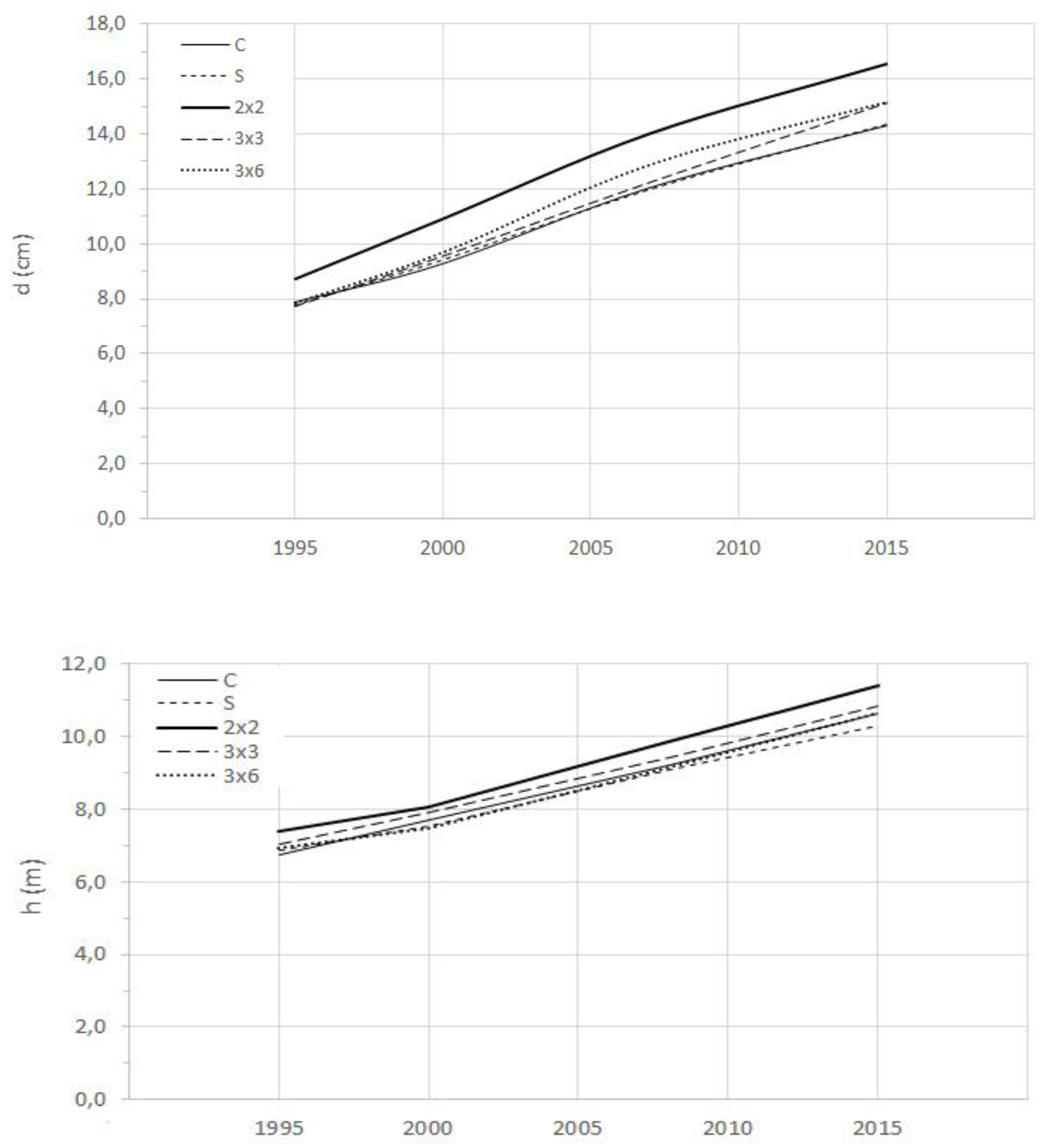
The analysis was performed to evaluate the effect of different treatments on the tree growth regarding diameter, height, crown diameter, and live crown length. Table 2 presents the growth of the trees for different treatments, and the relative increment compared with the control plot over the follow-up period (1995–2015).
There was a differentiated response among the thinning interventions for the increment of tree biometric variables. The intervention that most promoted diameter growth was 2 × 2 m, although not significantly different from the 3 × 3 m and 3 × 6 m. Regarding the tree height, the differences between treatments were of smaller magnitude compared with the diameter (Table 2, Figure 3). The S cutting presents the lowest increment in height. Crown growth was smaller in the control plot, both in terms of dc and lcl, as expected because of higher relative density. Lcl has decreased in the control and S intervention plots. Because the S thinning provides more adjacent tree-growing space, dc growth was higher. This was followed by 3 × 6 intervention, although not significantly different. This last intervention in addition to growing space from thinned strips also included inner selective thinning.
Concerning the tree diameter increment, a subsequent analysis was performed to evaluate the growth among tree crown classes (dominant, intermediate, and suppressed) (Table 3). Overall, the thinning interventions enhanced tree growth, particularly that of the dominant trees. There is a differentiated response among the tree crown classes. The dominant trees presented higher increments compared with the other classes with significantly different overall treatments (F ranging from 12.470 to 19.463, prob < 0.001). For dominant trees, the strip interventions were the ones that promoted tree growth. All cutting interventions benefited the suppressed trees, which responded to the reduction in density.
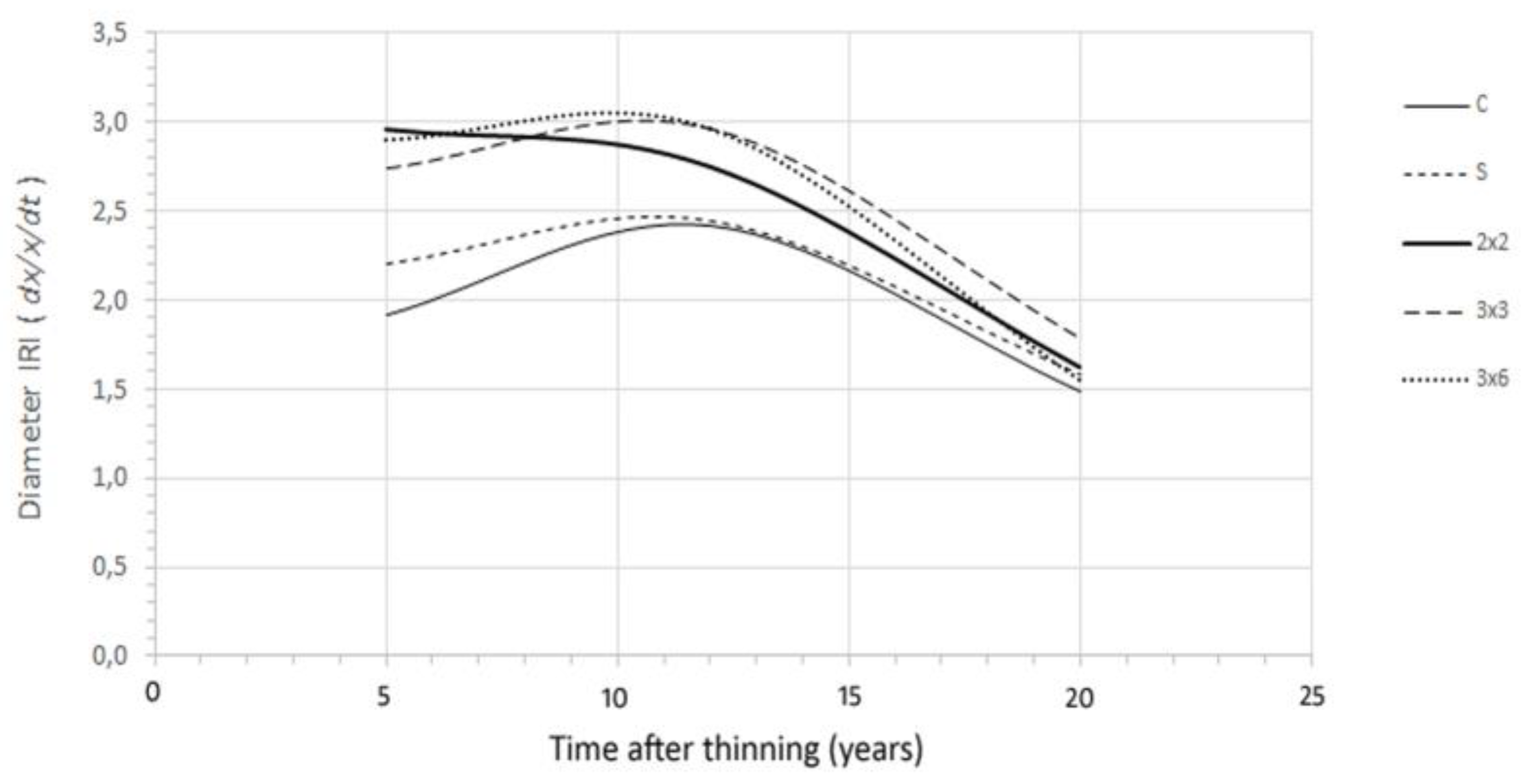
In order to evaluate the time response of thinning on tree growth, an increment rate index (IRI) for diameter was computed as ꝺx/x/ꝺt, a derivate in relation to the initial tree diameter size over time (Figure 4). It can be seen that after the intervention, there was a quick response in tree growth, which extended, in general, for 12 years. The response has been shown to vary among treatments, with 3 × 3 and 3 × 6 being the ones with a longer extended period of growth.
3.2. Structural Dynamics
Table 4 shows the dynamic of the crown classes for the sample trees in the same period. The records of the social crown position allowed for the analysis of the proportion of trees that ascend, maintain, or descend their crown structural position. Tree mortality also varied among the plots, and as expected, was higher in the control and 3 × 3 plots. Because the 3 × 6 also involved selective cutting in the standing strip, it provided more tree-growing space compared with the 2 × 2 and 3 × 3 plots (Table 4).
In all cutting interventions, a greater ascent in the tree crown class has occurred, which is also related to higher growth in tree height. The 3 × 6 presented the lower descent in the tree crown class, while S had the highest descent, which is also related to the combination of live crown length, height, and diameter increment. In general, the descent averaged 8.6% and occurred in both codominant and intermediate crown classes. The ascent occurred on average with 30.8% of trees, mostly from the intermediate class, while the majority of 60.6% maintained their social crown position. The cuttings made on the S and 3 × 6 plots imply a certain reduction in the vertical layering, which is reflected in the vertical structure index. The mortality rate was lower in the S and 3 × 6 treatments.
3.3. Epicormic Branching
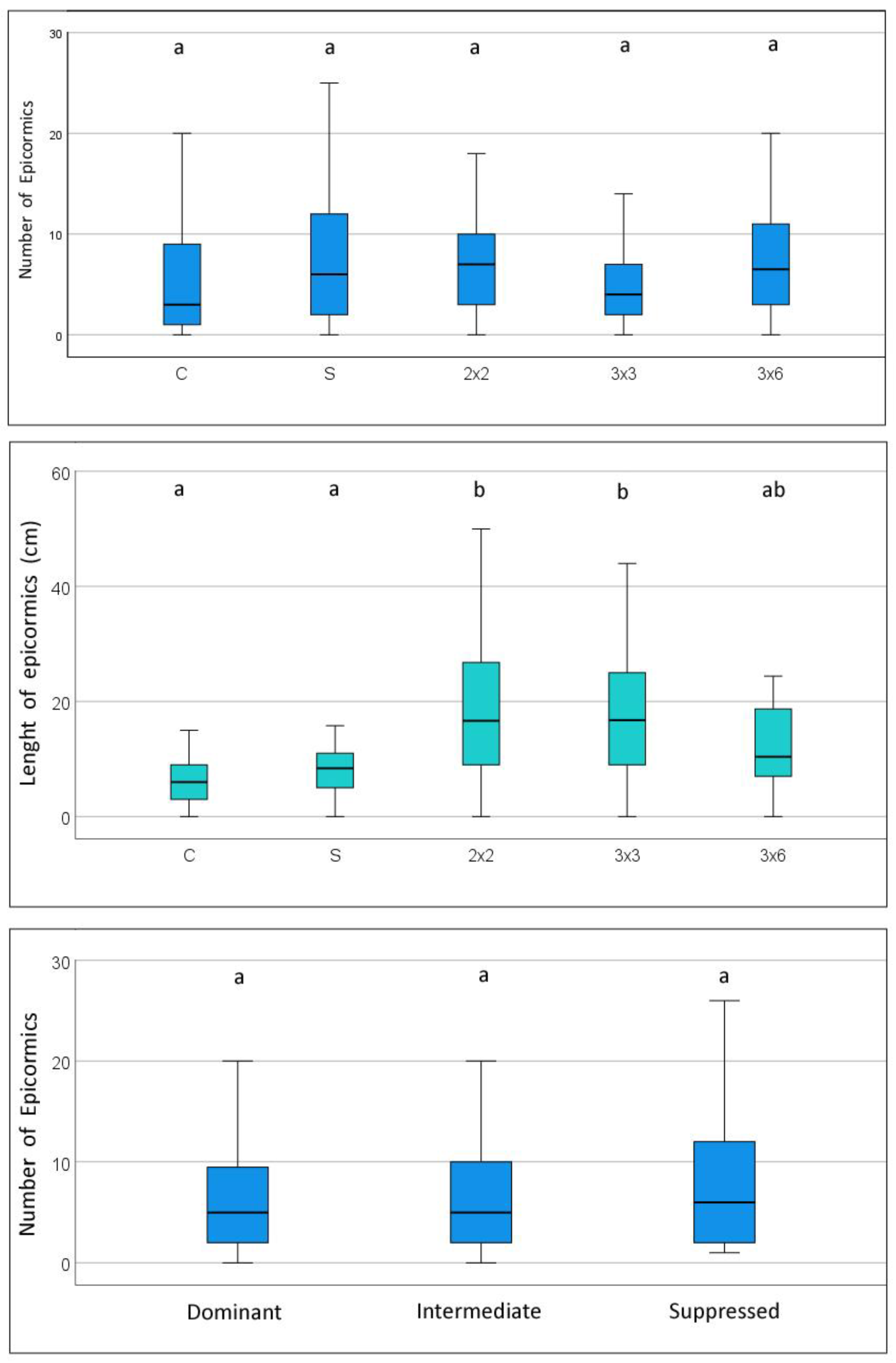
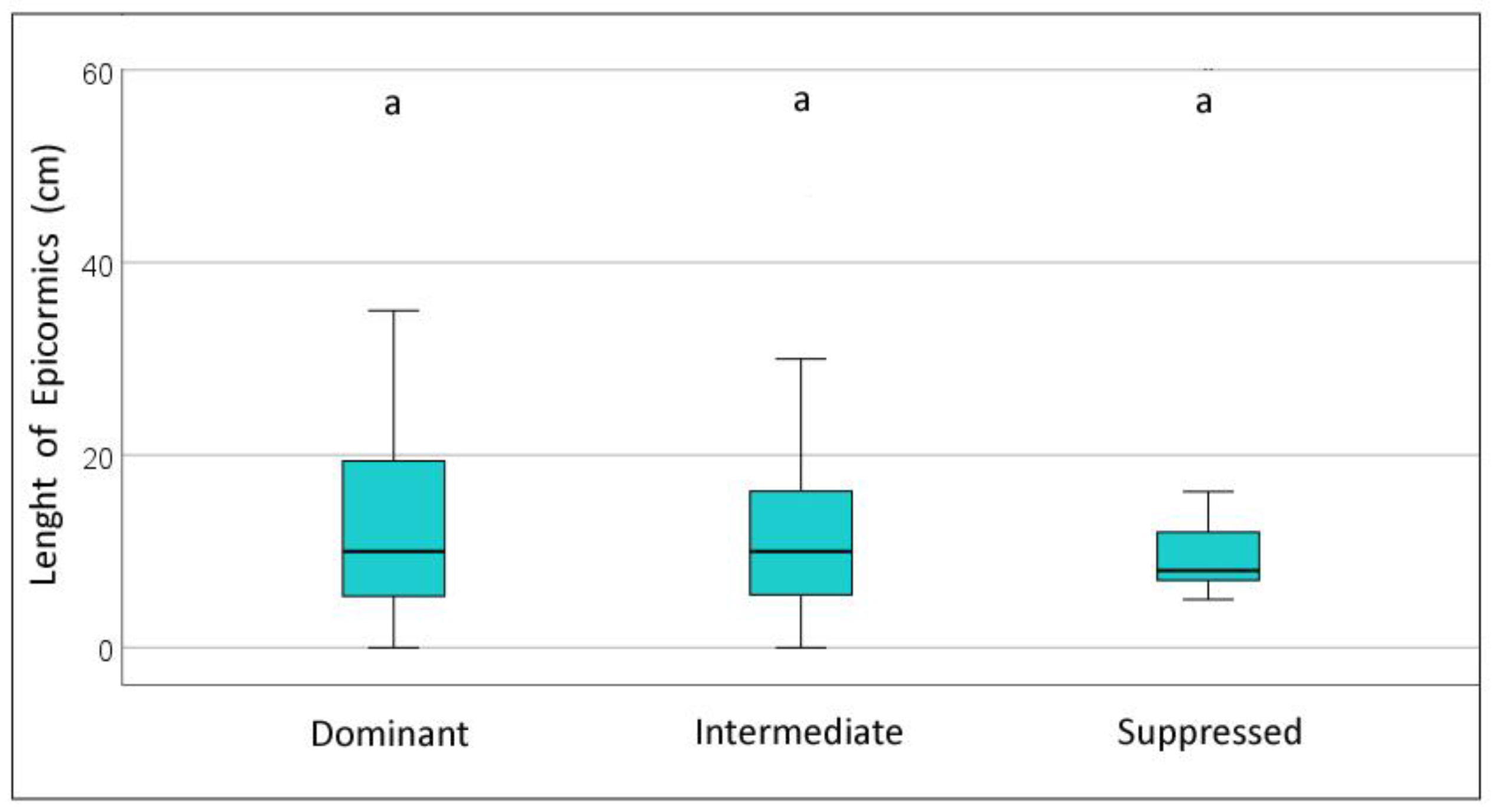
The presence and development of epicormic branches might be related to stem grading for certain specific wood end uses. Figure 5 presents the average differences concerning the number and length of epicormic branches. The results reveal that the number of epicormic branches is not significantly different among the treatments, averaging between 5.2 tree−1 (C) to 8.4 tree−1 (S). The number of epicormic branches varies widely within each plot, from tree to tree, with a large standard deviation within each treatment (coefficient of variation ranging from 80 to 110%), and several trees do not present any epicormic branches. Regarding the length of these branches, the control shows the lowest values (7.2 cm on average) whereas the strip treatments present the longer growth of the epicormics, although 3 × 6 (11.9 cm) was shorter than 2 × 2 and 3 × 3 and not significantly different from the control and selective cutting (10.4 cm). It was also observed that most of the smaller epicormics, up to 15 cm in length, usually tend to die in the short term. On average, both the number and the length of epicormics are not significantly different among the crown classes (Figure 5). The number of epicormics is slightly lower in dominant trees (6.3 tree−1) compared with intermediate (7.2 tree−1) and suppressed (8.9 tree−1), and the average length varies between 12.4 and 13.8 cm and is not significantly different.
4. Discussion
The study allowed analyzing the effects of different silvicultural interventions on tree development patterns over a long-term monitoring period, providing a better understanding of the applied conversion practices. In general, the results indicated that the Pyrenean oak coppice showed a positive response to thinning as an active management base outlined by different responses and comparable with other studies on oaks and Fagaceae (e.g., [17,44,55,56,57,58]). Thinning increased the growth performance, most likely due to the availability of light and the consequent recovery of available space [59,60,61].
The results of the study suggested that the silvicultural interventions with strip cutting presented better results. This might be explained by the relatively larger and closer available growing space provided by the cuttings, particularly by the 2 × 2 method. Among the strip-cutting methods, the 2 × 2 presented the preferable effect in terms of diameter growth, although not significantly different from the 3 × 3 and 3 × 6 methods. Comparatively, the selective thinning provided a lower diameter growth response. The differences in height growth were not substantial, although significantly lower in selective thinning. The better diameter response in the stripping methods was most likely related to a higher increment in the crown development, associated with the crown diameter and particularly the crown length, which increased in all applied cutting methods while it decreased in the untreated (control) and selective cutting.
Concerning the number of epicormic branches, no significant differences were found among the treatments, although the control presented the lowest number. The number of branches varied widely within each treatment and from tree to tree. In terms of length, it was larger in the 2 × 2 and 3 × 3 methods, while the 3 × 6, selective thinning and control presented shorter epicormics. The particular conditions that affect the emission of epicormic branches in oaks are not entirely explained and some authors (e.g., [62,63]) have found that most of the unaffected oak trees belong to the dominant crown class and tend to decrease with age, and they also have a genetic influence and variability [25,31,48]. The growth of epicormic shoots can be controlled by silvicultural interventions. With Pyrenean-oak, it tends to decrease with age and in the lower part of the stem. Its presence in Pyrenean oak is mostly superficial and has not limited the use of wood [5,6].
In general, considering both the growth response, structural dynamics, and the effect on epicormic branching, the 3 × 6 cutting method might be an advisable compromise. The increment of the tree diameter and crown diameter is higher than in the control and selective cutting, and not significantly different from the 2 × 2 and 3 × 3 cuttings, presenting lower tree mortality, as well as a smaller number and shorter epicormics than the 2 × 2 and 3 × 3 cuttings. The selective cutting presented a shorter epicormic length, although not significantly different from the 3 × 6 model. Furthermore, 3 × 6 required proportionally fewer strip clear-cuts compared with the 2 × 2 and 3 × 3 plots. As a first thinning in the conversion process, it presents the advantage of combining a selective cutting in the standing strip; at the same time, the strips also facilitate harvesting and extraction of the material, being more cost-effective [11]. This improvement combines making use of existing good-quality, vigorous trees with applying selective harvesting. This enables the production of high-gradetimber through the process of conversion. The conversion might be applied when certain conditions are ensured, namely, a potential number of trees well-adapted to the site, growing stock able to produce good-grade timber, and the possibility of future natural regeneration of the species. This allows for management flexibility and avoiding a clear-cut, thus maintaining the forest environment, biodiversity, and soil protection. It also allows for a valuation of the stand in terms of timber, non-wood products, and ecosystem services [20]. Periodic revenues are obtained and installation costs are avoided by using further natural regeneration.
The study findings also allowed us to depict the time effect of the cutting treatments for this combination of species, site quality, and thinning, helping to define a relevant silvicultural issue related to the frequency or time interval of the interventions [40]. The diameter growth rate index indicates that, in general, it starts to decrease 12 years after thinning, while the 3 × 3 and 3 × 6 models have presented an extended period of growth response.
5. Conclusions
The study analyzes the long-term response of a set of variables to silvicultural interventions on the development of a young stand of Pyrenean oak coppice. The results have confirmed a positive and differential tree response to the applied interventions that last for a certain time as has been verified by the increment rate analysis.
Pyrenean oak–dominated coppices may be managed with first conversion thinning in pursuing further development stages and steady interventions. The study presented the usefulness of active conversion practices for the management of Pyrenean oak coppices. The silvicultural practices endorse growth and dynamics allowing progression toward mature stand conditions and promoting management profitability from multiple uses and functions.
Funding
This research work was supported by FCT—Foundation for Science and Technology, under the project UIDB/04033/2020 (CITAB), the Project FCT-Scapefire PCIF/MOS/0046 ‘A sustainable landscape planning model for rural fires prevention’, and the COST Action FP1301 ‘Innovative management and multifunctional utilization of traditional coppice forests’. The trial installation and research were initiated under the Project MEDCOP AIR–EU2 CT94–0905 ‘Improvement of coppice forests in the Mediterranean region’ (1994-98).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Acknowledgments
The author also wants to thank ICNF (Institute for Nature Conservation and Forests), the Forest Service Administration of Bragança, and the municipalities of Rebordaos and Carrazeda for their support as well as to Jill Barbour for text revision and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- NFI National Forest Inventory; ICNF: Lisbon, Portugal, 2015; p. 284.
- Carvalho, J.; Alves, A.; Grosso-Silva, J.; Santos, T. Biology and ecology of Pyrenean-oak forests. In Arvores e Florestas de Portugal Vol 2—Os Carvalhais; Silva, J., Ed.; LPN, FLAD: Lisbon, Portugal, 2007; pp. 99–120. (In Portuguese) [Google Scholar]
- Salas, F. Donde ardera el bosque? Previsión de incendios forestales mediante un SIG. In Los Sistemas de Información Geográfica en la Gestión Territorial; AESIGYT: Madrid, Spain, 1992; pp. 430–446. [Google Scholar]
- González, J.; Palahi, M.; Trasobares, A.; Pukalla, T. A fire probability model for forest stands in Catalonia (north-east Spain). Ann. For. Sci. 2006, 63, 169–176. [Google Scholar] [CrossRef]
- Carvalho, J. Oak Forest Management. In Oaks: Ecology, Types and Management; Chuteira, A., Bispo, A., Eds.; Science Publishers: New York, NY, USA, 2012; pp. 1–58. [Google Scholar]
- Santos, J.; Carvalho, J.; Santos, J. Oak wood. In Oaks: Ecology, Types and Management; Chuteira, A., Bispo, A., Eds.; Science Publishers: New York, NY, USA, 2012; pp. 119–150. [Google Scholar]
- Montoya, J. Selvicultura, Ordenacion y Economia de los Rebollares de Quercus pyrenaica Willd. INIC 1982, 11, 3–13. [Google Scholar]
- Carvalho, J.; Loureiro, A. Stool and root resprouting according to different cutting seasons in a Quercus pyrenaica Willd. Coppice. Ann. Ist. Sper. Selvic. Arezzo 1996, 27, 83–88. [Google Scholar]
- Ozanne, C.; Hambler, C.; Foggo, A. The significance of edge effects in the management of forests for invertebrate biodiversity. In Canopy Arthropods; Stork, N.E., Adis, J., Didham, R.K., Eds.; Chapman & Hall: London, UK, 1997; pp. 534–550. [Google Scholar]
- Ausden, M. Habitat Management for Conservation; Oxford University Press: New York, NY, USA, 2007; p. 411. [Google Scholar]
- Sebek, P.; Bartos, M.; Benes, J.; Chlumska, Z.; Dolezal, J.; Kovar, J.; Mikatova, B.; Platek, M.; Polakova, S.; Skorpik, M.; et al. Effect of active conservation management on biodiversity: Multi-taxa survey in oak woodlands of Podyji National Park, Czech Republic. In Proceedings of the 5th Symposium for Research in Protected Areas, Mittersill, Austria, 10–12 June 2013; pp. 717–720. [Google Scholar]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of European forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Padula, M.; D’Amico, C.; Ricci, S. Gioffredi R Esperienze di conversione all’alto fusto di boschi cedui invecchiati di faggio nell’Appennino settentrionale (Italia). Ann. Acc. It. Sc. For. 1988, 37, 23–64. [Google Scholar]
- Amorini, E.; Bruschini, S.; Cutini, A.; Di Lorenzo, M.; Fabbio, G. Treatment of Turkey oak (Quercus cerris L.) coppices. Structure, biomass and silvicultural options. Ann. Ist. Sper. Selvicultura Arezzo 1996, 27, 105–111. [Google Scholar]
- Gubka, K. Development dynamics of young stands of sessile oak after clear cutting during the conversion of coppice forests in south Slovakia. Sumar. List 2000, 124, 495–502. [Google Scholar]
- Nocentini, S. Structure and management of beech (Fagus sylvatica L.) forests in Italy. Iforest—Biogeosciences For. 2009, 2, 105–113. [Google Scholar] [CrossRef]
- Ciancio, O.; Corona, P.; Lamonaca, A.; Portoghesi, L.; Travaglini, D. Conversion of clearcut beech coppices into high forests with continuous cover: A case study in central Italy. For. Ecol. Manag. 2006, 224, 235–240. [Google Scholar] [CrossRef]
- Mullerova, J.; Hedl, R.; Szabo, P. Coppice abandonment and its implications for species diversity in forest vegetation. For. Ecol. Manag. 2015, 343, 88–100. [Google Scholar] [CrossRef]
- von Teuffel, K. Consequences of increased tree growth on forest management planning and silviculture. In Causes and Consequences of Accelerating Tree Growth in Europe, Proceedings of the International Seminar held in Nancy, France, 14–16 May 1998; Karjalainen, T., Spiecker, H., Laroussinie, O., Eds.; European Forest Institute: Joensuu, Finland, 1999; pp. 229–236. [Google Scholar]
- Spiecker, H.; Hein, S.; Makkonen-Spiecker, K.; Thies, M. Valuable Broadleaved Forests in Europe; EFI Research Report 22; European Forest Institute, Brill: Leiden, The Netherlands, 2009; p. 276. [Google Scholar]
- Bourgenot, L. Conversion and Transformation Methods—General conclusion. Rev. For. Fr. 1969, 3, 199–204. [Google Scholar]
- Wolfslehner, B.; Krajter, S.; Jovic, D.; Nestorovski, L.; Velichkov, I. Framing stakeholder and policy issues for coppice forestry in selected central and south-eastern European Countries. Silva Balc. 2009, 10, 21–34. [Google Scholar]
- Nicolescu, V.; Carvalho, J.; Hochbichler, E.; Bruckman, V.; Piqué-Nicolau, M.; Hernea, C.; Viana, H.; Stochlová, P.; Ertekin, M.; Tijardovic, M.; et al. Silvicultural Guidelines for European Coppice Forests. In Coppice Forests in Europe; Unrau, A., Ed.; Albert Ludwig University Freiburg: Freiburg, Germany, 2018; pp. 46–63. [Google Scholar]
- Hubert, M. Cultiver les Arbres Feuillus Pour Récolter du Bois de Qualité; IDF: Paris, France, 1981; p. 278. [Google Scholar]
- Groome, H.; Perez, M.; Llorca, A. Quelques arguments économiques pour la valorisation et la conservation des forêts autochtones en Espagne. Forêt Méditerranéenne 1985, 7, 169–172. [Google Scholar]
- Thibaut, B. La valorisation des bois de petit diamètre. Forêt Enterp. 1993, 92, 32–37. [Google Scholar]
- Adger, W.N.; Huq, S.; Brown, K.; Conway, D.; Hulme, M. Adaptation to climate change in the developing world. Prog. Dev. Stud. 2003, 3, 179–195. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; Cannell, M.G.R. Managing forests for wood yield and carbon storage: A theoretical study. Tree Physiol. 2000, 20, 477–484. [Google Scholar] [CrossRef]
- Assmuth, A.; Rämö, J.; Tahvonen, O. Optimal Carbon Storage in Mixed-Species Size-Structured Forests. Env. Resour. Econ. 2021, 79, 249–275. [Google Scholar] [CrossRef]
- Sevrin, E. Différent modes de silviculture pour les chênes rouvre et pédonculé. Fôret Enterp. 1996, 111, 18–20. [Google Scholar]
- Triboulot, P.; Leban, J. Le bois—Nouvelles technologies et nouveaux matériaux. Forêt Enterp. 1998, 119, 17–22. [Google Scholar]
- Harmon, M.; Franklin, J.; Swanson, F.; Sollins, P.; Gregory, S.; Lattin, J.; Anderson, N.; Cline, S.; Aumen, N.; Sedell, J.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 59–234. [Google Scholar]
- Healy, W.; Brooks, R.; DeGraaf, R. Cavity trees in sawtimber size oak stands in central Massachusetts. North. J. Appl. For. 1989, 6, 61–65. [Google Scholar] [CrossRef]
- Barraclough, S. Toward Integrated and Sustainable Development? United Nations Research Institute for Social Development: Geneva, Switzerland, 2001; p. 45. [Google Scholar]
- Burger, J. Management effects on growth, production and sustainability of managed forest ecosystems: Past trends and future directions. For. Ecol. Manag. 2009, 258, 2335–2346. [Google Scholar] [CrossRef]
- Hubert, M. Les Terrains Boisés, Leur Mise en Valeur; IDF: Paris, France, 1999; p. 254. [Google Scholar]
- Amorini, E.; Gambi, G. Il método dell’invecchiamento nella conversione dei cedui di faggio. Ann. Ist. Sper. Selvic. Arezzo 1977, 8, 22–40. [Google Scholar]
- Harmer, R. The thinning of canopy and epicormics shoot growth in Quercus robur L. Forestry 1990, 63, 279–283. [Google Scholar] [CrossRef]
- Matthews, J. Silvicultural Systems; Clarendon Press: Oxford, UK, 1996; p. 283. [Google Scholar]
- Smith, D.; Larson, B.; Kelty, M.; Ashton, P. The Practice of Silviculture; John Wiley and Sons: New York, NY, USA, 1996; p. 560. [Google Scholar]
- Hubert, M. L’Amélioration des Taillis par Balivage Intensif; IDF: Paris, France, 1983; p. 117. [Google Scholar]
- Giannini, R.; Piussi, P. La conversion de taillis en futaie. L’expérience italienne. (The conversion of coppice into high forest. The Italian experience.). In Proceedings of the XVI IUFRO World Congress, Oslo, Norway, 20 June–2 July 1976; pp. 388–396. [Google Scholar]
- Hansen, J.; Spiecker, H.; Teuffel, K. The question of conversion of coniferous forests. In Proceedings of the International Conference Freiburger Forstliche Forschung, Freiburg im Breisgau, Germany, 27 September–2 October 2003; Berichte 47. p. 85. [Google Scholar]
- Notarangelo, M.; La Marca, O.; Moretti, N. Long-term effects of experimental cutting to convert an abandoned oak coppice into transitional high forest in a protected area of the Italian Mediterranean region. For. Ecol. Manag. 2018, 430, 241–249. [Google Scholar] [CrossRef]
- Tulbure, C.; Duduman, G. A conversion method of young hornbeam coppices and its possible impact on future stand structural attributes. Ann. Res. 2012, 55, 281–296. [Google Scholar]
- Ribeiro, A. Contribution à l’Étude Tectonique Tras-os-Montes Oriental; Serviços Geológicos de Portugal: Lisbon, Portugal, 1974; Volume 4, 168p. [Google Scholar]
- Teixeira, C. Geology of Portugal; Calouste Gulbenkian Foundation: Lisbon, Portugal, 1984; p. 630. [Google Scholar]
- Carvalho, J.; Parresol, B. Additivity in tree biomass components of Pyrenean oak (Quercus pyrenaica Willd.). For. Ecol. Manag. 2003, 179, 269–276. [Google Scholar] [CrossRef]
- Carvalho, J.; Parresol, B. A site model for Pyrenean oak (Quercus pyrenaica Willd.) stands using a dynamic algebraic difference equation. Can. J. For. Res. 2004, 35, 93–99. [Google Scholar] [CrossRef]
- Shannon, C.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963; p. 131. [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer: Heidelberg, Germany, 2009; p. 664. [Google Scholar]
- Swaine, M.; Lieberman, D. Note on the calculation of mortality rates. J. Trop. Ecol. 1987, 3, 289–290. [Google Scholar] [CrossRef]
- Steel, R.; Torrie, J. Principles and Procedures of Statistics—A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1986; p. 622. [Google Scholar]
- Zar, J. Biostatistical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1996; p. 662. [Google Scholar]
- Cantiani, P.; Spinelli, R. Conversion to high forest of Turkey oak coppice: Technical and economical assessment of the first conversion stage. Ann. Ist. Sper. Selvic. Arezzo 1998, 27, 191–200. [Google Scholar]
- Picchio, R.; Maesano, M.; Savelli, S.; Marchi, E. Productivity and energy balance in conversion of a Quercus cerris L. coppice stand into high forest in central Italy. Croat. J. For. Eng. 2009, 30, 15–26. [Google Scholar]
- Amorini, E.; Fabbio, G.; Bertini, G. Dinamica del ceduo oltre turno e avviamento ad alto fusto dei cedui di faggio. Ann. Silvic. Res. 2010, 36, 151–172. [Google Scholar]
- Jensen, J.S. Provenance variation in phenotypic traits in Quercus robur and Quercus petraea in Danish provenance trials. Scand. J. For. Res. 2000, 15, 297–308. [Google Scholar] [CrossRef]
- Lanier, L. Précis de Sylviculture; ENGREF: Nancy, France, 1986; p. 468. [Google Scholar]
- Le Goff, N.; Ottorini, J.M.; Ningre, F. Evaluation and comparison of size-density relationships for pure even-aged stands of ash (Fraxinus excelsior L.), beech (Fagus sylvatica L.), oak (Quercus petraea Liebl.), and sycamore maple (Acer pseudoplatanus L.). Ann. For. Sci. 2011, 68, 461–475. [Google Scholar] [CrossRef]
- Carnwath, G.C.; Peterson, D.W.; Nelson, C.R. Effect of crown class and habitat type on climate-growth relationships of ponderosa pine and Douglas-fir. For. Ecol. Manag. 2012, 285, 44–52. [Google Scholar] [CrossRef]
- Wignall, T.; Browning, G.; MacKenzie, A. The physiology of epicormics bud emergence in pedunculate oak (Quercus robur L.). Forestry 1987, 60, 45–56. [Google Scholar] [CrossRef]
- Colin, F.; Robert, N.; Druelle, J.L. Fontaine F Initial spacing has little influence on transient epicormics shoots in a 20-year-old sessile oak plantation. Ann. For. Sci. 2008, 65, 508–517. [Google Scholar] [CrossRef]
Figure 1.
Climatic diagram of the site (Nogueira mountain), with annual precipitation (P), mean temperature (T), mean minimum temperature for the coldest month (m), mean maximum temperature for the hottest month (M), and termicity index (It).
Figure 1.
Climatic diagram of the site (Nogueira mountain), with annual precipitation (P), mean temperature (T), mean minimum temperature for the coldest month (m), mean maximum temperature for the hottest month (M), and termicity index (It).

Figure 2.
Conversion-of-Pyrenean-oak-coppice research trial. From left to right: overview of control plot (a), selective (b) and strip cutting (c), and tree height measurement (d). Images at installation age of 28 years.
Figure 2.
Conversion-of-Pyrenean-oak-coppice research trial. From left to right: overview of control plot (a), selective (b) and strip cutting (c), and tree height measurement (d). Images at installation age of 28 years.


Figure 3.
Diameter (upper) and height (lower) growth of trees over time for different silvicultural interventions in the Pyrenean oak coppice. All interventions provided higher diameter growth compared with the control, although only strip thinning was significantly different. For the tree height, 2 × 2 thinning provided a higher increment, although not significantly different from the other strip thinning.
Figure 3.
Diameter (upper) and height (lower) growth of trees over time for different silvicultural interventions in the Pyrenean oak coppice. All interventions provided higher diameter growth compared with the control, although only strip thinning was significantly different. For the tree height, 2 × 2 thinning provided a higher increment, although not significantly different from the other strip thinning.

Figure 4.
Increment rate index (IRI) for diameter. Relative variation in tree growth in diameter over time since the first measurement (5 years after thinning of 1995). Generally, there is an increasing growth response for 12 years after thinning. After this period, tree growth decreases progressively. The response also varies among the treatments.
Figure 4.
Increment rate index (IRI) for diameter. Relative variation in tree growth in diameter over time since the first measurement (5 years after thinning of 1995). Generally, there is an increasing growth response for 12 years after thinning. After this period, tree growth decreases progressively. The response also varies among the treatments.

Figure 5.
Number and length of epicormic branches of the trees at the end of the period for different treatments of Pyrenean oak coppice. Above: by treatment; below: by tree crown class. Box-plots with confidence intervals. Significant differences between treatments (p < 0.05) are shown with different letters.
Figure 5.
Number and length of epicormic branches of the trees at the end of the period for different treatments of Pyrenean oak coppice. Above: by treatment; below: by tree crown class. Box-plots with confidence intervals. Significant differences between treatments (p < 0.05) are shown with different letters.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General biometric parameters of the experimental trial of Pyrenean oak coppice (before cutting).
Table 1.
General biometric parameters of the experimental trial of Pyrenean oak coppice (before cutting).
| Age (years) | N (trees.ha−1) | hd (m) | hg (m) | dg (cm) | SI50 (m) | HB Index | G (m2.ha−1) | W (t.ha−1) |
|---|---|---|---|---|---|---|---|---|
| 28 | 8865 | 7.8 | 5.9 | 5.6 | 10.7 | 26 | 20.2 | 35.4 |
N: number of trees; hd: dominant height; hg: mean tree height; dg: mean tree diameter; SI50: site index at base age of 50 years (site class IV); HB: Hart-Becking spacing index; G: stand basal area; W: stand biomass, for tree stem (D.W.).
Table 2.
Mean increment for tree diameter (id), tree height (ih), mean crown diameter (idc), and live crown length (ilcl) in the period of 1995–2015. Mean relative proportion (p(%)) in relation to the control plot of Pyrenean oak coppice. The same letter in the column indicates the group subset and nonsignificant differences from the mean comparison test, at the 5% significance level. Different box-filled colors represent relative scores.
Table 2.
Mean increment for tree diameter (id), tree height (ih), mean crown diameter (idc), and live crown length (ilcl) in the period of 1995–2015. Mean relative proportion (p(%)) in relation to the control plot of Pyrenean oak coppice. The same letter in the column indicates the group subset and nonsignificant differences from the mean comparison test, at the 5% significance level. Different box-filled colors represent relative scores.
| Treatment | id (cm) | ih (m) | idc (m) | ilcl (m) | Mean Relative Proportion, p(%) | |||
|---|---|---|---|---|---|---|---|---|
| id | ih | idc | ilcl | |||||
| C | 6.35 a | 3.79 ab | 2.35 a | −0.65 a | 100 a | 100 ab | 100 a | 100 a |
| S | 6.54 a | 3.47 a | 2.45 a | −0.29 a | 103 a | 91 a | 104 a | 118 a |
| 2 × 2 | 7.82 b | 4.05 b | 3.20 b | 0.65 b | 123 b | 107 b | 137 b | 171 b |
| 3 × 3 | 7.46 b | 3.92 ab | 2.55 a | 0.73 b | 117 b | 104 ab | 108 a | 155 b |
| 3 × 6 | 7.38 b | 3.85 ab | 2.75 ab | 0.41 b | 116 b | 101 ab | 117 ab | 147 b |
Table 3.
Mean tree diameter increment (id) for different crown classes (dominant, intermediate, suppressed) in Pyrenean oak coppice. The same letter in the column indicates the group subset and nonsignificant differences from the mean comparison test, at the 5% significance level.
Table 3.
Mean tree diameter increment (id) for different crown classes (dominant, intermediate, suppressed) in Pyrenean oak coppice. The same letter in the column indicates the group subset and nonsignificant differences from the mean comparison test, at the 5% significance level.
| Treatment | Mean Tree Diameter Increment, id (cm) | ||
|---|---|---|---|
| Dominant | Intermediate | Suppressed | |
| C | 7.4 a | 5.5 a | 3.0 a |
| S | 7.9 ab | 6.4 b | 4.1 ab |
| 2 × 2 | 8.7 c | 7.6 c | 4.7 b |
| 3 × 3 | 8.4 bc | 7.3 c | 4.9 b |
| 3 × 6 | 8.5 c | 6.9 bc | 5.1 b |
Table 4.
The dynamic of the tree crown class, vertical structure index, and mortality rate for the different cutting methods in Pyrenean oak coppice.
Table 4.
The dynamic of the tree crown class, vertical structure index, and mortality rate for the different cutting methods in Pyrenean oak coppice.
| Treatment | Mean Changes in Crown Class (%) | Vertical Structure Index (VI) | Mortality Rate | ||
|---|---|---|---|---|---|
| Ascend | Maintain | Descend | (%.y−1) | ||
| C | 19.9 | 74.6 | 5.5 | 0.74 | 1.81 |
| S | 32.7 | 51.8 | 15.5 | 0.58 | 0.61 |
| 2 × 2 | 40.7 | 47.4 | 11.9 | 0.75 | 1.13 |
| 3 × 3 | 27.3 | 64.2 | 8.5 | 0.77 | 1.78 |
| 3 × 6 | 33.3 | 65.0 | 1.7 | 0.53 | 0.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carvalho, J.P.F. Improvement Cuttings in the Conversion of Pyrenean Oak (Quercus pyrenaica Willd.) Coppice. Forests 2023, 14, 575. https://doi.org/10.3390/f14030575
AMA Style
Carvalho JPF. Improvement Cuttings in the Conversion of Pyrenean Oak (Quercus pyrenaica Willd.) Coppice. Forests. 2023; 14(3):575. https://doi.org/10.3390/f14030575
Chicago/Turabian StyleCarvalho, João P. F. 2023. "Improvement Cuttings in the Conversion of Pyrenean Oak (Quercus pyrenaica Willd.) Coppice" Forests 14, no. 3: 575. https://doi.org/10.3390/f14030575
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.