
Variation in Leaf Cutin Content and Chemical Composition along One Annual Cycle in the Mediterranean Cork Oak (Quercus suber L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Variables
2.3. Determination of Cuticular Waxes
2.4. Determination of Cutin Content
2.5. Chemical Composition of Cutin
2.6. Statistical Analysis
3. Results
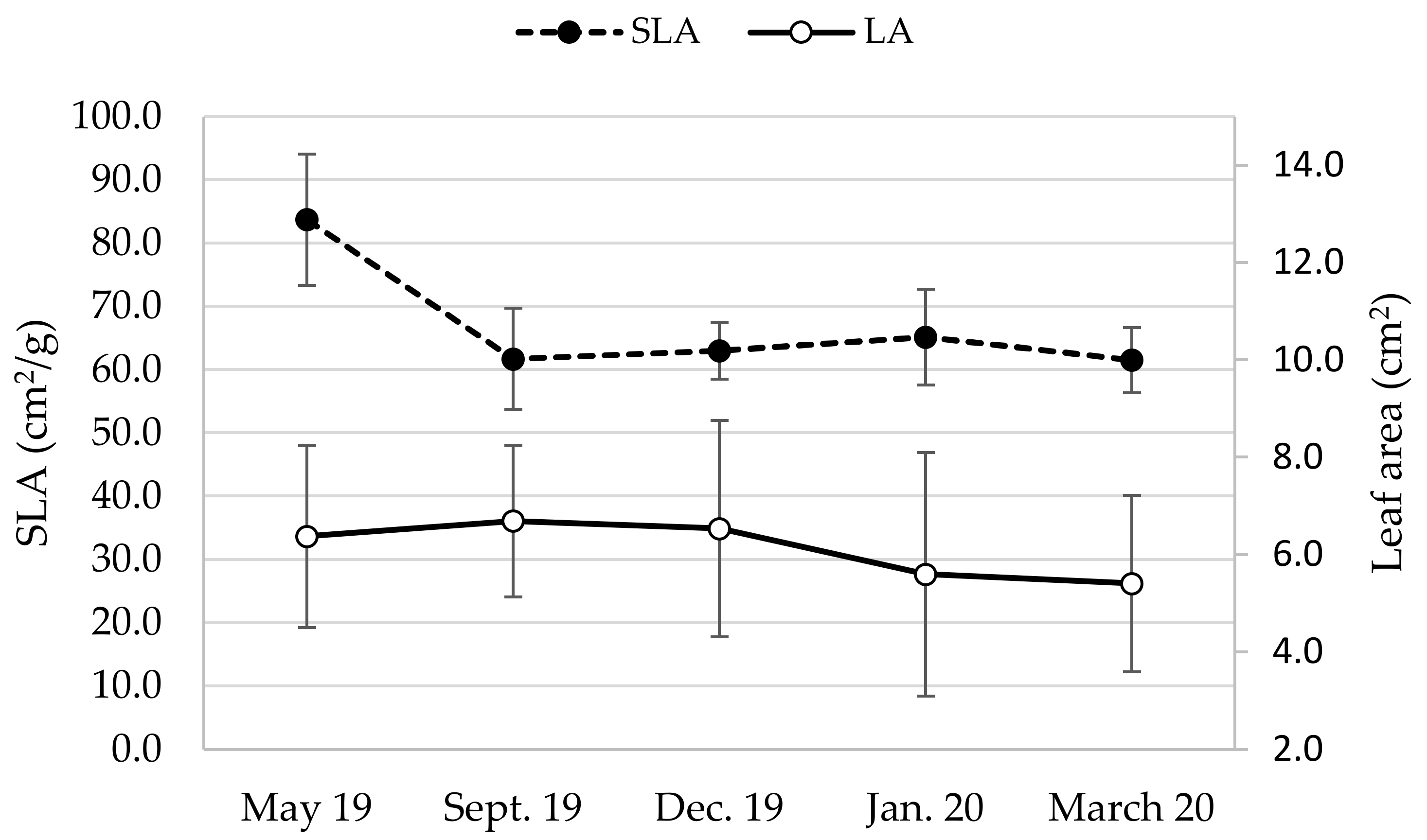
3.1. Leaf Area and Specific Leaf Area
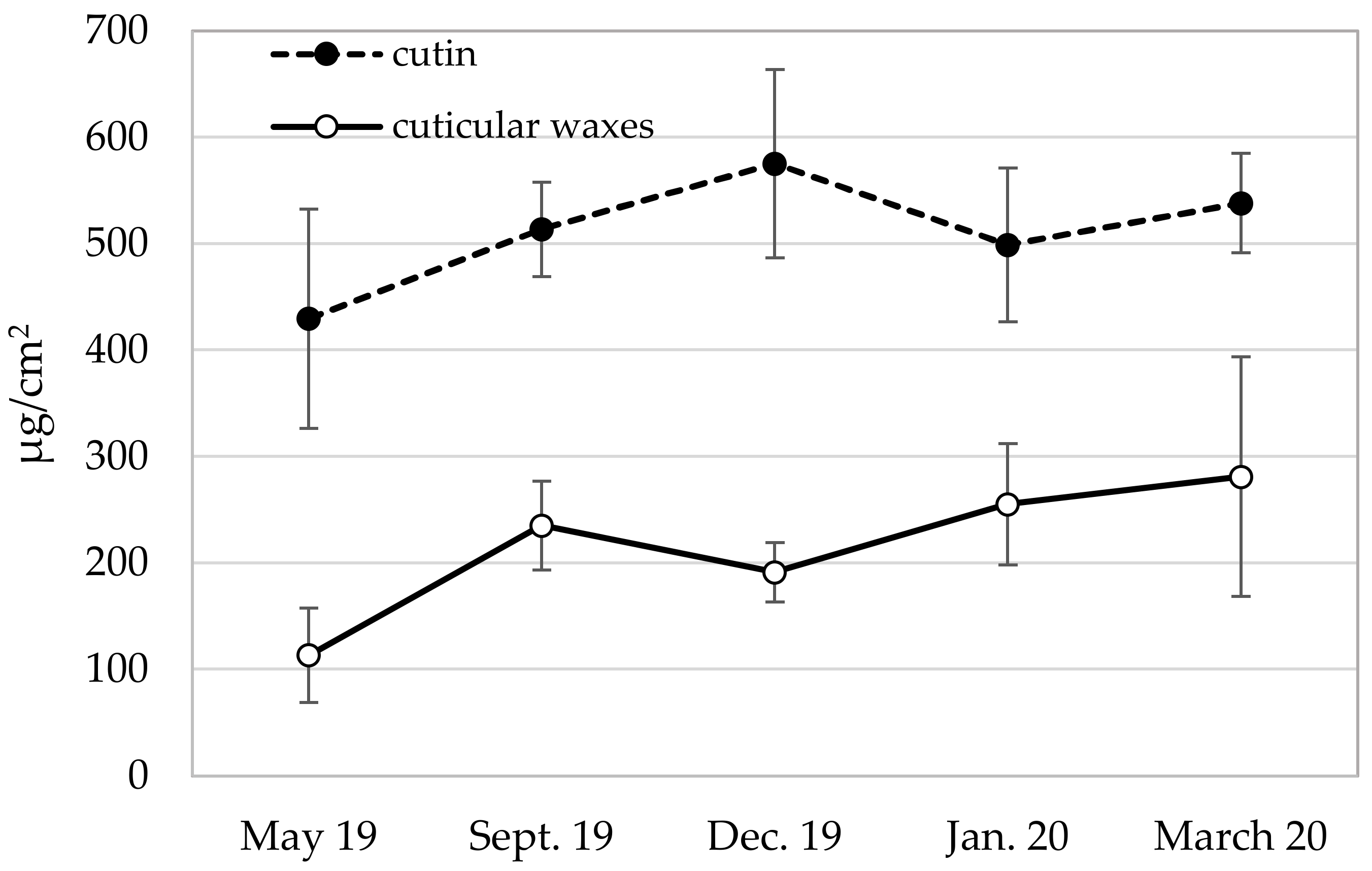
3.2. Cutin and Cuticular Wax Contents
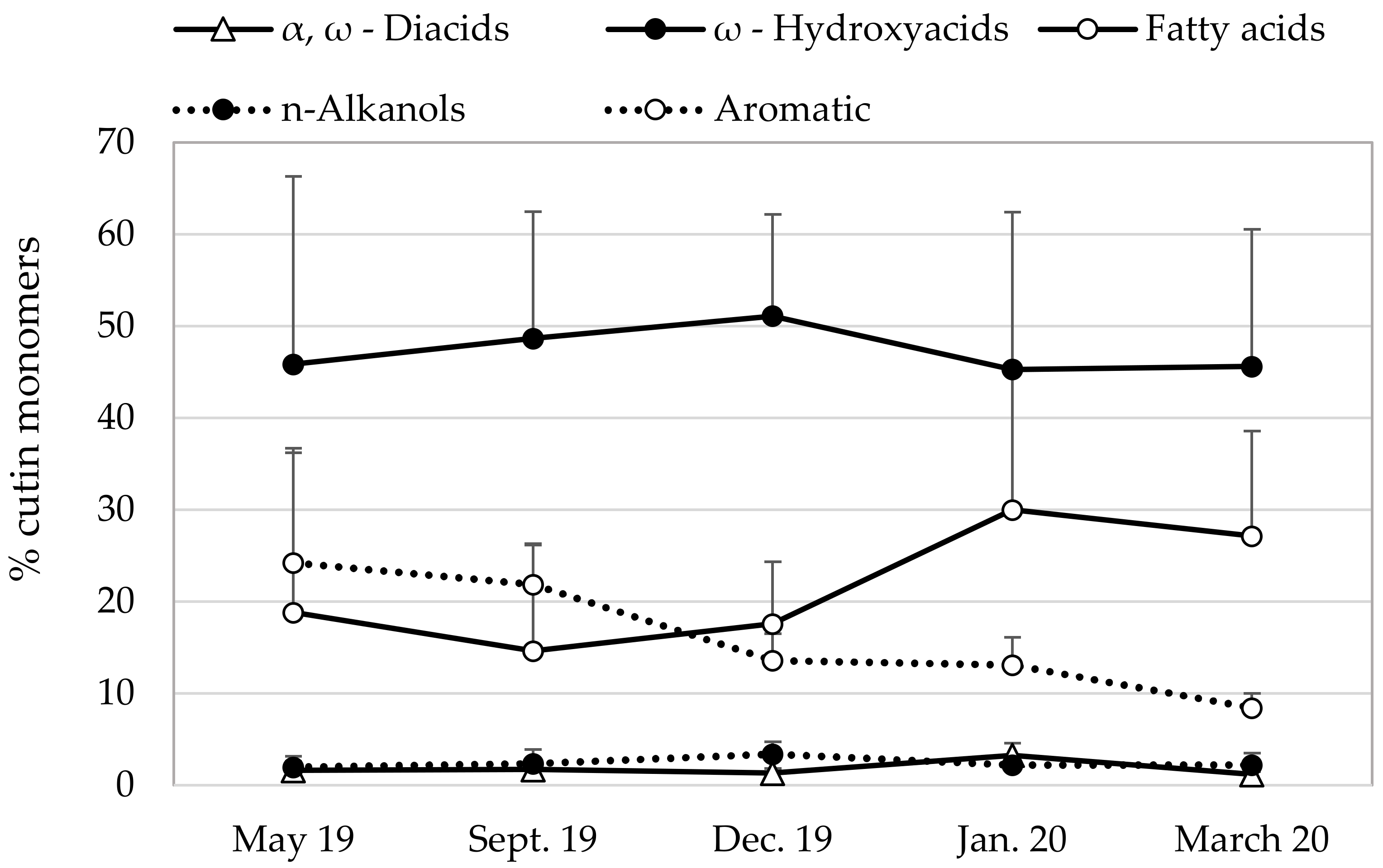
3.3. Cutin Chemical Compostion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kerstiens, G. Cuticular water permeability and its physiological significance. J. Exp. Bot. 1996, 47, 1813–1832. [Google Scholar] [CrossRef]
- Krauss, P.; Markstadter, C.; Riederer, M. Attenuation of UV radiation by plant cuticles from woody species. Plant Cell Environ. 1997, 20, 1079–1085. [Google Scholar] [CrossRef]
- Buschhaus, C.; Jetter, R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Jeffree, C.E. The fine structure of the plant cuticle. In Biology of the Plant Cuticle; Riederer, M., Müller, C., Eds.; Blackwell: Oxford, UK, 2007; pp. 11–125. [Google Scholar] [CrossRef]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing plant surfaces: Cuticular wax formation by epidermal cells. Annu. Rev. Plant Biol. 2008, 59, 683–707. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef]
- Guzmán-Delgado, P.; Graça, J.; Cabral, V.; Gil, L.; Fernández, V. The presence of cutan limits the interpretation of cuticular chemistry and structure: Ficus elastica leaf as an example. Physiol. Plant. 2016, 157, 205–220. [Google Scholar] [CrossRef]
- Leide, J.; Nierop, K.G.J.; Deininger, A.C.; Staiger, S.; Riederer, M.; de Leeuw, J.E. Leaf cuticle analyses: Implications for the existence of cutan/non-ester cutin and its biosynthetic origin. Ann. Bot. 2020, 126, 141–162. [Google Scholar] [CrossRef]
- Zeisler-Diehl, V.; Müller, Y.; Schreiber, L. Epicuticular wax on leaf cuticles does not establish the transpiration barrier, which is essentially formed by intracuticular wax. J. Plant Physiol. 2018, 227, 66–74. [Google Scholar] [CrossRef]
- Buschhaus, C.; Jetter, R. Composition and physiological function of the wax layers coating Arabidopsis leaves: β-amyrin negatively affects the intracuticular water barrier. Plant Physiol. 2012, 160, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Jetter, R.; Riederer, M. Localization of the transpiration barrier in the epi- and intracuticular waxes of eight plant species: Water transport tesistances are associated with fatty acyl rather than alicyclic componentes. Plant Physiol. 2016, 170, 921–934. [Google Scholar] [CrossRef]
- Seufert, P.; Staiger, S.; Arand, K.; Bueno, A.; Burghardt, M.; Riederer, M. Building a barrier: The influence of different wax fractions on the water transpiration barrier of leaf cuticles. Front. Plant Sci. 2022, 12, 766602. [Google Scholar] [CrossRef]
- Schuster, A.-C.; Burghardt, M.; Alfarhan, A.; Bueno, A.; Hedrich, R.; Leide, J.; Thomas, J.; Riederer, M. Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants 2016, 8, plw027. [Google Scholar] [CrossRef]
- Tsubaki, S.; Sugimura, K.; Teramoto, Y.; Yonemori, K.; and Azuma, J.I. Cuticular membrane of Fuyu persimmon fruit is strengthened by triterpenoid nano-fillers. PLoS ONE 2013, 8, 75275. [Google Scholar] [CrossRef]
- Fich, E.A.; Segerson, N.A.; Rose, J.K. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Verdier, G.; Xu, L.; Beisson, F. Cutin and suberin polyesters. In Essential for Life Science (eLS); John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 1–12. [Google Scholar] [CrossRef]
- Pereira, H. Cork: Biology, Production and Uses; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar] [CrossRef]
- Escudero, A.; Del Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of leaf longevity and retranslocation efficiency on the retention time of nutrients in the leaf biomass of different woody species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef]
- Fialho, C.; Lopes, F.; Perreira, H. The effect of cork removal on the radial growth and phenology of young cork oak trees. For. Ecol. Manag. 2001, 141, 251–258. [Google Scholar] [CrossRef]
- Oliveira, G.; Correia, O.; Martins-Loução, M.A.; Catarino, F. Phenological and growth patterns of the Mediterranean oak Quercus suber L. Trees 1994, 9, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.S.; Beyschlag, G.; Lange, O.L.; Beyschlag, W.; Tenhunen, J.D. Comparative phenology of four mediterranean shrub species growing in Portugal. In Plant Response to Stress; Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 503–514. [Google Scholar]
- Simões, R.; Rodrigues, A.; Ferreira-Dias, S.; Miranda, I.; Pereira, H. Chemical composition of cuticular waxes, pigments and morphology of leaves of Quercus suber trees from different provenances. Plants 2020, 9, 1165. [Google Scholar] [CrossRef]
- Simões, R.; Miranda, I.; Pereira, H. Chemical composition of leaf cutin in six Quercus suber provenances. Phytochemistry 2021, 181, 112570. [Google Scholar] [CrossRef]
- Simões, R.; Miranda, I.; Pereira, H. Effect of seasonal variation on leaf cuticular waxes’ composition in the Mediterranean cork oak (Quercus suber L.). Forests 2022, 13, 1236. [Google Scholar] [CrossRef]
- Sampaio, T.; Gonçalves, E.; Patrício, M.S.; Cota, T.M.; Almeida, M.H. Seed origin drives differences in survival and growth traits of cork oak (Quercus suber L.) populations. For. Ecol. Manag. 2019, 448, 267–277. [Google Scholar] [CrossRef]
- Varela, M.C. European Network for the Evaluation of Genetic Resources of Cork Oak for Appropriate Use in Breeding and Gene Conservation Strategies; INIA: Lisbon, Portugal, 2020. [Google Scholar]
- Pereira, H. Chemical composition and variability of cork from Quercus suber L. Wood Sci. Technol. 1988, 22, 211–218. [Google Scholar] [CrossRef]
- Grant, O.M.; Tronina, L.; Ramalho, J.C.; Besson, C.K.; Lobo-Do-Vale, R.; Pereira, J.S.; Jones, H.G.; Chaves, M.M. The impact of drought on leaf physiology of Quercus suber L. trees: Comparison of an extreme drought event with chronic rainfall reduction. J. Exp. Bot. 2010, 61, 4361–4371. [Google Scholar] [CrossRef]
- Prats, K.A.; Brodersen, C.R.; Ashton, M.S. Influence of dry season on Quercus suber L. leaf traits in the Iberian Peninsula. Am. J. Bot. 2019, 106, 656–666. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Hobbs, R.J.; Mayfield, M.M. Specific leaf area responses to environmental gradients through space and time. Ecology 2014, 95, 399–410. [Google Scholar] [CrossRef]
- Aranda, I.; Pardos, M.; Puértolas, J.; Jiménez, M.D.; Pardos, J.A. Water-use efficiency in cork oak (Quercus suber) is modified by the interaction of water and light availabilities. Tree Physiol. 2007, 27, 671–677. [Google Scholar] [CrossRef]
- Gouveia, A.C.; Freitas, H. Modulation of leaf attributes and water use efficiency in Quercus suber along a rainfall gradient. Trees 2009, 23, 267–275. [Google Scholar] [CrossRef]
- Passarinho, J.A.P.; Lamosa, P.; Baeta, J.P.; Santos, H.; Ricardo, C.P.P. Annual changes in the concentration of minerals and organic compounds of Quercus suber leaves. Physiol. Plant 2006, 127, 100–110. [Google Scholar] [CrossRef]
- Gülz, P.-G.; Müller, E. Seasonal variation in the composition of epicuticular waxes of Quercus robur leaves. Z. Naturforsch. C. 1992, 47, 800–806. [Google Scholar] [CrossRef]
- Prasad, R.B.N.; Gülz, P.G. Surface structure and chemical composition of leaf waxes from Quercus robur L., Acer pseudoplatanus L. and JugIans regia L. Z. Naturforsch. C. 1990, 45, 813–817. [Google Scholar] [CrossRef]
- Hauke, V.; Schreiber, L. Ontogenetic and seasonal development of wax composition and cuticular transpiration of ivy (Hedera helix L.) sun and shade leaves. Planta 1998, 207, 67–75. [Google Scholar] [CrossRef]
- Celano, G.; D’Auria, M.; Xiloyannis, C.; Mauriello, G.; Baldassarre, M. Composition and seasonal variation of soluble cuticular waxes in Actinidia deliciosa leaves. Nat. Prod. Res. 2006, 20, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Broeckling, C.D.; Blancaflor, E.B.; Sledge, M.K.; Sumner, L.W.; Wang, Z.Y. Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa). Plant J. 2005, 42, 689–707. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumu- lation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Jetter, R.; Schäffer, S. Chemical composition of the Prunus laurocerasus leaf surface. dynamic changes of the epicuticular wax film during leaf development. Plant Physiol. 2001, 126, 1725–1737. [Google Scholar] [CrossRef]
- Neinhuis, C.; Koch, K.; Barthlott, W. Movement and regeneration of epicuticlar waxes through plant cuticles. Planta 2001, 213, 427–434. [Google Scholar] [CrossRef]
- Riley, R.G.; Kolattukudy, P.E. Evidence for covalently attached p-voumaric acid and ferulic acid in cutins and suberins. Plant Physiol. 1975, 56, 650–654. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| May 2019 | September 2019 | December 2019 | January 2020 | March 2020 | |
|---|---|---|---|---|---|
| Cutin (g/1000 g dry leaves) | 71.0 ± 16.2 | 66.2 ± 6.2 | 71.2 ± 8.7 | 63.5 ± 6.8 | 65.5 ± 1.6 |
| Cutin (% of cuticle) | 79.3 ± 5.5 | 70.1 ± 5.0 | 75.7 ± 4.9 | 66.3 ± 4.7 | 70.4 ± 3.4 |
| Cuticular waxes (g/1000 g dry leaves) | 18.4 ± 5.8 | 28.5 ± 6.0 | 22.8 ± 4.7 | 32.8 ± 8.6 b | 27.6 ± 4.0 |
| Cuticular waxes (% of cuticle) | 20.7 ± 5.5 | 29.9 ± 5.0 | 24.3 ± 4.9 | 33.7 ± 4.7 | 29.6 ± 3.4 |
| Cuticle (cutin + cuticular waxes) (g/1000 g dry leaves) | 89.3 ± 18.3 | 94.6 ± 9.0 | 94.0 ± 8.4 | 96.3 ± 13.8 | 93.1 ± 3.3 |
| May 2019 | September 2019 | December 2019 | January 2020 | March 2020 | |
|---|---|---|---|---|---|
| n-Alkanols | |||||
| Hexadecan-1-ol | 0.24 ± 0.12 | 0.65 ± 0.48 | 1.00 ± 0.54 | 0.77 ± 0.45 | 0.84 ± 0.51 |
| Octadecan-1-ol | 0.21 ± 0.08 | 0.26 ± 0.13 | 0.15 ± 0.08 | 0.10 ± 0.03 | 0.16 ± 0.06 |
| Eicosan-1-ol | 0.17 ± 0.09 | 0.14 ± 0.09 | 0.14 ± 0.08 | 0.13 ± 0.05 | 0.22 ± 0.33 |
| Docosan-1-ol | 0.22 ± 0.10 | 0.16 ± 0.06 | 0.18 ± 0.04 | 0.15 ± 0.02 | 0.21 ± 0.06 |
| Tetracosan-1-ol | 0.81 ± 0.42 | 0.44 ± 0.23 | 1.17 ± 0.30 | 0.65 ± 0.36 | 0.39 ± 0.15 |
| Hexacosan-1-ol | 0.16 ± 0.21 | 0.40 ± 0.26 | 0.34 ± 0.12 | 0.17 ± 0.03 | - |
| 1,2-Dodecanediol | 0.04 ± 0.07 | 0.25 ± 0.23 | 0.27 ± 0.09 | 0.12 ± 0.09 | 0.27 ± 0.11 |
| Alkanes | |||||
| Heptacosane | 0.22 ± 0.01 | 0.06 ± 0.05 | 0.06 ± 0.03 | 0.06 ± 0.01 | 0.07 ± 0.04 |
| Nonacosane | 0.08 ± 0.01 | 0.21 ± 0.16 | 0.08 ± 0.04 | 0.17 ± 0.17 | 0.39 ± 0.34 |
| Triacontane | – | 0.11 ± 0.06 | 0.06 ± 0.01 | 0.06 ± 0.02 | 0.08 ± 0.03 |
| α, ω–Diacids | |||||
| Butanedioic acid | 0.03 ± 0.02 | 0.24 ± 0.22 | 0.02 ± 0.01 | 0.67 ± 0.20 | 0.04 ± 0.01 |
| Nonanedioic acid | 0.58 ± 0.39 | – | 0.61 ± 0.25 | – | 0.74 ± 0.34 |
| Decanedioic acid | 1.12 ± 0.66 | 0.45 ± 0.33 | 0.13 ± 0.02 | 9.99 ± 5.40 | 6.50 ± 3.78 |
| Hexadecanedioic acid | 0.40 ± 0.16 | 0.55 ± 0.21 | 0.50 ± 0.12 | 0.31 ± 0.19 | 0.47 ± 0.11 |
| Eicosanedioic acid | 0.54 ± 0.45 | 0.37 ± 0.28 | – | – | – |
| Octadecanedioic acid | 0.08 ± 0.03 | 0.08 ± 0.02 | 0.13 ± 0.02 | 0.13 ± 0.06 | 0.15 ± 0.02 |
| Tetracosanedioic acid | 0.07 ± 0.03 | 0.11 ± 0.05 | 0.05 ± 0.02 | 0.07 ± 0.04 | 0.08 ± 0.05 |
| Cis-4-Decene-1,10-dioic acid | 0.02 ± 0.01 | – | 0.10 ± 0.05 | – | 0.04 ± 0.01 |
| 9-Octadecenedioic acid 1,18-dimethyl ester | 0.14 ± 0.13 | 0.24 ± 0.12 | 0.15 ± 0.09 | 0.13 ± 0.06 | 0.10 ± 0.03 |
| 10,12-Docosadiynedioic acid | 0.09 ± 0.04 | – | 0.04 ± 0.01 | 2.10 ± 1.77 | – |
| 9,10-Dihydroxyoctadecanedioic acid | 0.31 ± 0.17 | 0.38 ± 0.16 | 0.43 ± 0.26 | 0.54 ± 0.61 | 0.39 ± 0.09 |
| 2-Hydroxydecanedioic acid | 0.48 ± 0.43 | – | 0.17 ± 0.11 | 0.34 ± 0.34 | 0.30 ± 0.15 |
| 2-Hydroxy-3-isopropylsuccinic acid | 0.13 ± 0.05 | 0.17 ± 0.11 | 0.07 ± 0.03 | 0.07 ± 0.03 | 0.11 ± 0.05 |
| ω–Hydroxyacids | |||||
| 16-Hydroxyhexadecanoic acid | 1.24 ± 1.02 | 1.82 ± 1.22 | 2.66 ± 0.83 | 1.41 ± 0.92 | 2.20 ± 0.27 |
| 22-Hydroxydocosanoic acid | 0.37 ± 0.29 | 0.45 ± 0.24 | 1.43 ± 0.79 | 2.87 ± 2.15 | 2.02 ± 1.05 |
| 9-Octadecenoic acid 18-hydroxy- methyl ester | 1.15 ± 1.78 | 2.69 ± 1.82 | 2.08 ± 1.17 | 1.09 ± 0.69 | 0.50 ± 0.40 |
| 10,16-Dihydroxyhexadecanoic acid | 29.28 ± 8.32 | 25.93 ± 4.15 | 26.47 ± 3.92 | 20.22 ± 3.22 | 18.39 ± 3.46 |
| 9,10-Epoxy-18-hydroxyoctadecanoic acid | 2.30 ± 2.09 | 2.14 ± 1.91 | 6.51 ± 2.63 | 0.49 ± 0.25 | 8.96 ± 3.82 |
| 9,16-Dihydroxyhexadecanoic acid | 0.49 ± 0.25 | 0.30 ± 0.16 | 0.43 ± 0.13 | 9.59 ± 5.63 | 1.41 ± 3.41 |
| 9,12-Octadecadienoic acid 18-hydroxy- methyl ester | 0.60 ± 0.61 | 0.30 ± 0.26 | 0.24 ± 0.10 | 0.18 ± 0.11 | 0.50 ± 0.32 |
| 9,10,18 Trihydroxyoctadecanoic acid | 10.37 ± 6.08 | 14.97 ± 4.07 | 11.23 ± 1.49 | 9.41 ± 4.15 | 11.57 ± 2.22 |
| Carboxylic acids | |||||
| Ethanoic acid | 0.55 ± 0.42 | – | 0.13 ± 0.11 | 0.37 ± 0.22 | 0.23 ± 0.12 |
| Octanoic acid | 2.34 ± 3.17 | 0.51 ± 0.69 | 0.52 ± 0.72 | 0.47 ± 0.90 | 0.84 ± 0.78 |
| Nonanoic acid | 0.18 ± 0.09 | 0.06 ± 0.11 | 0.17 ± 0.05 | 0.10 ± 0.11 | 0.26 ± 0.09 |
| Dodecanoic acid | 0.20 ± 0.27 | 0.07 ± 0.02 | 0.08 ± 0.06 | 0.12 ± 0.10 | 0.08 ± 0.05 |
| Tetradecanoic acid | 0.16 ± 0.19 | 0.13 ± 0.21 | 0.30 ± 0.10 | 0.23 ± 0.21 | 0.42 ± 0.10 |
| Hexadecanoic acid | 3.24 ± 2.52 | 2.68 ± 1.38 | 5.49 ± 0.98 | 5.86 ± 1.74 | 7.25 ± 1.32 |
| Octadecanoic acid | 0.31 ± 0.20 | 0.43 ± 0.30 | 0.34 ± 0.10 | 0.26 ± 0.19 | 0.41 ± 0.20 |
| Eicosanoic acid | 0.21 ± 0.10 | 0.32 ± 0.20 | 0.29 ± 0.08 | 0.22 ± 0.14 | 0.24 ± 0.10 |
| Docosanoic acid | 0.49 ± 0.29 | 0.31 ± 0.26 | 0.23 ± 0.15 | 0.29 ± 0.11 | – |
| Tetracosanoic acid | 1.22 ± 1.01 | 0.38 ± 0.10 | 0.26 ± 0.07 | 0.50 ± 0.15 | 0.69 ± 0.36 |
| Octacosanoic acid | 0.07 ± 0.04 | 0.17 ± 0.17 | 0.10 ± 0.04 | 0.13 ± 0.14 | 0.11 ± 0.04 |
| Triacontanoic acid | 0.16 ± 0.13 | 0.29 ± 0.28 | 0.23 ± 0.07 | 0.20 ± 0.19 | 0.31 ± 0.12 |
| 7-Hexadecenoic acid (Z)- | – | 0.03 ± 0.01 | 0.06 ± 0.02 | – | 0.12 ± 0.15 |
| 9-Hexadecenoic acid (Z)- | – | 0.14 ± 0.19 | 0.03 ± 0.01 | 0.07 ± 0.01 | 0.17 ± 0.44 |
| 9,12,15-Octadecatrienoic acid (Z,Z,Z)- | 0.42 ± 0.85 | 0.85 ± 0.71 | 0.39 ± 0.28 | 1.36 ± 1.33 | 0.30 ± 0.27 |
| 9,12-Octadecadienoic acid (Z,Z)- | 0.38 ± 0.62 | 2.09 ± 2.28 | – | 0.40 ± 0.20 | 0.10 ± 0.08 |
| 9-Octadecenoic acid (Z)- | 0.20 ± 0.21 | 1.35 ± 1.81 | 1.39 ± 1.01 | 1.35 ± 0.73 | 0.09 ± 0.08 |
| 3,6,9-Octadecatrienoic acid | 0.30 ± 0.38 | 0.23 ± 0.12 | – | 0.15 ± 0.04 | 0.11 ± 0.02 |
| 13-Octadecenoic acid | – | – | 0.15 ± 0.07 | 0.12 ± 0.06 | 0.17 ± 0.06 |
| 5,8,11-Eicosatriynoic acid | 1.96 ± 2.66 | 0.20 ± 0.18 | 0.83 ± 0.47 | 1.34 ± 0.95 | 1.51 ± 0.46 |
| 2-Oxobutanoic acid | 0.11 ± 0.04 | 0.13 ± 0.11 | 0.05 ± 0.03 | 0.07 ± 0.03 | 0.04 ± 0.02 |
| 9-Oxononanoic acid | 0.13 ± 0.07 | 0.14 ± 0.02 | 0.10 ± 0.04 | 0.13 ± 0.07 | 0.24 ± 0.08 |
| Methyl 10-oxo-8-decenoate | 0.07 ± 0.04 | 0.30 ± 0.12 | 0.06 ± 0.04 | 0.15 ± 0.08 | 0.36 ± 0.15 |
| 12-Methyl tetradecanoic acid | 0.08 ± 0.04 | 0.07 ± 0.02 | 0.08 ± 0.08 | 0.06 ± 0.01 | 0.13 ± 0.05 |
| 14-Methyl hexadecanoic acid | 0.08 ± 0.02 | 0.09 ± 0.03 | 0.15 ± 0.06 | 0.11 ± 0.04 | 0.14 ± 0.04 |
| 7-Hydroxyoctanoic acid | 0.22 ± 0.15 | 0.19 ± 0.11 | 0.27 ± 0.12 | 0.20 ± 0.09 | 0.28 ± 0.09 |
| 21-Methyl docosanoic acid | 0.11 ± 0.04 | 0.07 ± 0.07 | 0.14 ± 0.01 | 0.16 ± 0.04 | 0.19 ± 0.07 |
| 2-Hydroxytetradecanoic acid | 2.87 ± 2.07 | 2.50 ± 1.23 | 4.15 ± 1.03 | 4.16 ± 1.48 | 4.07 ± 1.24 |
| 2-Hydroxypentanoic acid | – | – | 0.30 ± 0.41 | 0.18 ± 0.06 | 0.35 ± 0.40 |
| Aromatic compounds | |||||
| Methyl p-coumarate | 22.99 ± 12.0 | 19.58 ± 4.07 | 11.35 ± 2.01 | 11.79 ± 2.61 | 6.29 ± 1.23 |
| Other aromatic compounds | 1.09 ± 0.42 | 2.14 ± 0.36 | 2.04 ± 0.84 | 1.24 ± 0.36 | 2.14 ± 0.36 |
| Glycerol + derivatives | |||||
| Ethylene glycol | 0.13 ± 0.10 | 0.21 ± 0.16 | 0.04 ± 0.05 | 0.12 ± 0.10 | 0.07 ± 0.05 |
| Glycerol | 0.02 ± 0.01 | – | 0.03 ± 0.02 | 0.05 ± 0.02 | 0.10 ± 0.06 |
| Oleoylglycerol | 0.20 ± 0.17 | 0.22 ± 0.18 | 0.44 ± 0.16 | 0.32 ± 0.18 | 0.18 ± 0.06 |
| Monopalmitin | – | 0.26 ± 0.34 | – | – | – |
| 9,12-Octadecadienoic acid (Z,Z)- 2,3-dihydroxypropyl ester | 0.22 ± 0.10 | 0.29 ± 0.21 | 0.30 ± 0.18 | 0.25 ± 0.10 | 0.37 ± 0.14 |
| Terpernes | |||||
| Phytol | – | 0.10 ± 0.10 | 0.11 ± 0.05 | 0.11 ± 0.04 | – |
| β-Amyrin | 0.24 ± 0.16 | 0.35 ± 0.20 | 0.26 ± 0.16 | 0.35 ± 0.27 | 0.26 ± 0.09 |
| α-Amyrin | 0.16 ± 0.12 | 0.10 ± 0.10 | 0.22 ± 0.14 | 0.10 ± 0.06 | 0.17 ± 0.15 |
| Germanicol | 0.15 ± 0.17 | 0.13 ± 0.07 | 0.14 ± 0.14 | 0.27 ± 0.26 | 0.16 ± 0.13 |
| Lupeol | – | 0.33 ± 0.10 | 0.25 ± 0.18 | 0.14 ± 0.10 | 0.24 ± 0.20 |
| D:A-Friedooleanan-7-ol (7α)- | 0.07 ± 0.02 | 0.36 ± 0.39 | 0.09 ± 0.03 | – | 0.08 ± 0.04 |
| Friedelan-3-one | – | 0.19 ± 0.10 | 0.05 ± 0.02 | 0.07 ± 0.04 | 0.09 ± 0.09 |
| Betulin | 0.11 ± 0.11 | 0.08 ± 0.01 | – | – | 0.04 ± 0.02 |
| Steroids | |||||
| Epimethendiol | 0.07 ± 0.03 | 0.11 ± 0.03 | 0.14 ± 0.14 | 0.12 ± 0.06 | 0.11 ± 0.13 |
| 12α-Hydroxy-5α-pregnane- dimethyl(isopropyl)silyl ether | 0.74 ± 0.45 | 1.01 ± 0.43 | 1.29 ± 0.24 | 0.88 ± 0.21 | 1.08 ± 0.29 |
| β-Sistosterol | – | 0.46 ± 0.36 | 0.32 ± 0.24 | 0.14 ± 0.10 | 0.27 ± 0.13 |
| Vitamin E | |||||
| α-Tocospiro A | 0.14 ± 0.18 | 0.13 ± 0.10 | 0.10 ± 0.05 | 0.28 ± 0.28 | 0.39 ± 0.73 |
| α-Tocospiro B | 0.10 ± 0.02 | 0.11 ± 0.05 | 0.11 ± 0.05 | 0.11 ± 0.04 | 0.11 ± 0.04 |
| Other compounds | |||||
| 2-Pentadecanone 6,10,14-trimethyl- | – | 0.18 ± 0.12 | 0.10 ± 0.04 | 0.12 ± 0.05 | 0.14 ± 0.09 |
| D-Pinitol | 0.04 ± 0.01 | – | 0.08 ± 0.04 | 0.05 ± 0.05 | 0.10 ± 0.11 |
| Sitosteryl-3β-D-glucopiranoside | 0.08 ± 0.01 | 0.17 ± 0.11 | – | 0.021 ± 0.002 | 0.04 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, R.; Miranda, I.; Pereira, H. Variation in Leaf Cutin Content and Chemical Composition along One Annual Cycle in the Mediterranean Cork Oak (Quercus suber L.). Forests 2023, 14, 334. https://doi.org/10.3390/f14020334
Simões R, Miranda I, Pereira H. Variation in Leaf Cutin Content and Chemical Composition along One Annual Cycle in the Mediterranean Cork Oak (Quercus suber L.). Forests. 2023; 14(2):334. https://doi.org/10.3390/f14020334
Chicago/Turabian StyleSimões, Rita, Isabel Miranda, and Helena Pereira. 2023. "Variation in Leaf Cutin Content and Chemical Composition along One Annual Cycle in the Mediterranean Cork Oak (Quercus suber L.)" Forests 14, no. 2: 334. https://doi.org/10.3390/f14020334