

Relationship between the Floristic Composition and Soil Characteristics of a Tropical Rainforest (TRF)
 , , ,
, , ,  ,
,  , and
, and .jpg)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Plant and Soil Sampling
2.3. Plant Analysis
2.4. Soil Analysis
2.4.1. Statistical Analysis
2.4.2. Data Processing and Analysis to Assess the Correlation between Dendrometric Parameters and Soil Chemical Properties of Plots
3. Results
3.1. Floristic Composition
3.2. Species Importance
3.2.1. Relative Density
3.2.2. Relative Frequency
3.2.3. Basal Area
3.2.4. Importance Value Index
3.3. Soil Characteristics
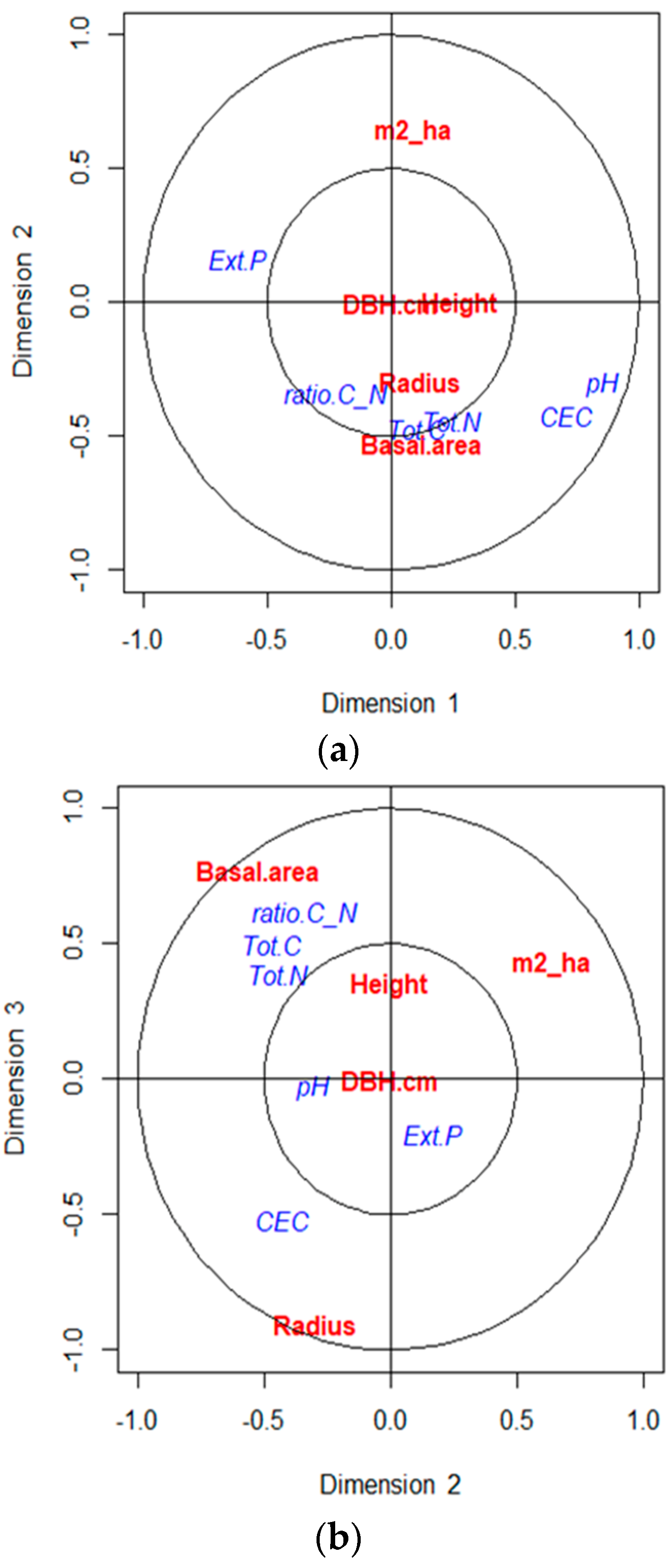
3.4. Correlation between Dendrometric Parameters and Soil Chemical Properties of Plots
4. Discussion
4.1. Floristic Composition
4.2. Soil Characteristics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.N.; Badshah, L. Floristic diversity and utility of flora of district Charsadda, Khyber Pakhtunkhwa. Acta Ecol. Sin. 2019, 39, 306–320. [Google Scholar]
- Khan, M.N.; Ali, S.; Razak, S.A.; Zaman, A.; Iqbal, M.; Shah, S.N. Assessment of floristic diversity in the mountain ecosystem of Marghazar Valley, Hindukush Range, Swat, Pakistan. Biodivers. J. Biol. Divers. 2022, 23, 1000–1013. [Google Scholar]
- Ibrahim, M.; Khan, M.; Ali, S.; Razzaq, A.; Zaman, A.; Iqbal, M.; Jan, F. Floristic composition and species diversity of plant resources of rural area “Takht Bhai” district Mardan, Khyber Pakhtunkhwa, Pakistan. Med. Aromat. Plants 2019, 8, 2167-0412. [Google Scholar]
- Khan, M.N.; Ali, S.; Yaseen, T.; Ullah, S.; Zaman, A.; Iqbal, M.; Shah, S. Eco-Taxonomic Study of Family Poaceae (Gramineae). RADS J. Biol. Res. Appl. Sci. 2019, 10, 63–75. [Google Scholar]
- Dossou-Yovo, E.R.; Devkota, K.P.; Akpoti, K.; Danvi, A.; Duku, C.; Zwart, S.J. Thirty years of water management research for rice in sub-Saharan Africa: Achievement and perspectives. Field Crops Res. 2022, 283, 108548. [Google Scholar]
- Dossou-Yovo, H.O.; Vodouhè, F.G.; Kaplan, A.; Sinsin, B. Application of ethnobotanical indices in the utilization of five medicinal herbaceous plant species in Benin, West Africa. Diversity 2022, 14, 612. [Google Scholar]
- Karam, D.S.; Arifin, A.; Radziah, O.; Shamshuddin, J.; Hazandy, A.-H.; Majid, N.M.; Mohanaselvi, P.; Nor, H. Assessing soil biological properties of natural and planted forests in the Malaysian tropical lowland dipterocarp forest. Am. J. Appl. Sci. 2011, 8, 854–859. [Google Scholar]
- Nesha, M.K.; Herold, M.; De Sy, V.; Duchelle, A.E.; Martius, C.; Branthomme, A.; Garzuglia, M.; Jonsson, O.; Pekkarinen, A. An assessment of data sources, data quality and changes in national forest monitoring capacities in the Global Forest Resources Assessment 2005–2020. Environ. Res. Lett. 2021, 16, 054029. [Google Scholar]
- Saiful, I.; Latiff, A. Effects of selective logging on tree species composition, richness and diversity in a hill dipterocarp forest in Malaysia. J. Trop. For. Sci. 2014, 26, 188–202. [Google Scholar]
- Krishnapillay, D.B.; Razak, M.A.; Appanah, S. Forest rehabilitation–the malaysian experience. Keep Asia Green 2007, 1, 85–123. [Google Scholar]
- Suratman, M.N. Tree species diversity and forest stand structure of Pahang National Park, Malaysia. Biodivers. Enrich. Divers. World 2012, 19, 45–56. [Google Scholar]
- Tan, K.H. Soil Sampling, Preparation, and Analysis; CRC press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Mexia, T.; Vieira, J.; Príncipe, A.; Anjos, A.; Silva, P.; Lopes, N.; Freitas, C.; Santos-Reis, M.; Correia, O.; Branquinho, C. Ecosystem services: Urban parks under a magnifying glass. Environ. Res. 2018, 160, 469–478. [Google Scholar]
- Curtis, J.T. The Vegetation of Wisconsin: An Ordination of Plant Communities; University of Wisconsin Press: Madison, WI, USA, 1959. [Google Scholar]
- Sarah, A. A Phytosociological Study of Aquilaria Malaccensis Lamk. and Its Communities at Sungai Udang Forest Reserve. Ph.D. Thesis, University of Malaya, Kuala Lumpur, Malaysia, 2016. [Google Scholar]
- Millet, J.; Pascal, J.; Kiet, L. Effects of disturbance over 60 years on a lowland forest in southern Vietnam. J. Trop. For. Sci. 2010, 22, 237–246. [Google Scholar]
- Sato, H. Simulation of the vegetation structure and function in a Malaysian tropical rain forest using the individual-based dynamic vegetation model SEIB-DGVM. For. Ecol. Manag. 2009, 257, 2277–2286. [Google Scholar]
- di Pahang, R. Floristic variation of tree communities in two distinct habitats within a forest park in Pahang, Peninsular Malaysia. Sains Malays. 2012, 41, 1–10. [Google Scholar]
- Wang, P.-C.; Xu, Y.-R.; Zhou, Z.-X. Influence of anthropogenic disturbances on understory plant diversity of urban forests in Wuhan, Central China. Sains Malays. 2012, 41, 1495–1501. [Google Scholar]
- di Negeri Sembilan, R. Association of liana communities with their soil properties in a lowland forest of Negeri Sembilan, Peninsular Malaysia. Sains Malays. 2012, 41, 679–690. [Google Scholar]
- Norbaiah, M.; Nazre, M.; Kamziah, A.K.; Nur, N.M.I. Tree health status of mixed species in urban forest of Taman Rimba Alam, Putrajaya, Malaysia. Malays. Appl. Biol. J. 2017, 46, 91–101. [Google Scholar]
- Tans, S.J.; Verschueren, A.R.; Dekker, C. Room-temperature transistor based on a single carbon nanotube. Nature 1998, 393, 49–52. [Google Scholar]
- Husch, B.; Beers, T.W.; Kershaw, J.A., Jr. Forest Mensuration; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Hayat, M.A.; Abd Kudus, K. Assessment of plant species diversity at pasir tengkorak forest reserve, Langkawi Island, Malaysia. J. Agric. Sci. 2010, 2, 31. [Google Scholar]
- Soltis, D.; Soltis, P.; Endress, P.; Chase, M.W.; Manchester, S.; Judd, W.; Majure, L.; Mavrodiev, E. Phylogeny and Evolution of the Angiosperms: Revised and Updated Edition; University of Chicago Press: Chicago, IL, USA, 2018. [Google Scholar]
- Othman, Y.; Shamshuddin, J. Sains Tanah; Dewan Bahasa dan Pustaka, Kuala Lumpur: Kuala Lumpur, Malaysia, 1982. [Google Scholar]
- Hallbäcken, L.; Tamm, C.O. Changes in soil acidity from 1927 to 1982–1984 in a forest area of south-west Sweden. Scand. J. For. Res. 1986, 1, 219–232. [Google Scholar]
- McLean, E. Soil pH and lime requirement. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; American Society of Agronomy, Inc.: Madison, WI, USA, 1983; Volume 9, pp. 199–224. Available online: https://acsess.onlinelibrary.wiley.com/doi/book/10.2134/agronmonogr9.2.2ed (accessed on 26 November 2022).
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis: Part 3 Chemical Methods; ASA-SSSA: Madison, WI, USA, 1996; Volume 5, pp. 475–490. [Google Scholar]
- Bremner, J. Organic nitrogen in soils. Soil Nitrogen 1965, 10, 93–149. [Google Scholar]
- Team, R. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015; Volume 42, p. 84. Available online: http://www.rstudio.com (accessed on 26 November 2022).
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 26 November 2022).
- González, I.; Déjean, S. Package CCA: Canonical Correlation Analysis. Available online: https://cran.r-project.org/web/packages/CCA/index.html (accessed on 26 November 2022).
- Lajuni, J.; Latiff, A. Biomass and floristic composition of Bangi Permanent Forest Reserve, a twice-logged lowland dipterocarp forest in Peninsular Malaysia. Sains Malays. 2013, 42, 1517–1521. [Google Scholar]
- Khairil, M.; Juliana, W.W.; Nizam, M. Edaphic influences on tree species composition and community structure in a tropical watershed forest in Peninsular Malaysia. J. Trop. For. Sci. 2014, 26, 284–294. [Google Scholar]
- Nizam, M.; Norziana, J.; Sahibin, A.; Latiff, A. Edaphic relationships among tree species in the National Park at Merapoh, Pahang, Malaysia. J. Biosci. 2006, 17, 37–53. [Google Scholar]
- Fujii, K. Soil acidification and adaptations of plants and microorganisms in Bornean tropical forests. Ecol. Res. 2014, 29, 371–381. [Google Scholar]
- McCauley, A.; Jones, C.; Jacobsen, J. Soil pH and organic matter. Nutr. Manag. Modul. 2009, 8, 1–12. [Google Scholar]
- Aprile, F.; Lorandi, R. Evaluation of cation exchange capacity (CEC) in tropical soils using four different analytical methods. J. Agric. Sci. 2012, 4, 278. [Google Scholar]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Heil, D.; Sposito, G. Chemical attributes and processes affecting soil quality. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 1997; Volume 25, pp. 59–79. [Google Scholar]
- Hartemink, A.E. Soil Analysis-An Interpretation Manual-KI Peverill, LA Sparrow & DJ Reuter (Eds.). CSIRO Publishing, Collingwood, Vic, 369 pp., 1999. Hardback. ISBN 0 643 06376 5 (A $94.95). Catena 2000, 39, 301–303. [Google Scholar]
- Kong, J.; He, Z.; Chen, L.; Yang, R.; Du, J.; Lin, P.; Zhu, X.; Tian, Q. Elevational gradients and distributions of aggregate associated organic carbon and nitrogen and stability in alpine forest ecosystems. Soil Sci. Soc. Am. J. 2020, 84, 1971–1982. [Google Scholar]
- Marrs, R.; Proctor, J.; Heaney, A.; Mountford, M. Changes in soil nitrogen-mineralization and nitrification along an altitudinal transect in tropical rain forest in Costa Rica. J. Ecol. 1988, 76, 466–482. [Google Scholar]
- Spain, A.V. Influence of environmental conditions and some soil chemical properties on the carbon and nitrogen contents of some tropical Australian rainforest soils. Soil Res. 1990, 28, 825–839. [Google Scholar]
- Alvarez-Clare, S.; Mack, M.C. Influence of precipitation on soil and foliar nutrients across nine Costa Rican forests. Biotropica 2011, 43, 433–441. [Google Scholar]
- Adnan, M.; Fahad, S.; Saleem, M.H.; Ali, B.; Mussart, M.; Ullah, R.; Arif, M.; Ahmad, M.; Shah, W.A.; Romman, M. Comparative efficacy of phosphorous supplements with phosphate solubilizing bacteria for optimizing wheat yield in calcareous soils. Sci. Rep. 2022, 12, 11997. [Google Scholar]
- Ali, B.; Hafeez, A.; Javed, M.A.; Afridi, M.S.; Abbasi, H.A.; Qayyum, A.; Batool, T.; Ullah, A.; Marc, R.A.; Al Jaouni, S.K. Role of endophytic bacteria in salinity stress amelioration by physiological and molecular mechanisms of defense: A comprehensive review. S. Afr. J. Bot. 2022, 151, 33–46. [Google Scholar]
- Ahmad, M.; Ishaq, M.; Shah, W.A.; Adnan, M.; Fahad, S.; Saleem, M.H.; Khan, F.U.; Mussarat, M.; Khan, S.; Ali, B. Managing phosphorus availability from organic and inorganic sources for optimum wheat production in calcareous soils. Sustainability 2022, 14, 7669. [Google Scholar]
- Schlesinger, W.H.; Bernhardt, E.S. Biogeochemistry: An Analysis of Global Change; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Havlin, J.L.; Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers; Pearson Education India: Chennai, India, 2016. [Google Scholar]
- Fahad, S.; Chavan, S.B.; Chichaghare, A.R.; Uthappa, A.R.; Kumar, M.; Kakade, V.; Pradhan, A.; Jinger, D.; Rawale, G.; Yadav, D.K. Agroforestry Systems for Soil Health Improvement and Maintenance. Sustainability 2022, 14, 14877. [Google Scholar]
- Terborgh, J. Diversity and the Tropical Rain Forest; Scientific American Library: Newyork, NY, USA, 1992. [Google Scholar]
- Afridi, M.; Javed, M.; Ali, S.; De Medeiros, F.; Ali, B.; Salam, A.; Sumaira; Marc, R.A.; Alkhalifah, D.H.M.; Selim, S.; et al. New opportunities in plant microbiome engineering for increasing agricultural sustainability under stressful conditions. Front. Plant Sci. 2022, 13, 899464. [Google Scholar]
- Banin, L.; Lewis, S.L.; Lopez-Gonzalez, G.; Baker, T.R.; Quesada, C.A.; Chao, K.J.; Burslem, D.F.; Nilus, R.; Abu Salim, K.; Keeling, H.C. Tropical forest wood production: A cross-continental comparison. J. Ecol. 2014, 102, 1025–1037. [Google Scholar]
- Ashton, P.S. Dipterocarp biology as a window to the understanding of tropical forest structure. Annu. Rev. Ecol. Syst. 1988, 19, 347–370. [Google Scholar]
- Numata, S.; Yasuda, M.; Suzuki, R.O.; Hosaka, T.; Noor, N.S.M.; Fletcher, C.D.; Hashim, M. Geographical pattern and environmental correlates of regional-scale general flowering in Peninsular Malaysia. PLoS ONE 2013, 8, e79095. [Google Scholar]
- Suzuki, E.; Ashton, P.S. Sepal and nut size ratio of fruits of Asian Dipterocarpaceae and its implications for dispersal. J. Trop. Ecol. 1996, 12, 853–870. [Google Scholar]
- Smith, J.R.; Bagchi, R.; Ellens, J.; Kettle, C.J.; Burslem, D.F.; Maycock, C.R.; Khoo, E.; Ghazoul, J. Predicting dispersal of auto-gyrating fruit in tropical trees: A case study from the D ipterocarpaceae. Ecol. Evol. 2015, 5, 1794–1801. [Google Scholar] [PubMed] [Green Version]
- Brearley, F.Q. Ectomycorrhizal associations of the Dipterocarpaceae. Biotropica 2012, 44, 637–648. [Google Scholar]
- Berrill, J.-P.; O’Hara, K.L. How do biophysical factors contribute to height and basal area development in a mixed multiaged coast redwood stand? Forestry 2016, 89, 170–181. [Google Scholar]
- Livesley, S.; Ossola, A.; Threlfall, C.; Hahs, A.; Williams, N. Soil carbon and carbon/nitrogen ratio change under tree canopy, tall grass, and turf grass areas of urban green space. J. Environ. Qual. 2016, 45, 215–223. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Number. | Family | Number of Genera | Number of Species | Number of Individuals |
|---|---|---|---|---|
| 1. | Achariaceae | 1 | 2 | 3 |
| 2. | Anacardiaceae | 1 | 2 | 15 |
| 3. | Arecaceae | 1 | 1 | 4 |
| 4. | Dipterocarpaceae | 3 | 10 | 26 |
| 5. | Fabaceae | 2 | 2 | 5 |
| 6. | Lauraceae | 1 | 1 | 4 |
| 7. | Meliaceae | 2 | 2 | 6 |
| 8. | Moraceae | 1 | 2 | 3 |
| 9. | Phyllanthaceae | 1 | 1 | 9 |
| 10. | Salicaceae | 1 | 3 | 10 |
| 11. | Sapindaceae | 1 | 1 | 8 |
| Total | 15 | 27 | 93 |
| DBH | Number of Trees |
|---|---|
| C-1: 05.00–14.90 cm | 6 |
| C-2: 15.00–24.90 cm | 13 |
| C-3: 25.00–34.90 cm | 20 |
| C-4: 35.00–44.90 cm | 15 |
| C-5: 45.00–54.90 cm | 12 |
| C-6: 55.00–64.90 cm | 8 |
| C-7: Above 65.00 cm | 19 |
| Family | Species | Number of Individuals | Relative Density (%) | Frequency (%) |
|---|---|---|---|---|
| Achariaceae | Hydnocarpus castanea Hook. f. and Thoms. | 2 | 2.2 | 17 |
| Achariaceae | Hydnocarpus scortechinii King. | 1 | 1.1 | 17 |
| Anacardiaceae | Mangifera indica L. | 1 | 1.1 | 17 |
| Anacardiaceae | Mangifera odorata Griff. | 14 | 15.1 | 33 |
| Apocynaceae | Alstonia scholaris (L.) R.Br. | 4 | 4.3 | 33 |
| Arecaceae | Ptychosperma macarthurii H.Wendl. ex H.J.Veitch) H.Wendl. ex Hook.f. | 4 | 4.3 | 33 |
| Dipterocarpaceae | Hopea helferi Brandis. | 3 | 3.2 | 17 |
| Dipterocarpaceae | Hopea nervosa King. | 2 | 2.2 | 17 |
| Dipterocarpaceae | Hopea nutans Ridl. | 4 | 4.3 | 17 |
| Dipterocarpaceae | Hopea sp. | 1 | 1.1 | 17 |
| Dipterocarpaceae | Shorea assamica subsp. globifera (Ridl.) Syme | 1 | 1.1 | 17 |
| Dipterocarpaceae | Shorea foxworthyi Symington. | 5 | 5.4 | 17 |
| Dipterocarpaceae | Shorea laevis Ridl. | 2 | 2.2 | 17 |
| Dipterocarpaceae | Shorea macrophylla (de Vriese) P.S.Ashton | 1 | 1.1 | 17 |
| Dipterocarpaceae | Shorea rubra P.S.Ashton | 3 | 3.2 | 17 |
| Fabaceae | Koompassia excelsa (Becc.) Taub. | 3 | 3.2 | 17 |
| Fabaceae | Pongamia pinnata (L.) Pierre | 2 | 2.2 | 17 |
| Lauraceae | Actinodaphne macrophylla (Blume) Nees | 4 | 4.3 | 17 |
| Meliaceae | Aglaia korthalsii Miq. Pellegr. | 3 | 3.2 | 17 |
| Meliaceae | Sandoricum koetjape (Burm.f.) Merr. | 3 | 3.2 | 17 |
| Moraceae | Ficus benjamina L. | 2 | 2.2 | 17 |
| Moraceae | Ficus microcarpa L.f. | 1 | 1.1 | 17 |
| Phyllanthaceae | Baccaurea motleyana (Müll.Arg.) Müll.Arg. | 9 | 9.7 | 17 |
| Salicaceae | Flacourtia indica (Burm.f.) Merr. | 5 | 5.4 | 17 |
| Salicaceae | Flacourtia inermis Roxb. | 1 | 1.1 | 17 |
| Salicaceae | Flacourtia jangomas (Lour.) Raeusch. | 4 | 4.3 | 17 |
| Sapindaceae | Pometia pinnata J.R.Forst. and G.Forst. | 8 | 8.6 | 33 |
| T | ||||
| Species | IVi (%) |
|---|---|
| Mangifera odorata Griff. | 68.8 |
| Pometia pinnata J.R.Forst. and G.Forst. | 46.84 |
| Alstonia scholaris (L.) R.Br. | 37.79 |
| Ptychosperma macarthurii H.Wendl. ex H.J.Veitch) H.Wendl. ex Hook.f. | 37.57 |
| Sandoricum koetjape (Burm.f.) Merr. | 35.01 |
| Baccaurea motleyana (Müll.Arg.) Müll.Arg. | 33.95 |
| Koompassia excelsa (Becc.) Taub. | 28.06 |
| Pongamia pinnata (L.) Pierre | 26.54 |
| Actinodaphne macrophylla (Blume) Nees | 25.67 |
| Shorea foxworthyi Symington | 25.45 |
| Plot | pH | CEC | Total (%) | Extractable (mg/kg) | |
|---|---|---|---|---|---|
| (cmol/kg) | C | N | P | ||
| Plot 1 | 4.89 | 4.80 | 0.94 | 0.07 | 1.92 |
| Plot 2 | 4.54 | 3.67 | 1.29 | 0.08 | 2.32 |
| Plot 3 | 4.55 | 3.62 | 0.85 | 0.06 | 2.30 |
| Plot 4 | 4.41 | 2.87 | 0.67 | 0.05 | 2.24 |
| Plot 5 | 4.67 | 5.35 | 0.81 | 0.06 | 2.29 |
| Plot 6 | 4.46 | 3.66 | 1.28 | 0.09 | 2.26 |
| Mean | 4.69 | 4.0 | 0.97 | 0.07 | 2.22 |
| Min | 4.41 | 2.87 | 0.81 | 0.05 | 1.92 |
| Max | 4.89 | 4.80 | 1.29 | 0.09 | 2.32 |
| Plot | Total Percentage (%) | C/N Ratio | |
|---|---|---|---|
| C | N | ||
| 1 | 0.94 | 0.07 | 13.43 |
| 2 | 1.29 | 0.08 | 16.13 |
| 3 | 0.85 | 0.06 | 14.17 |
| 4 | 0.67 | 0.05 | 13.40 |
| 5 | 0.81 | 0.06 | 13.5 |
| 6 | 1.28 | 0.09 | 14.22 |
| pH | CEC | C | N | |
|---|---|---|---|---|
| CEC | 0.8 | |||
| C | −0.087 | −0.057 | ||
| N | 0.005 | 0.041 | 0.956 ** | |
| P | −0.778 | −0.359 | 0.138 | −0.008 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isa, N.; Razak, S.A.; Abdullah, R.; Khan, M.N.; Hamzah, S.N.; Kaplan, A.; Dossou-Yovo, H.O.; Ali, B.; Razzaq, A.; Wahab, S.; et al. Relationship between the Floristic Composition and Soil Characteristics of a Tropical Rainforest (TRF). Forests 2023, 14, 306. https://doi.org/10.3390/f14020306
Isa N, Razak SA, Abdullah R, Khan MN, Hamzah SN, Kaplan A, Dossou-Yovo HO, Ali B, Razzaq A, Wahab S, et al. Relationship between the Floristic Composition and Soil Characteristics of a Tropical Rainforest (TRF). Forests. 2023; 14(2):306. https://doi.org/10.3390/f14020306
Chicago/Turabian StyleIsa, Nurrunnuha, Sarah Abdul Razak, Rosazlin Abdullah, Muhammad Nauman Khan, Siti Nasuha Hamzah, Alevcan Kaplan, Hubert Olivier Dossou-Yovo, Baber Ali, Abdul Razzaq, Sana Wahab, and et al. 2023. "Relationship between the Floristic Composition and Soil Characteristics of a Tropical Rainforest (TRF)" Forests 14, no. 2: 306. https://doi.org/10.3390/f14020306