
Infestation by Ips amitinus (Eichhoff, 1872), Its Associated Fungi, and Butt Rots in Stands of Pinus sibirica in South-Western Siberia
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Collecting Beetles
2.2. DNA Isolation, PCR Amplification, Sequencing, and Bioinformatics
2.3. Statistical Analyses
2.4. Isolation and Molecular Identification of Fungal Cultures
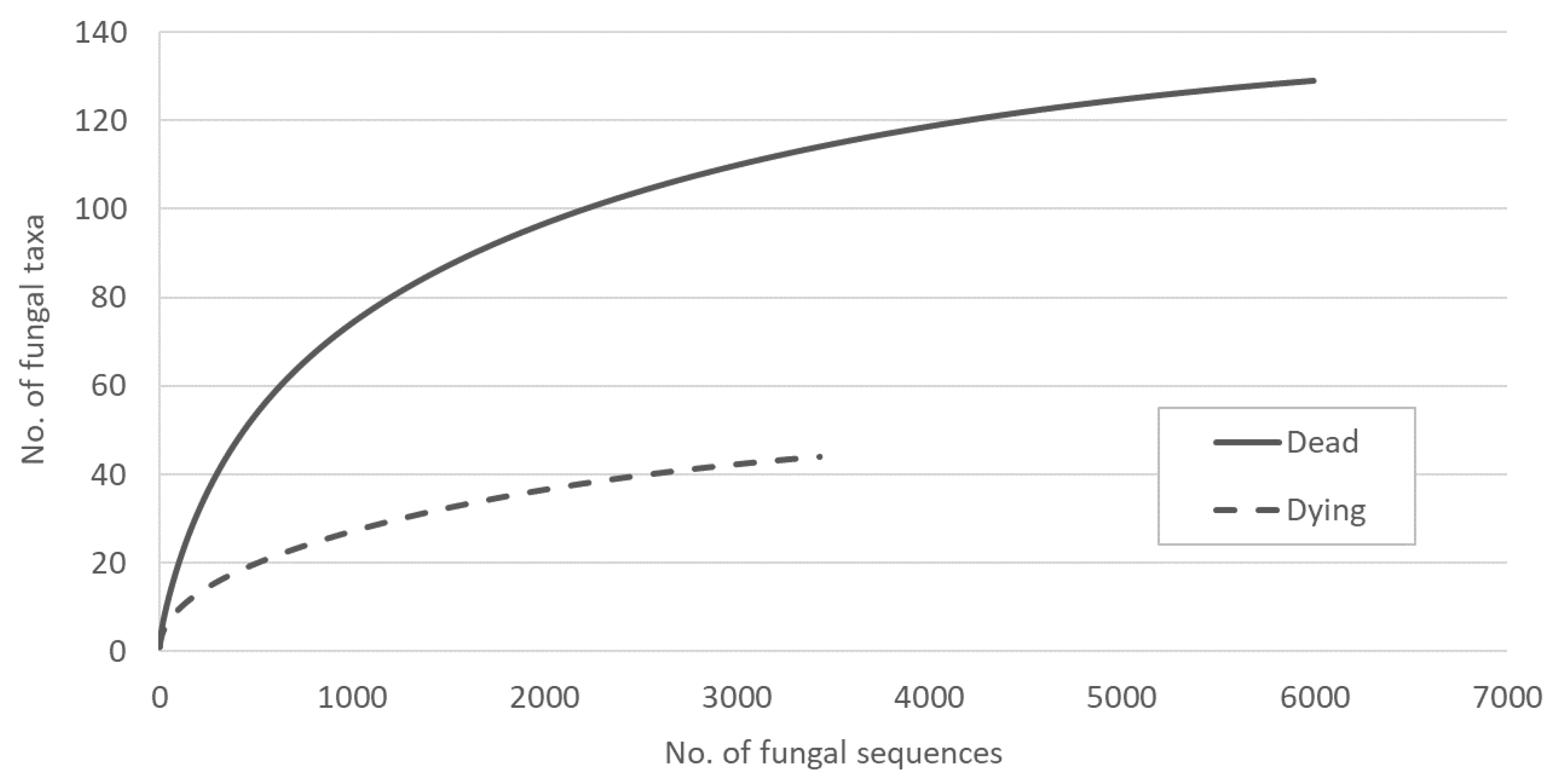
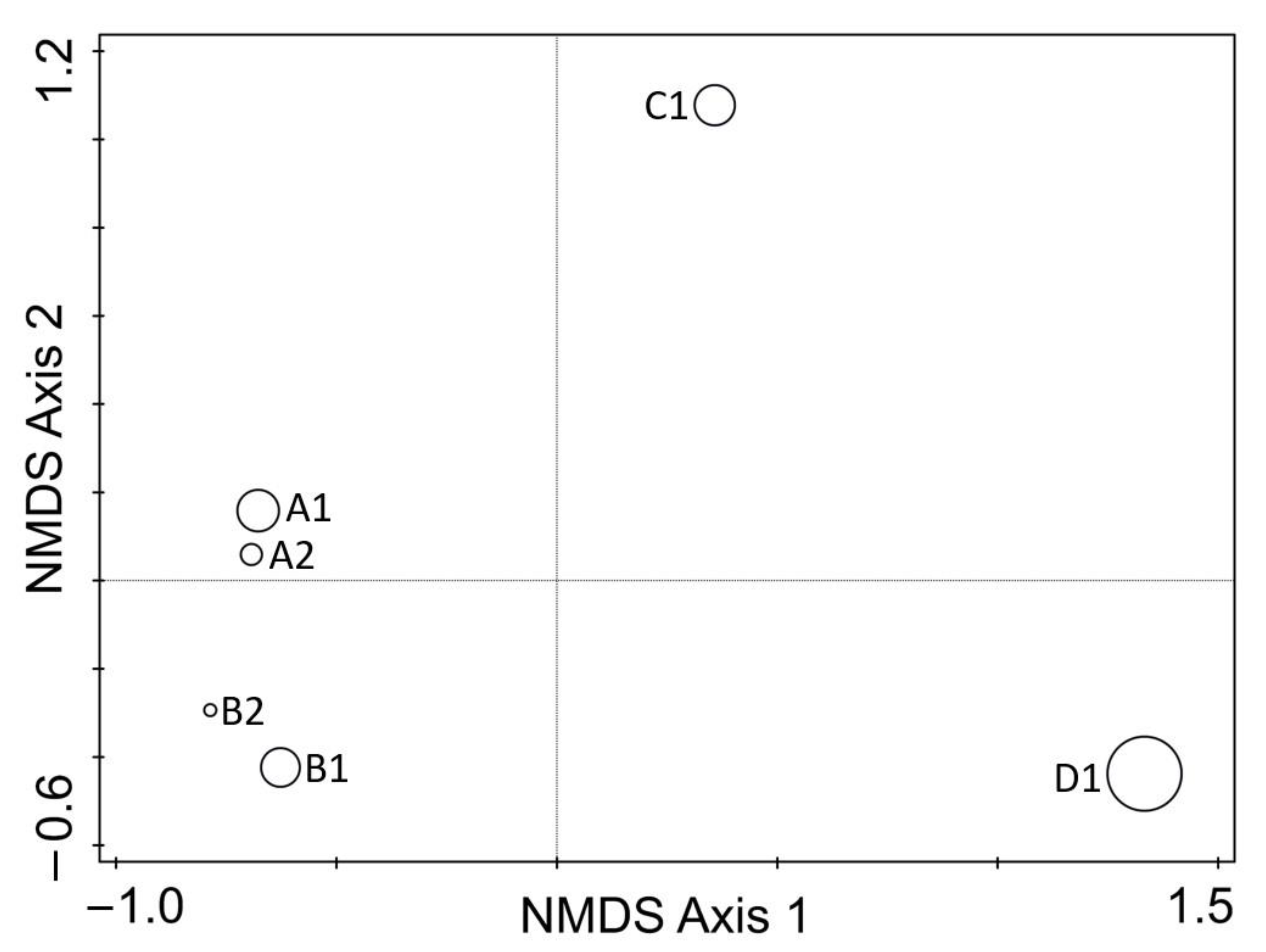
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knížek, M. Curculionidae: Scolytinae. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2011; Volume 7, Curculionidae I; pp. 86–88, 204–251. [Google Scholar]
- Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive insect pests of forests and urban trees in Russia: Origin, pathways, damage, and management. Forests 2022, 13, 521. [Google Scholar] [CrossRef]
- Pfeffer, A. Zentral- und Westpaläarktishe Borken- und Kernkäfer (Coleoptera: Scolytidae, Platypodidae). Entomol. Basiliensia 1994, 17, 5–310. [Google Scholar]
- Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Mandelshtam, M.Y.; Musolin, D.L.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agric. For. Entomol. 2019, 21, 286–298. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Krivets, S.A. An attack of Ips amitinus (Coleoptera, Curculionidae: Scolytinae) on arboretum in West Siberia: New host of invasive bark beetle among exotic conifers. J. Asia–Pac. Entomol. 2021, 24, 148–152. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Mandelshtam, M.Y.; Krivets, S.A.; Ilinsky, Y.Y. Small spruce bark beetle Ips amitinus (Eichhoff, 1872) (Coleoptera, Curculionidae: Scolytinae): A new alien species West Siberia. Entomol. Rev. 2019, 99, 592–599. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Krivets, S.A.; Bisirova, E.M.; Smirnov, N.A. Distribution of the small spruce bark beetle Ips amitinus (Eichhoff, 1872) (Coleoptera, Curculionidae: Scolytinae) in Western Siberia. Russ. J. Biol. Invasions 2022, 13, 77–84. [Google Scholar] [CrossRef]
- Krutovskii, K.V.; Bergmann, F. Introgressive hybridization and phylogenetic relationships between Norway, Picea abies (L.) Karst., and Siberian, P. obovata Ledeb., spruce species studied by isozyme loci. Heredity 1995, 74, 464–480. [Google Scholar] [CrossRef]
- Erbilgin, N.; Ma, C.; Whitehouse, C.; Shan, B.; Najar, A.; Evenden, M. Chemical similarity between historical and novel host plants promotes range and host expansion of the mountain pine beetle in a naive host ecosystem. New Phytol. 2014, 201, 940–950. [Google Scholar] [CrossRef]
- Rosenberger, D.W.; Venette, R.C.; Maddox, M.P.; Aukema, B.H. Colonization behaviors of mountain pine beetle on novel hosts: Implications for range expansion into northeastern North America. PLoS ONE 2017, 12, e0176269. [Google Scholar] [CrossRef]
- Bekh, I.A.; Danchenko, A.M.; Kibish, I.V. Sosna Kedrovaya Sibirskaya: Sibirskoe Chudo-derevo (Siberian Stone Pine: A Siberian Wonder Tree); Tomsk Gosudarstvennyi Universitet: Tomsk, Russia, 2004. (In Russian) [Google Scholar]
- Kirisits, T. Fungal associates of European bark beetles with special emphasis on the Ophiostomatoid fungi. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H.F., Eds.; Kluwer: Dordrecht, The Netherlands, 2004; pp. 181–236. [Google Scholar]
- Jankowiak, R.; Kacprzyk, M.; Mlynarczik, M. Diversity of ophiostomatoid fungi associated with bark beetles (Coleoptera: Scolytidae) colonizing branches of Norway spruce (Picea abies) in southern Poland. Biologia 2009, 64, 1170–1177. [Google Scholar] [CrossRef]
- Repe, A.; Kirisits, T.; Piskur, B.; de Groot, M.; Kump, B.; Jurc, M. Ophiostomatoid fungi associated with three spruce-infesting bark beetles in Slovenia. Ann. For. Sci. 2013, 70, 717–727. [Google Scholar] [CrossRef]
- Lukasova, K.; Holusa, J.; Turcani, M. Pathogens of Ips amitinus: New species and comparison with Ips typographus. J. Appl. Entomol. 2013, 137, 188–196. [Google Scholar] [CrossRef]
- Wegensteiner, R.; Stradner, A.; Handel, U. Occurrence of pathogens in Ips typographus (Coleoptera: Curculionidae) and in other spruce bark beetles from the wilderness reserve Durrenstein (Lower Austria). Biologia 2014, 69, 92–100. [Google Scholar] [CrossRef]
- Holusa, J.; Lukasova, K.; Zizka, Z.; Handel, U.; Haidler, B.; Wegensteiner, R. Occurrence of Microsporidium sp. and other pathogens in Ips amitinus (Coleoptera: Curculionidae). Acta Patasitol. 2016, 61, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Hyblerova, S.; Medo, J.; Barta, M. Diversity and prevalence of entomopathogenic fungi (Ascomycota, Hypocreales) in epidemic populations of bark beetles (Coleoptera, Scolitynae) in spruce forests of the Tatra National Park in Slovakia. Ann. For. Res. 2021, 64, 129–145. [Google Scholar] [CrossRef]
- Kernaghan, G.; Sigler, L.; Khasa, D. Mycorrhizal and root endophytic fungi of containerized Picea glauca seedlings assessed by rDNA sequence analysis. Microb. Ecol. 2003, 45, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.W.; Lindgren, S. Relationship between spruce beetle and tomentosus root disease: Two natural disturbance agents of spruce. Can. J. For. Res. 2002, 32, 31–37. [Google Scholar] [CrossRef]
- Kulhavy, D.L.; Partridge, A.D.; Stark, R.W. Root diseases and blister rust associated with bark beetles (Coleoptera: Scolytidae) in Western White pine in Idaho. Environ. Entomol. 1984, 13, 813–817. [Google Scholar] [CrossRef]
- Jankovsky, L.; Cudlin, P.; Moravec, I. Root decays as a potential predisposition factor of a bark beetle disaster in the Sumava Mts. J. For. Sci. 2003, 49, 125–132. [Google Scholar] [CrossRef]
- Christiansen, E.; Huse, K.J. Infestation ability of Ips typographus L. in Norway spruce in relation to butt rot, tree vitality and increment. Nor. Skogforsk. 1980, 35, 473–482. [Google Scholar]
- Pavlov, I.N. Biotic and abiotic factors as causes of coniferous forests dieback in Siberia and Far East. Contemp. Probl. Ecol. 2015, 8, 440–456. [Google Scholar] [CrossRef]
- Arefyev, S.P. Xylotrophic fungi—The causal agents of Siberian pine (Pinus sibirica du Tour) rot in the central taiga Irtish river basin. Mikol. I Fitopatol. 1991, 25, 419–425. (In Russian) [Google Scholar]
- Menkis, A.; Östbrant, I.-L.; Davydenko, K.; Bakys, R.; Balalaikins, M.; Vasaitis, R. Scolytus multistriatus associated with Dutch elm disease on the island of Gotland: Phenology and communities of vectored fungi. Mycol. Prog. 2016, 15, 55. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylo-genetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Fowler, J.; Cohen, L.; Jarvis, P. Practical Statistics for Field Biology; Wiley: Chichester, UK, 2001. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination, Version 4; Microcomputer Power: Ithaca, NY, USA, 1998. [Google Scholar]
- Menkis, A.; Vasaitis, R.; Taylor, A.F.S.; Stenström, E.; Stenlid, J.; Finlay, R. Fungi in decayed roots of conifer seedlings in forest nurseries, afforested clear-cuts and abandoned farmland. Plant Pathol. 2006, 55, 117–129. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Lou, Q.-Z.; Lu, M.; Sun, J.-H. Yeast diversity associated with invasive Dendroctonus valens killing Pinus tabuliformis in China using culturing and molecular methods. Microb. Ecol. 2014, 68, 397–415. [Google Scholar] [CrossRef]
- Mercado, J.E.; Ortiz-Santana, B.; Kay, S.L. Endemic Jeffrey pine beetle associates: Beetle/mite fungal dissemination strategies and interactions that may influence beetle population levels. Microorganisms 2021, 9, 1641. [Google Scholar] [CrossRef]
- Hausner, G.; Iranpour, M.; Kim, J.-J.; Breuil, C.; Davis, C.N.; Gibb, E.A.; Reid, J.; Loewen, P.C.; Hopkin, A.A. Fungi vectored by the introduced bark beetle Tomicus piniperda in Ontario, Canada, and comments on the taxonomy of Leptographium lundbergii, Leptographium terebrantis, Leptographium truncatum, and Leptographium wingfieldii. Can. J. Bot. 2005, 83, 10. [Google Scholar] [CrossRef]
- Chakraborty, A.; Modlinger, R.; Ashraf, M.Z.; Synek, J.; Schlyter, F.; Roy, A. Core mycobiome and their ecological relevance in the gut of five Ips bark beetles (Coleoptera: Curculionidae: Scolytinae). Front. Microbiol. 2020, 11, 568853. [Google Scholar] [CrossRef]
- Kluting, K.; Strid, Y.; Six, D.; Rosling, A. Forest fire influence on Tomicus piniperda—associated fungal communities and phloem nutrient availability of colonized Pinus sylvestris. Microb. Ecol. 2023, 86, 224–239. [Google Scholar] [CrossRef]
- Weiser, J.; Wegensteiner, R.; Händel, U.; Zizka, Z. Infections with the Ascomycete fungus Metschnikowia typographi sp.nov. in the bark beetles Ips typographus and Ips amitinus (Coleoptera, Scolitydae). Folia Microbiol. 2003, 48, 611–618. [Google Scholar] [CrossRef]
- Woodward, S.; Stenlid, J.; Karjalainen, R.; Huttermann, A. (Eds.) Heterobasidion Annosum, Biology, Ecology, Impact and Control; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Gonthier, P.; Thor, M. Annosus root and butt rots. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: Wallingford, UK, 2013; pp. 128–158. [Google Scholar]
- Dai, Y.C.; Cui, B.K.; Yuan, H.S.; Li, B.D. Pathogenic wood-decaying fungi in China. For. Pathol. 2007, 37, 105–120. [Google Scholar] [CrossRef]
- Tomsovsky, M.; Sedlak, P.; Jankovsky, L. Species recognition and phylogenetic relationships of European Porodaedalea (Basidiomycota, Hymenochaetales). Mycol. Prog. 2010, 9, 225–233. [Google Scholar] [CrossRef]
- Nitare, J. Indicator Species for Assessing the Nature Conservation Value of Woodland Sites; Skogsstyrelsen: Jönköping, Sweden, 2000. (In Swedish) [Google Scholar]
- Lannenpaa, A.; Aakala, T.; Kauhanen, H.; Kuuluvainen, T. Tree mortality agents in pristine Norway spruce forests in northern Fennoscandia. Silva Fenn. 2008, 42, 151–163. [Google Scholar] [CrossRef]
- Shaw, C.G.; Kile, G.A. (Eds.) Armillaria Root Disease; Agriculture Handbook No. 691; USDA Forest Service: Washington, DC, USA, 1991. [Google Scholar]
- Guillaumin, J.-J.; Legrand, P. Armillaria root rots. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: Wallingford, UK, 2013; pp. 159–177. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site/Tree | Tree/Branch Condition | Geographic Position | Sequences (No.) | Taxa (No.) | Shannon Diversity Index |
| A1 | dead | 56.3099, 85.1733 | 1445 | 46 | 1.6 |
| A2 | dying | “–” | 1657 | 32 | 1.3 |
| B1 | dead | 56.3102, 85.1362 | 1466 | 44 | 1.3 |
| B2 | dying | “–” | 1772 | 26 | 1.3 |
| C1 | dead | 56.5389, 84.7366 | 1483 | 45 | 1.6 |
| D1 | dead | 56.3855, 85.2599 | 1599 | 68 | 2.7 |
| All | 9422 | 143 |
| Site/Tree | A1 | B1 | C1 | D1 | A2 | B2 |
|---|---|---|---|---|---|---|
| A1 | - | 0.48 | 0.40 | 0.20 | 0.51 | 0.41 |
| B1 | - | 0.35 | 0.27 | 0.43 | 0.40 | |
| C1 | - | 0.36 | 0.38 | 0.27 | ||
| D1 | - | 0.21 | 0.20 | |||
| A2 | - | 0.45 | ||||
| B2 | - |
| Taxon | Sequence Reference | Similarity | GenBank Source | Abundance of Sequences, % | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| This Study | GenBank | bp | % | Description | A1 | B1 | C1 | D1 | A2 | B2 | All | |
| Bark-beetle-associated yeasts | ||||||||||||
| Nakazawaea holstii | OM614738 | KY104368 | 286/286 | 100 | Frass on Picea rubens, Quebec (CBS:2091) a | 57.2 | 61.6 | 58.2 | 38.4 | 70.6 | 66.9 | 59.0 |
| Kuraishia molischiana | OM614742 | KY103921 | 274/276 | 99 | Rotten wood of Fagus sylvatica, Hungary a, b | 17.4 | 22.6 | 6.5 | 5.2 | 17.2 | 9.9 | 13.0 |
| Nakazawaea ambrosiae | OM614743 | NR_165549 | 295/296 | 99 | Ips typographus gallery, Picea abies, Moscow | 13.0 | 7.8 | 18.5 | 7.5 | 4.6 | 9.8 | 10.1 |
| Ogataea pini | OM614767 | HQ413282 | 280/289 | 99 | Dendroctonus brevicomis, Pinus ponderosa, Arizona | 0.6 | 0.2 | 0.5 | 0.6 | 1.1 | 0.3 | 0.6 |
| Ogataea neixiangensis | OM614788 | NR_173264 | 275/280 | 98 | Rotten wood, central China | 0.2 | 0.6 | - | - | 0.1 | 0.2 | 0.2 |
| Cryptococcus sp. 58 | OM614753 | MH697750 | 280/280 | 100 | Dendroctonus micans gallery, Picea sp., Moscow | 0.1 | - | 0.1 | - | 0.1 | - | 0.1 |
| Myxozyma melibiosi | OM614830 | KY037840 | 243/245 | 99 | Dendroctonus monticolae, Pinus ponderosa, California | 0.2 | 0.1 | - | - | - | 0.2 | 0.1 |
| All yeasts | 88.7 | 92.9 | 83.8 | 51.7 | 93.6 | 87.3 | 92.9 | |||||
| Ophiostomatoid fungi | ||||||||||||
| Ophiostoma bicolor | OM614758 | HE866712 | 315/316 | 99 | Pityogenes chalcographus, Picea abies, Slovenia | 0.3 | 1.0 | 0.1 | - | 0.8 | 5.2 | 1.3 |
| Ophiostoma sp. 1 | OM614762 | MW460002 | 326/329 | 99 | Dendroctonus armandii, Pinus armandii, China | 1.2 | 0.3 | 2.2 | 0.8 | 0.5 | 1.1 | 1.0 |
| Ophiostoma hongxingense | OM614777 | MK748194 | 321/323 | 99 | Ips subelongatus, Larix gmelinii, northeast China | 0.3 | 0.7 | 0.5 | - | 0.1 | 0.6 | 0.4 |
| Leptographium truncatum | OM614847 | AB907609 | 295/297 | 99 | Pinus densiflora snag, Japan c | 0.2 | - | - | - | - | - | 0.03 |
| All ophiostomatoid fungi | 2.0 | 2.0 | 2.8 | 0.8 | 1.4 | 6.9 | 2.0 | |||||
| Some other identified wood- and bark-inhabiting fungi (lichens) | ||||||||||||
| Lophium arboricola | OM614739 | MK159395 | 250/251 | 99 | Abies koreana, Republic of Korea | - | 0.1 | 1.7 | - | - | - | 0.3 |
| Hypoxylon serpens | OM614784 | MG098299 | 258/258 | 100 | Twig of Pinus sylvestris, Germany | 1.3 | - | - | - | - | - | 0.2 |
| Orbilia vinosa | OM614792 | KT215265 | 286/289 | 99 | Branch of Pinus sylvestris, France | - | - | 0.3 | 0.6 | - | - | 0.2 |
| Claussenomyces sp. 22 | OM614746 | KY633581 | 239/242 | 99 | Picea rubens, east Canada | 0.2 | - | 0.3 | - | - | - | 0.1 |
| Stereum sanguinolentum | OM614831 | KF996533 | 293/294 | 99 | Wood of Picea abies, Italy | - | - | - | - | 0.3 | - | 0.1 |
| (Parmelia sulcata) | OM614750 | MN387112 | 244/244 | 100 | Lichen, Poland | - | - | 0.1 | 1.3 | - | - | 0.2 |
| (Cyphelium lucidum) | OM614798 | EF551165 | 251/251 | 100 | Lichen, Sweden | - | - | - | 0.8 | - | - | 0.1 |
| Sydowia polyspora | OM614744 | MK762617 | 256/256 | 100 | Pine, Tennessee, USA | - | 0.07 | - | - | 0.06 | - | 0.02 |
| Sarea resinae | OM614747 | MN720382 | 249/249 | 100 | Bark of Larix kaempferi, Japan | - | - | 0.13 | - | - | - | 0.02 |
| Celosporium laricicola | OM614779 | FJ997287 | 249/253 | 98 | Twigs of Larix lyallii, Alberta, Canada | - | - | 0.13 | - | - | - | 0.02 |
| All other identified wood fungi | 1.5 | 0.1 | 2.4 | 2.7 | 0.3 | - | 1.2 | |||||
| Total no. of generated sequences | 1445 | 1466 | 1483 | 1599 | 1657 | 1772 | 9422 | |||||
| Sequences of taxa presented in the table, no. | 1405 | 1393 | 1320 | 883 | 1579 | 1669 | 8249 | |||||
| % | 97.2 | 95.0 | 89.0 | 55.2 | 95.3 | 94.2 | 87.5 | |||||
| All detected taxa, no. | 46 | 44 | 45 | 68 | 32 | 26 | 143 | |||||
| Taxa presented in the table, no. | 15 | 11 | 14 | 8 | 11 | 9 | 23 | |||||
| % | 32.6 | 25.0 | 31.1 | 11.8 | 34.4 | 34.6 | 16.1 | |||||
| Fungal Species Sensu Lato | GenBank | Number of Beetles Harbouring Respective Fungus | ||||||
|---|---|---|---|---|---|---|---|---|
| Access. No. | In Branches/Trees a | |||||||
| Dead | Dying | All | ||||||
| A1 | B1 | C1 | D1 | A2 | B2 | (%) | ||
| Beauveria bassiana | OR584254 | 5 | - | - | 1 | 9 | - | 15 (25) |
| Lophium arboricola | OQ536234 | - | 1 | 2 | - | - | 1 | 4 (7) |
| Ophiostoma canum | OR574404 | - | 1 | - | 2 | - | 1 | 4 (7) |
| Ophiostoma bicolor | OR592582 | - | - | - | - | 1 | 2 | 3 (5) |
| Ophiostoma hongxingense | OR588874 | - | 1 | 1 | - | - | - | 2 (3) |
| Ophiostoma sp. 2 | OR589101 | 2 | - | - | - | - | - | 2 (3) |
| Chaetomium crispatum | OR584257 | - | - | - | 1 | - | - | 1 (2) |
| Fusarium avenaceum | 1 | - | - | - | - | - | 1 (2) | |
| Trichocladium griseum | OR589102 | - | - | - | 1 | - | - | 1 (2) |
| All fungi | 8 | 3 | 3 | 5 | 10 | 4 | 33 (55) | |
| Number of beetles subjected to isolation | 10 | 10 | 10 | 10 | 10 | 10 | 60 | |
| Fungus | GenBank | No. of Stems (%) | Of Those, at Height | |
|---|---|---|---|---|
| Access. No. | 0.2 m | 1.2 m | ||
| Heterobasidion parviporum | OR574397 | 12 (46) | 12 (46) | - |
| Phaeolus schweinitzii | OR512100 | 8 (31) | 8 (31) | - |
| Fomitopsis pinicola | OR512102 | 4 (15) | 2 (8) | 2 (8) |
| Porodaedalea pini | OR574401 | 3 (12) | 1 (4) | 2 (8) |
| Armillaria borealis | OR575851 | 2 (8) | 2 (8) | - |
| No. of all isolates a | 29 | 25 (96) | 4 (15) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlov, I.N.; Vasaitis, R.; Litovka, Y.A.; Timofeev, A.A.; Menkis, A. Infestation by Ips amitinus (Eichhoff, 1872), Its Associated Fungi, and Butt Rots in Stands of Pinus sibirica in South-Western Siberia. Forests 2023, 14, 2383. https://doi.org/10.3390/f14122383
Pavlov IN, Vasaitis R, Litovka YA, Timofeev AA, Menkis A. Infestation by Ips amitinus (Eichhoff, 1872), Its Associated Fungi, and Butt Rots in Stands of Pinus sibirica in South-Western Siberia. Forests. 2023; 14(12):2383. https://doi.org/10.3390/f14122383
Chicago/Turabian StylePavlov, Igor N., Rimvydas Vasaitis, Yulia A. Litovka, Anton A. Timofeev, and Audrius Menkis. 2023. "Infestation by Ips amitinus (Eichhoff, 1872), Its Associated Fungi, and Butt Rots in Stands of Pinus sibirica in South-Western Siberia" Forests 14, no. 12: 2383. https://doi.org/10.3390/f14122383