
Grazing Impacts on Soil Enzyme Activities Vary with Vegetation Types in the Forest-Steppe Ecotone of Northeastern China
Abstract
:1. Introduction
2. Materials and Methods
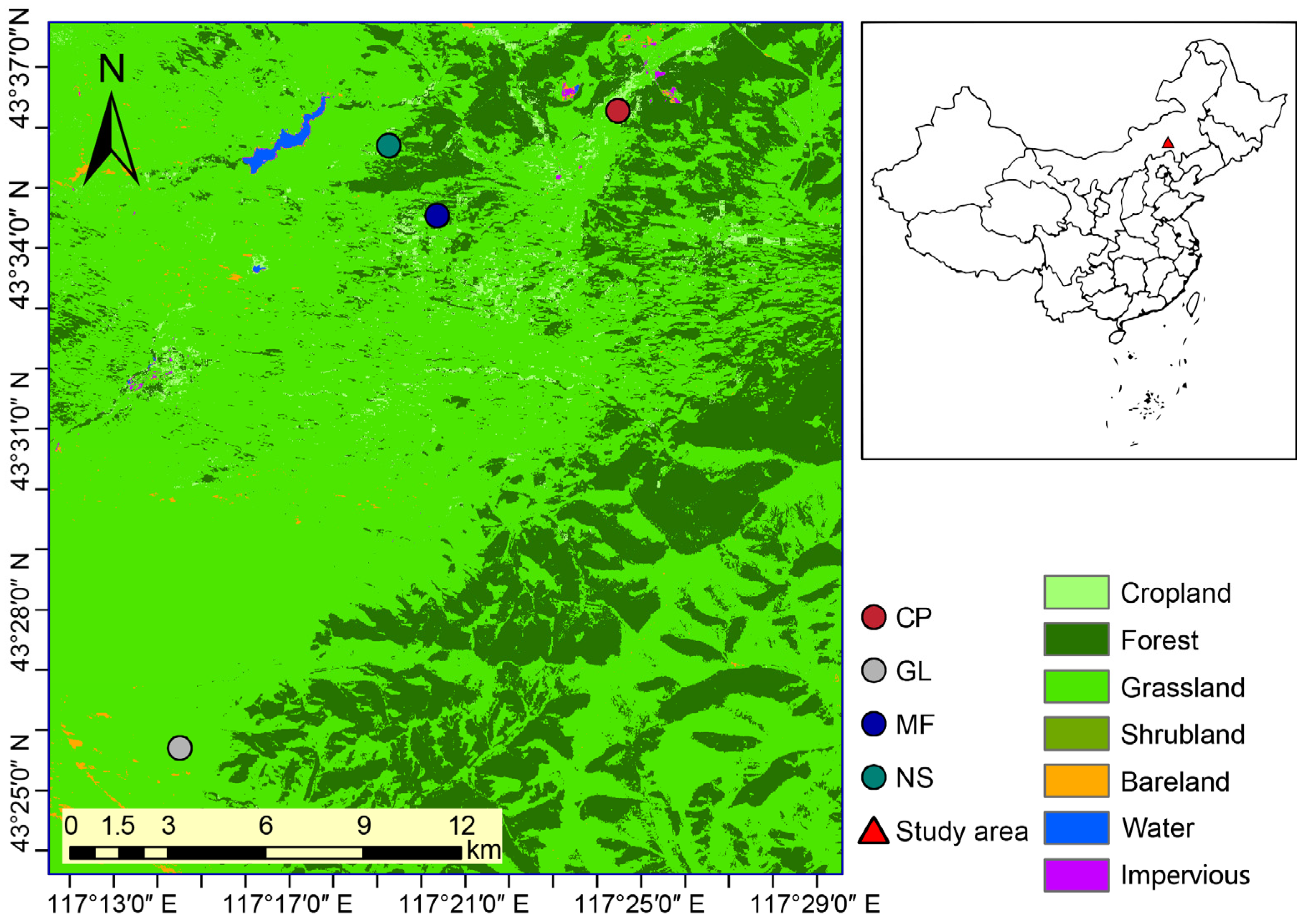
2.1. Study Area
2.2. Soil Sampling
2.3. Laboratory Analyses
2.4. Statistical Analysis
3. Results
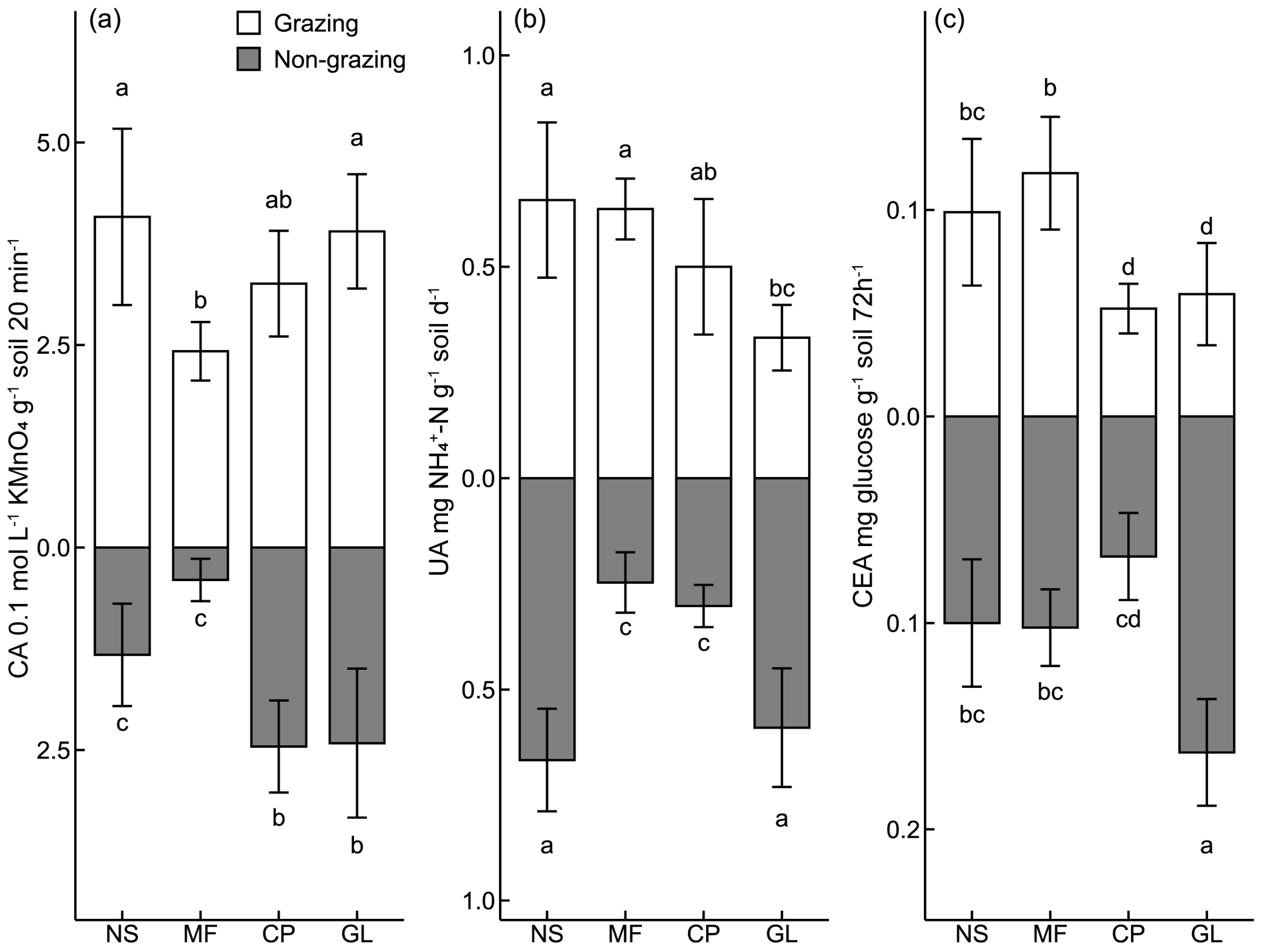
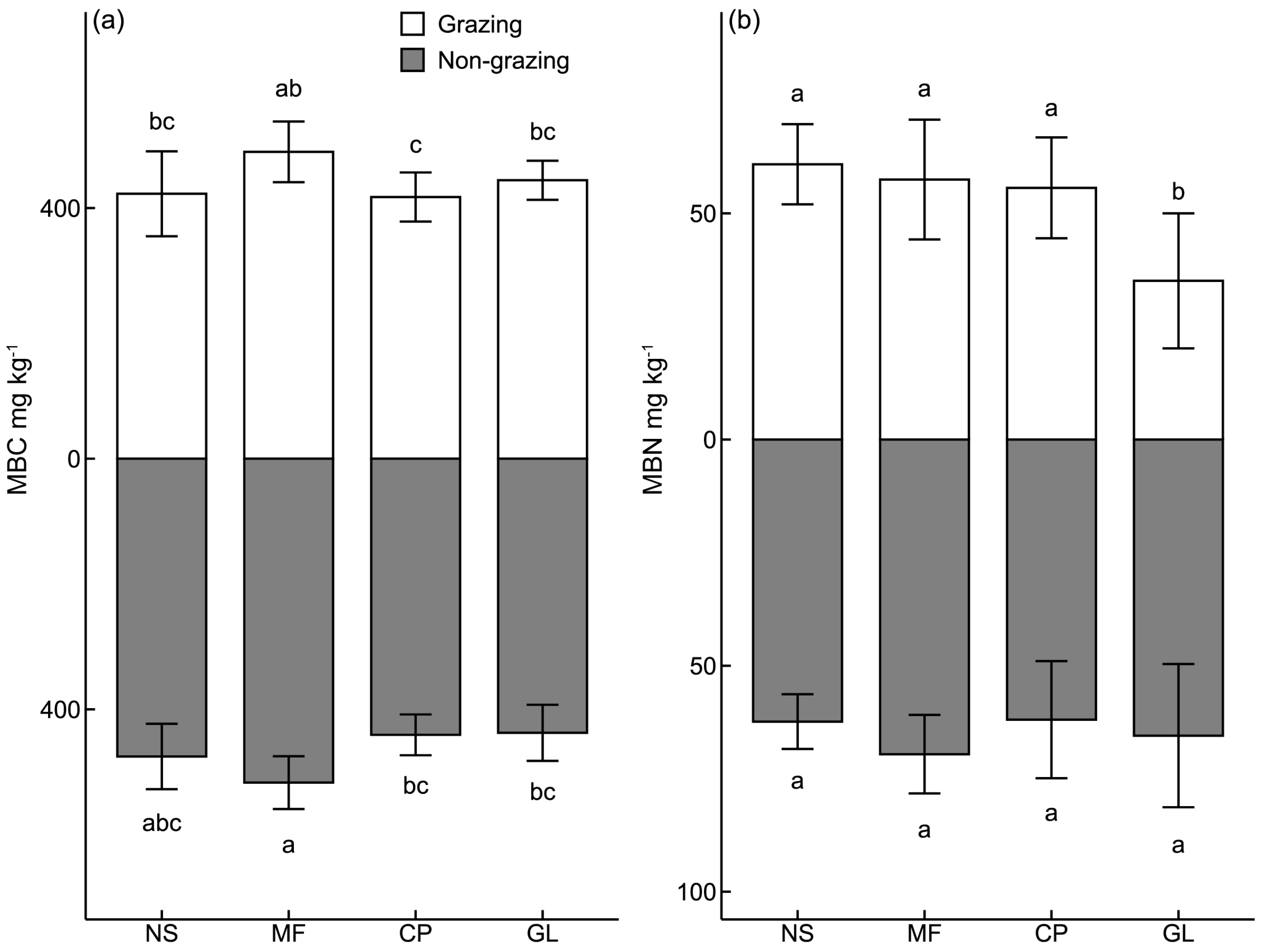
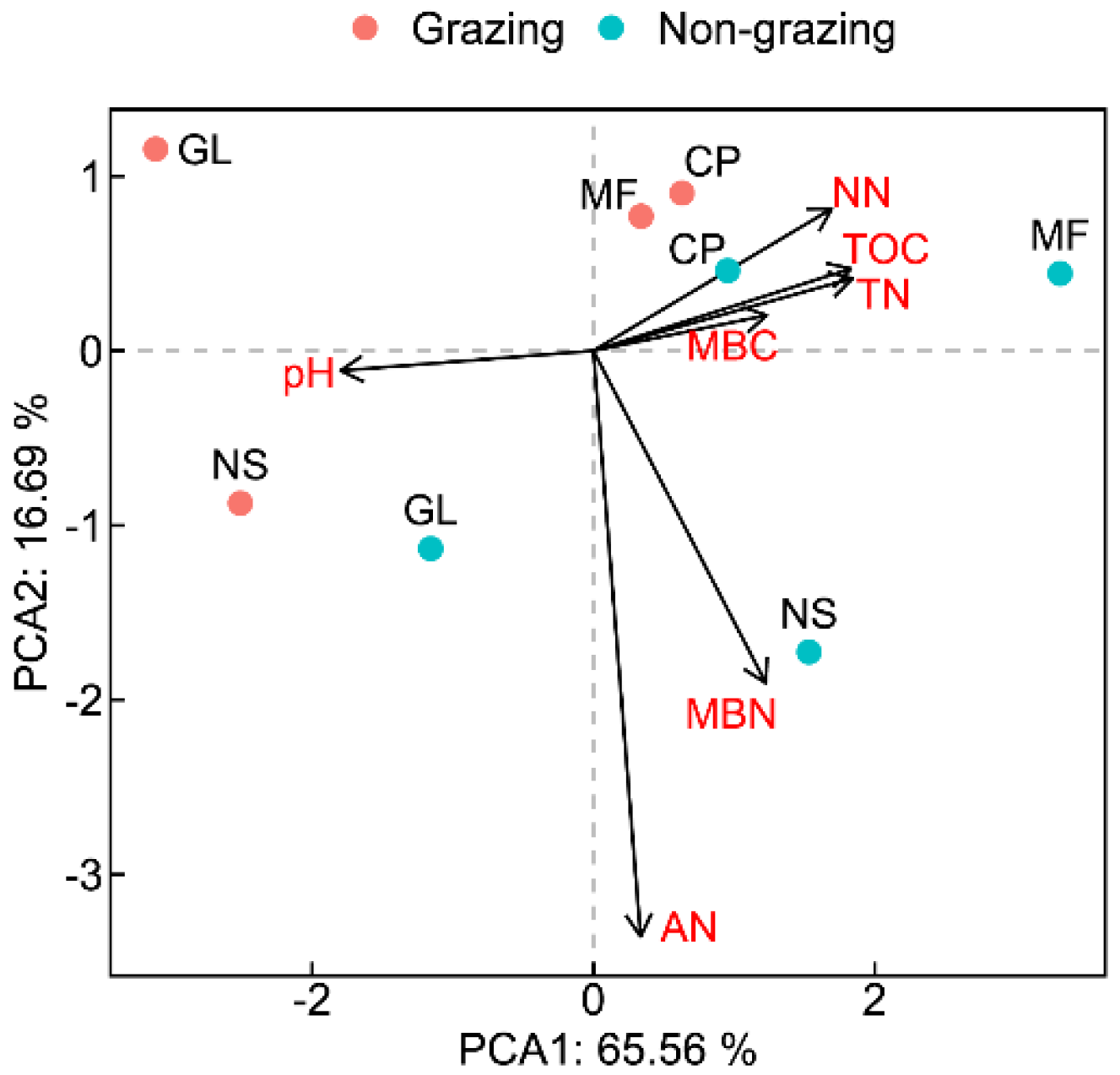
3.1. Characteristics of Soil Enzyme Activities and Microbial Biomass
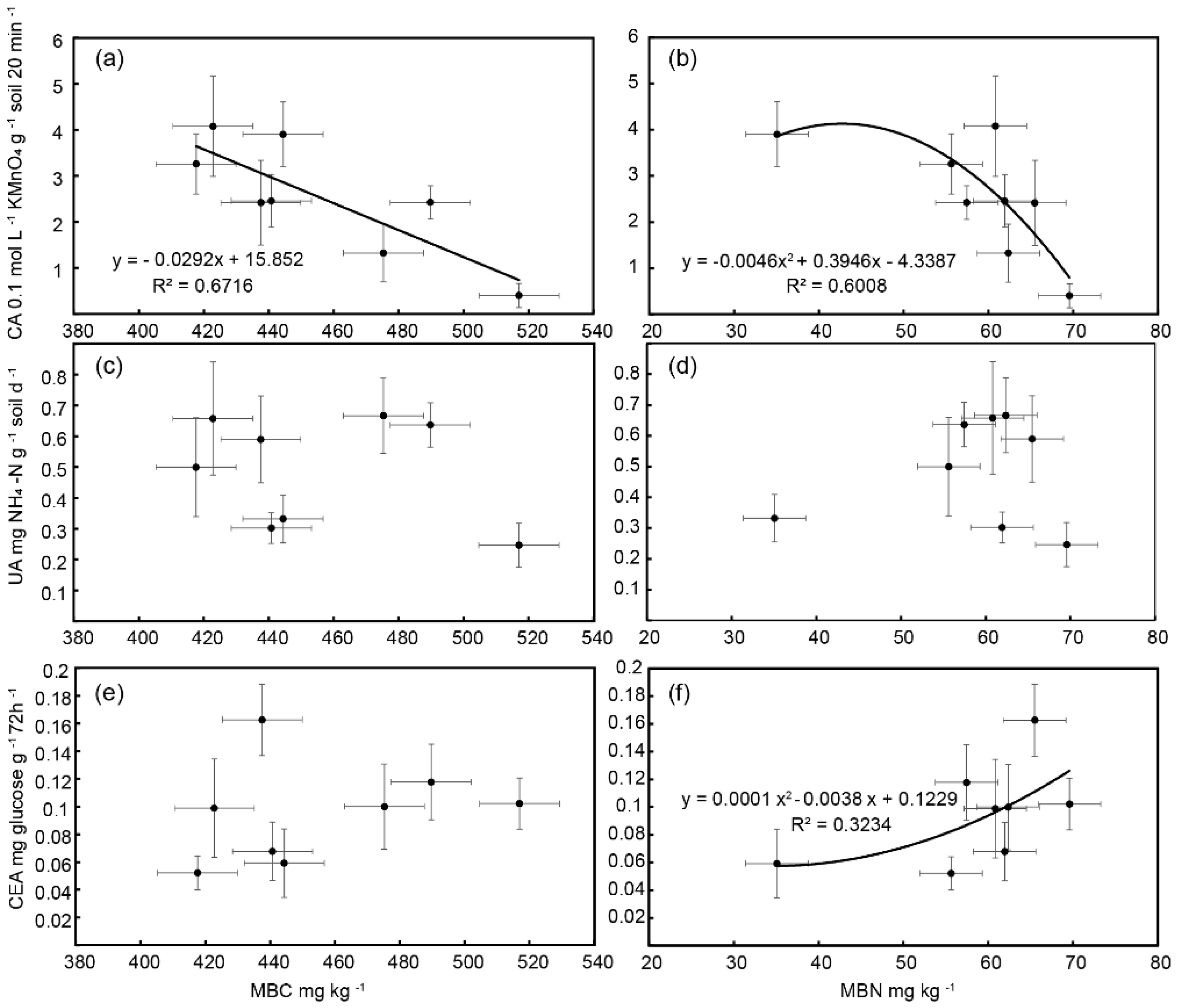
3.2. The Associations between Soil Enzyme Activities and Microbial Biomass
3.3. Differences and Commonalities in the Soil Environment
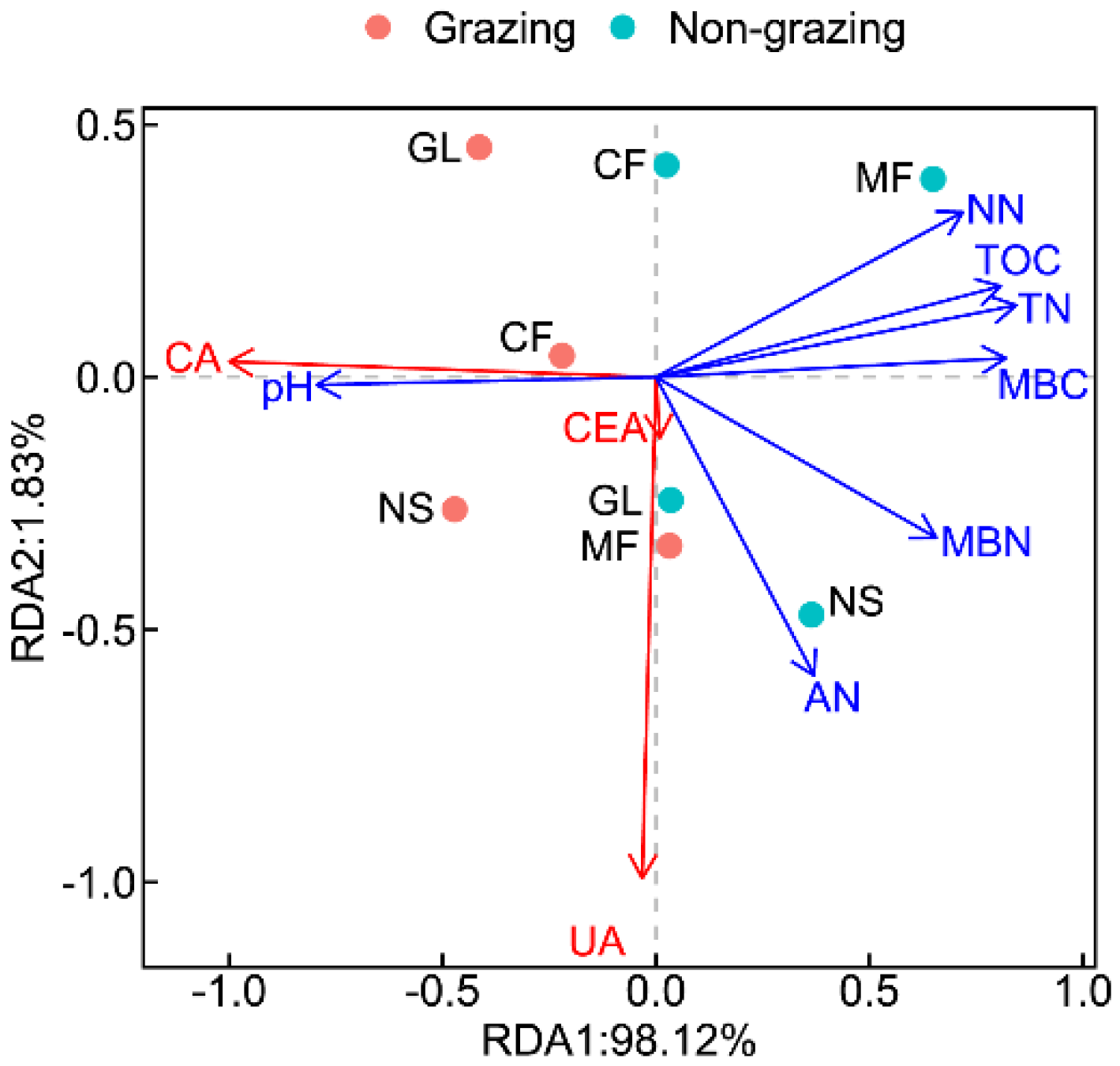
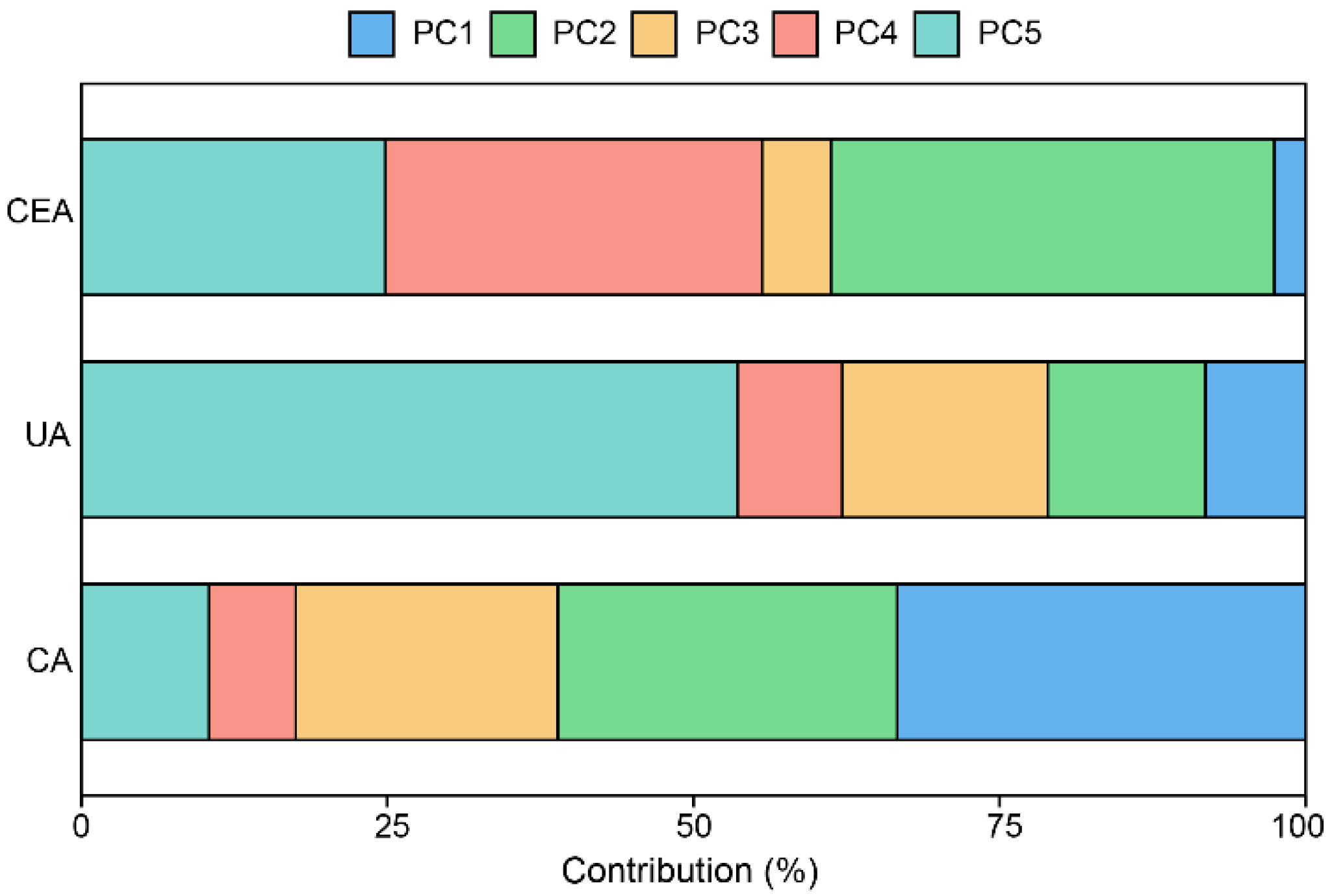
3.4. The Impact of Environmental Factors on Enzyme Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CA | catalase activities |
| UA | urease activities |
| CEA | cellulase activities |
| NS | natural secondary forests |
| MF | mixed forests |
| CP | coniferous plantations |
| GL | grasslands |
| MBC | microbial biomass carbon |
| MBN | microbial biomass nitrogen |
| TOC | total organic carbon |
| TN | total nitrogen |
| NN | nitrate nitrogen |
| AN | ammonium nitrogen |
References
- Feyissa, A.; Gurmesa, G.A.; Yang, F.; Long, C.; Zhang, Q.; Cheng, X. Soil enzyme activity and stoichiometry in secondary grasslands along a climatic gradient of subtropical China. Sci. Total Environ. 2022, 825, 154019. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Liu, C.; Song, Y.; Dong, X.; Wang, X.; Ma, X.; Zhao, G.; Zang, S. Soil enzyme activities and their relationships with soil C, N, and P in peatlands from different types of permafrost regions, Northeast China. Front. Environ. Sci. 2021, 9, 143. [Google Scholar] [CrossRef]
- Luo, R.; Fan, J.; Wang, W.; Luo, J.; Kuzyakov, Y.; He, J.; Chu, H.; Ding, W. Nitrogen and phosphorus enrichment accelerates soil organic carbon loss in alpine grassland on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2019, 650, 303–312. [Google Scholar] [CrossRef]
- Wang, H.; Wu, J.; Li, G.; Yan, L. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Agrawal, K.; Verma, P. Current perspective on production and applications of microbial cellulases: A review. Bioresour. Bioprocess. 2021, 8, 95. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Liu, G.; Xue, S. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China. Catena 2021, 196, 104812. [Google Scholar] [CrossRef]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China. Soil Tillage Res. 2019, 195, 104355. [Google Scholar] [CrossRef]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil enzyme activities as biological indicators of soil health. Rev. Environ. Health 2003, 18, 65. [Google Scholar] [CrossRef]
- Carrara, J.E.; Walter, C.A.; Hawkins, J.S.; Peterjohn, W.T.; Averill, C.; Brzostek, E.R. Interactions among plants, bacteria, and fungi reduce extracellular enzyme activities under long-term N fertilization. Glob. Chang. Biol. 2018, 24, 2721–2734. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, C.; Xu, X.; Wang, C.; Liu, G.; Liang, C.; Zuo, X.; Wang, C.; Lv, Y.; Wang, R. Different facets of bacterial and fungal communities drive soil multifunctionality in grasslands spanning a 3500 km transect. Funct. Ecol. 2022, 36, 3120–3133. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Ren, C.; Kang, D.; Wu, J.P.; Zhao, F.; Yang, G.; Han, X.; Feng, Y.; Ren, G. Temporal variation in soil enzyme activities after afforestation in the Loess Plateau, China. Geoderma 2016, 282, 103–111. [Google Scholar] [CrossRef]
- Jourgholami, M.; Ghassemi, T.; Labelle, E.R. Soil physio-chemical and biological indicators to evaluate the restoration of compacted soil following reforestation. Ecol. Indic. 2019, 101, 102–110. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Castaldi, S.; Ascoli, R.D.; Papa, S.; Carfora, A.; Marzaioli, R.; Fioretto, A. Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment. Appl. Soil Ecol. 2009, 43, 40–46. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Li, P.; Li, Q.; Xue, S. Ecoenzymatic stoichiometry and microbial nutrient limitation during secondary succession of natural grassland on the Loess Plateau, China. Soil Tillage Res. 2020, 200, 104605. [Google Scholar] [CrossRef]
- Xun, W.; Yan, R.; Ren, Y.; Jin, D.; Xiong, W.; Zhang, G.; Cui, Z.; Xin, X.; Zhang, R. Grazing-induced microbiome alterations drive soil organic carbon turnover and productivity in meadow steppe. Microbiome 2018, 6, 170. [Google Scholar] [CrossRef]
- Datt, N.; Singh, D. Enzymes in relation to soil biological properties and sustainability. In Sustainable Management of Soil and Environment; Springer: Berlin/Heidelberg, Germany, 2019; pp. 383–406. [Google Scholar]
- Chen, Y.; Wei, T.; Sha, G.; Zhu, Q.; Liu, Z.; Ren, K.; Yang, C. Soil enzyme activities of typical plant communities after vegetation restoration on the Loess Plateau, China. Appl. Soil Ecol. 2022, 170, 104292. [Google Scholar] [CrossRef]
- Prieto, L.H.; Bertiller, M.B.; Carrera, A.L.; Olivera, N.L. Soil enzyme and microbial activities in a grazing ecosystem of Patagonian Monte, Argentina. Geoderma 2011, 162, 281–287. [Google Scholar] [CrossRef]
- Fan, J.; Jin, H.; Zhang, C.; Zheng, J.; Zhang, J.; Han, G. Grazing intensity induced alternations of soil microbial community composition in aggregates drive soil organic carbon turnover in a desert steppe. Agric. Ecosyst. Environ. 2021, 313, 107387. [Google Scholar] [CrossRef]
- Lin, B.; Zhao, X.; Zheng, Y.; Qi, S.; Liu, X. Effect of grazing intensity on protozoan community, microbial biomass, and enzyme activity in an alpine meadow on the Tibetan Plateau. J. Soils Sediments 2017, 17, 2752–2762. [Google Scholar] [CrossRef]
- Byrnes, R.C.; Eastburn, D.J.; Tate, K.W.; Roche, L.M. A Global Meta-Analysis of Grazing Impacts on Soil Health Indicators. J. Environ. Qual. 2018, 47, 758–765. [Google Scholar] [CrossRef]
- Wang, L.; Pang, X.; Li, N.; Qi, K.; Huang, J.; Yin, C. Effects of vegetation type, fine and coarse roots on soil microbial communities and enzyme activities in eastern Tibetan plateau. Catena 2020, 194, 104694. [Google Scholar] [CrossRef]
- Kooch, Y.; Sanji, R.; Tabari, M. Increasing tree diversity enhances microbial and enzyme activities in temperate Iranian forests. Trees 2018, 32, 809–822. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.; Fanin, N.; Han, X.; Du, X.; Liu, H.; Li, Y.; Li, Q. Adaptation of soil micro-food web to elemental limitation: Evidence from the forest-steppe ecotone. Soil Biol. Biochem. 2022, 170, 108698. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Li, Y.; Li, B.; Han, X.; Li, Y.; Mahamood, M.; Li, Q. Soil community richness and composition jointly influence the multifunctionality of soil along the forest-steppe ecotone. Ecol. Indic. 2022, 139, 108900. [Google Scholar] [CrossRef]
- Schmidt, M.; Jochheim, H.; Kersebaum, K.; Lischeid, G.; Nendel, C. Gradients of microclimate, carbon and nitrogen in transition zones of fragmented landscapes—A review. Agric. For. Meteorol. 2017, 232, 659–671. [Google Scholar] [CrossRef]
- Joimel, S.; Cortet, J.; Jolivet, C.C.; Saby, N.P.A.; Chenot, E.D.; Branchu, P.; Consalès, J.N.; Lefort, C.; Morel, J.L.; Schwartz, C. Physico-chemical characteristics of topsoil for contrasted forest, agricultural, urban and industrial land uses in France. Sci. Total Environ. 2016, 545–546, 40–47. [Google Scholar] [CrossRef]
- Erdős, L.; Ambarlı, D.; Anenkhonov, O.A.; Bátori, Z.; Cserhalmi, D.; Kiss, M.; Kröel-Dulay, G.; Liu, H.; Magnes, M.; Molnár, Z.; et al. The edge of two worlds: A new review and synthesis on Eurasian forest-steppes. Appl. Veg. Sci. 2018, 21, 345–362. [Google Scholar] [CrossRef]
- Du, X.; Li, Y.; Han, X.; Ahmad, W.; Li, Q. Using high-throughput sequencing quantitatively to investigate soil nematode community composition in a steppe-forest ecotone. Appl. Soil Ecol. 2020, 152, 103562. [Google Scholar] [CrossRef]
- You, G.; Liu, B.; Zou, C.; Li, H.; Mckenzie, S.; He, Y.; Gao, J.; Jia, X.; Altaf Arain, M.; Wang, S.; et al. Sensitivity of vegetation dynamics to climate variability in a forest-steppe transition ecozone, north-eastern Inner Mongolia, China. Ecol. Indic. 2021, 120, 106833. [Google Scholar] [CrossRef]
- Erdős, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Körmöczi, L.; Lengyel, A.; Bátori, Z. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications. Ecol. Complex. 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Shi, X.Z.; Yu, D.S.; Warner, E.D.; Pan, X.Z.; Petersen, G.W.; Gong, Z.G.; Weindorf, D. Soil Database of 1:1,000,000 Digital Soil Survey and Reference System of the Chinese Genetic Soil Classification System. Soil Surv. Horiz. 2004, 45, 129–136. [Google Scholar] [CrossRef]
- Gao, F.; Chu, H.; Yu, G.; Xiang, K.; Hu, R.; Zhuang, Q.; Wang, X.; Zhao, Y.; Ma, L.; Sun, J.; et al. Study on species diversity and various grassland types in Hexigten Banner in Innner Mongolia. J. Qingdao Agric. Univ. 2019, 36, 19–25. [Google Scholar]
- Ren, H.; Eviner, V.T.; Gui, W.; Wilson, G.W.T.; Cobb, A.B.; Yang, G.; Zhang, Y.; Hu, S.; Bai, Y. Livestock grazing regulates ecosystem multifunctionality in semi-arid grassland. Funct. Ecol. 2018, 32, 2790–2800. [Google Scholar] [CrossRef]
- Marriott, C.A.; Hudson, G.; Hamilton, D.; Neilson, R.; Boag, B.; Handley, L.L.; Wishart, J.; Scrimgeour, C.M.; Robinson, D. Spatial variability of soil total C and N and their stable isotopes in an upland Scottish grassland. Plant Soil 1997, 196, 151–162. [Google Scholar] [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis: Part 3 Chemical Methods; Soil Science Society of America, Inc.: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar]
- Zhang, C.; Liu, G.; Xue, S.; Song, Z. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Setia, R.; Verma, S.L.; Marschner, P. Measuring microbial biomass carbon by direct extraction—Comparison with chloroform fumigation-extraction. Eur. J. Soil Biol. 2012, 53, 103–106. [Google Scholar] [CrossRef]
- Bhuyan, P.; Deka, P.; Prakash, A.; Balachandran, S.; Hoque, R.R. Chemical characterization and source apportionment of aerosol over mid Brahmaputra Valley, India. Environ. Pollut. 2018, 234, 997–1010. [Google Scholar] [CrossRef]
- Gong, P.; Liu, H.; Zhang, M.; Li, C.; Wang, J.; Huang, H.; Clinton, N.; Ji, L.; Li, W.; Bai, Y.; et al. Stable classification with limited sample: Transferring a 30-m resolution sample set collected in 2015 to mapping 10-m resolution global land cover in 2017. Sci. Bull. 2019, 64, 370–373. [Google Scholar] [CrossRef]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.Y.H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Chang. Biol. 2019, 25, 1078–1088. [Google Scholar] [CrossRef]
- Donovan, M.; Monaghan, R. Impacts of grazing on ground cover, soil physical properties and soil loss via surface erosion: A novel geospatial modelling approach. J. Environ. Manag. 2021, 287, 112206. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Herbivore-mediated linkages between aboveground and belowground communities. Ecology 2003, 84, 2258–2268. [Google Scholar] [CrossRef]
- Zhang, S.; Wei, Y.; Liu, N.; Wang, Y.; Manlike, A.; Zhang, Y.; Zhang, B. Mowing Facilitated Shoot and Root Litter Decomposition Compared with Grazing. Plants 2022, 11, 846. [Google Scholar] [CrossRef]
- Bagchi, S.; Ritchie, M.E. Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition. Ecol. Lett. 2010, 13, 959–968. [Google Scholar] [CrossRef]
- Manzano, M.G.; Návar, J. Processes of desertification by goats overgrazing in the Tamaulipan thornscrub (matorral) in north-eastern Mexico. J. Arid Environ. 2000, 44, 1–17. [Google Scholar] [CrossRef]
- Wu, J.; Wang, H.; Li, G.; Ma, W.; Wu, J.; Gong, Y.; Xu, G. Vegetation degradation impacts soil nutrients and enzyme activities in wet meadow on the Qinghai-Tibet Plateau. Sci. Rep. 2020, 10, 21271. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Ren, H.; Zhao, T.; Li, Q.; Wang, S.; Zhang, Y.; Xiao, F.; Wang, X. BAK1 mediates light intensity to phosphorylate and activate catalases to regulate plant growth and development. Int. J. Mol. Sci. 2020, 21, 1437. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, W.; Sai, X.; Chun, F.; Li, X.; Lu, X.; Wang, H. Grazing altered soil aggregates, nutrients and enzyme activities in a Stipa kirschnii steppe of Inner Mongolia. Soil Tillage Res. 2022, 219, 105327. [Google Scholar] [CrossRef]
- Chen, J.; Szwedo, J.; Wang, B.; Zheng, Y.; Jiang, H.; Jiang, T.; Wang, X.; Zhang, H. A new bizarre cicadomorph family in mid-Cretaceous Burmese amber (Hemiptera, Clypeata). Cretac. Res. 2019, 97, 1–15. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Hao, Y.; Wang, Y. Intermediate grazing intensities by sheep increase soil bacterial diversities in an Inner Mongolian steppe. Biol. Fertil. Soils 2010, 46, 817–824. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.H.; Chen, X.; Huang, Z. Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration. Nat. Commun. 2019, 10, 1332. [Google Scholar] [CrossRef]
- Zhu, G.; Shangguan, Z.; Hu, X.; Deng, L. Effects of land use changes on soil organic carbon, nitrogen and their losses in a typical watershed of the Loess Plateau, China. Ecol. Indic. 2021, 133, 108443. [Google Scholar] [CrossRef]
- Mustafa, A.; Minggang, X.; Ali Shah, S.A.; Abrar, M.M.; Nan, S.; Baoren, W.; Zejiang, C.; Saeed, Q.; Naveed, M.; Mehmood, K.; et al. Soil aggregation and soil aggregate stability regulate organic carbon and nitrogen storage in a red soil of southern China. J. Environ. Manag. 2020, 270, 110894. [Google Scholar] [CrossRef]
- Zhang, J.; Zuo, X.; Zhou, X.; Lv, P.; Lian, J.; Yue, X. Long-term grazing effects on vegetation characteristics and soil properties in a semiarid grassland, northern China. Environ. Monit. Assess. 2017, 189, 216. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Ludwig, M.; Achtenhagen, J.; Miltner, A.; Eckhardt, K.; Leinweber, P.; Emmerling, C.; Thiele-Bruhn, S. Microbial contribution to SOM quantity and quality in density fractions of temperate arable soils. Soil Biol. Biochem. 2015, 81, 311–322. [Google Scholar] [CrossRef]
- Sun, X.; Tang, Z.; Ryan, M.G.; You, Y.; Sun, O.J. Changes in soil organic carbon contents and fractionations of forests along a climatic gradient in China. For. Ecosyst. 2019, 6, 1. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z. Recalcitrant carbon controls the magnitude of soil organic matter mineralization in temperate forests of northern China. For. Ecosyst. 2018, 5, 17. [Google Scholar] [CrossRef]
- Lu, X.; Kelsey, K.C.; Yan, Y.; Sun, J.; Wang, X.; Cheng, G.; Neff, J.C. Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai—Tibetan Plateau: A synthesis. Ecosphere 2017, 8, e1656. [Google Scholar] [CrossRef]
- Wang, B.; Liu, D.; Yang, J.; Zhu, Z.; Darboux, F.; Jiao, J.; An, S. Effects of forest floor characteristics on soil labile carbon as varied by topography and vegetation type in the Chinese Loess Plateau. Catena 2021, 196, 104825. [Google Scholar] [CrossRef]
- Tan, X.; Chang, S.X.; Kabzems, R. Soil compaction and forest floor removal reduced microbial biomass and enzyme activities in a boreal aspen forest soil. Biol. Fertil. Soils 2008, 44, 471–479. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, Q.; Noll, L.; Hu, Y.; Wanek, W. Environmental effects on soil microbial nitrogen use efficiency are controlled by allocation of organic nitrogen to microbial growth and regulate gross N mineralization. Soil Biol. Biochem. 2019, 135, 304–315. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Sethi, S.; Datta, A.; Gupta, B.L.; Gupta, S. Optimization of Cellulase Production from Bacteria Isolated from Soil. ISRN Biotechnol. 2013, 2013, 985685. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Kooijman, A.M.; Smit, A. Grazing as a measure to reduce nutrient availability and plant productivity in acid dune grasslands and pine forests in The Netherlands. Ecol. Eng. 2001, 17, 63–77. [Google Scholar] [CrossRef]
- Martina, S.; Petr, B. Effects of soil properties and management on the activity of soil organic matter transforming enzymes and the quantification of soil-bound and free activity. Plant Soil 2011, 338, 99–110. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, S.; Hu, Y.; Chen, Y.; Jiang, H.; Duan, B.; Lu, X. Links between chemical composition of soil organic matter and soil enzyme activity in alpine grassland ecosystems of the Tibetan Plateau. Catena 2022, 218, 106565. [Google Scholar] [CrossRef]
- Ma, C.; Hao, X.; He, F.; Baoyin, T.; Yang, J.; Dong, S. Effects of seasonal grazing on plant and soil microbial diversity of typical temperate grassland. Front. Plant Sci. 2022, 13, 1040377. [Google Scholar] [CrossRef]
- Ghosh, A.; Mahanta, S.K.; Manna, M.C.; Singh, S.; Bhattacharyya, R.; Tyagi, V.C.; Singh, J.B.; Ram, S.N.; Srinivasan, R.; Singh, A.K.; et al. Long-Term Grazing Mediates Soil Organic Carbon Dynamics by Reorienting Enzyme Activities and Elemental Stoichiometry in Semi-arid Tropical Inceptisol. J. Soil Sci. Plant Nutr. 2022, 22, 1422–1433. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, T.; Lv, M.; Fang, Y.; Liu, R.; Luo, Y.; Xu, C.; Tian, X. Grazing Effects of Soil Fauna on White-Rot Fungi: Biomass, Enzyme Production and Litter Decomposition Ability. J. Fungi 2022, 8, 348. [Google Scholar] [CrossRef]
- He, P.; Fontana, S.; Ma, C.; Liu, H.; Xu, L.; Wang, R.; Jiang, Y.; Li, M.H. Using leaf traits to explain species co-existence and its consequences for primary productivity across a forest-steppe ecotone. Sci. Total Environ. 2023, 859, 160139. [Google Scholar] [CrossRef]
- Illiger, P.; Schmidt, G.; Walde, I.; Hese, S.; Kudrjavzev, A.E.; Kurepina, N.; Mizgirev, A.; Stephan, E.; Bondarovich, A.; Frühauf, M. Estimation of regional soil organic carbon stocks merging classified land-use information with detailed soil data. Sci. Total Environ. 2019, 695, 133755. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Deng, L.; Guo, X.; Han, F.; Ju, W.; Wang, X.; Chen, H.; Tan, W.; Zhang, X. Patterns of soil microbial nutrient limitations and their roles in the variation of soil organic carbon across a precipitation gradient in an arid and semi-arid region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef]
- Wahsha, M.; Nadimi-Goki, M.; Fornasier, F.; Al-Jawasreh, R.; Hussein, E.I.; Bini, C. Microbial enzymes as an early warning management tool for monitoring mining site soils. Catena 2017, 148, 40–45. [Google Scholar] [CrossRef]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps in the forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Jourgholami, M.; Ramineh, A.; Zahedi Amiri, G.; Labelle, E. The Influence of Slope Positions on the Recovery Response of Compacted Soil Properties and Enzyme Activity in an Oriental Beech Stand in the Hyrcanian Forests, Iran. Sustainability 2019, 11, 1940. [Google Scholar] [CrossRef]
- Fazlollahi Mohammadi, M.; Jalali, S.G.; Kooch, Y.; Said-Pullicino, D. The effect of landform on soil microbial activity and biomass in a Hyrcanian oriental beech stand. Catena 2017, 149, 309–317. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Site | Treatment | pH | Slope (°) | Aspect | Soil Type | Geographic Coordination | Altitude (m) | Plant Community | Main Community Species |
|---|---|---|---|---|---|---|---|---|---|
| NS1 | Grazing | 7.71 | 18 | South | Gleyic Chernozems | 43.595° E 117.322° N | 1490 | Populus davidiana + Betula platyphylla | Rosa davurica, Galium verum, Deyeuxia angustifolia |
| NS2 | Non-grazing | 6.02 | 16 | South | |||||
| MF1 | Grazing | 6.29 | 15 | South | Gleyic Chernozems | 43.575° E 117.341° N | 1525 | Betula platyphylla + Pinus sylvestris | Sanguisorba officinalis, Taraxacum mongolicum, Rosa davurica, Spiraea salicifolia |
| MF2 | Non-grazing | 5.56 | 10 | South | |||||
| CP1 | Grazing | 5.94 | 11 | South | Gleyic Chernozems | 43.428° E 117.242° N | 1405 | Pinus sylvestris | Carex duriuscula, Potentilla bifurca |
| CP2 | Non-grazing | 5.99 | 13 | South | |||||
| GL1 | Grazing | 7.95 | 17 | South | Luvic Kastanozems | 43.604° E 117.410° N | 1540 | Leymus chinensis | Carex duriuscula, Cleistogenes squarros, Agropyron cristatum, Potentilla acaulis |
| GL2 | Non-grazing | 6.79 | 18 | South |
| Parameters | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| MBC | 0.168 | 0.117 | 0.864 | 0.406 | 0.105 |
| MBN | 0.207 | 0.290 | −0.428 | 0.782 | 0.168 |
| TN | 0.641 | −0.037 | 0.061 | −0.115 | 0.026 |
| TOC | 0.647 | −0.071 | −0.004 | −0.089 | −0.049 |
| NN | 0.613 | −0.125 | 0.086 | −0.239 | 0.149 |
| AN | 0.069 | 0.646 | −0.061 | −0.358 | 0.612 |
| pH | −0.637 | 0.027 | 0.072 | 0.047 | 0.009 |
| Grazing | −0.263 | −0.502 | 0.052 | 0.088 | 0.731 |
| Vegetation types | 0.315 | −0.459 | −0.217 | 0.088 | 0.161 |
| Eigenvalue | 2.176 | 1.313 | 0.973 | 0.867 | 0.624 |
| Proportion of variance (%) | 52.61% | 19.15% | 10.53% | 8.34% | 4.32% |
| Cumulative proportion (%) | 52.61% | 71.77% | 82.29% | 90.64% | 94.96% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Jia, Z.; Li, Q.; He, L.; Tian, J.; Ding, W.; Liu, T.; Gao, Y.; Zhang, J.; Han, D.; et al. Grazing Impacts on Soil Enzyme Activities Vary with Vegetation Types in the Forest-Steppe Ecotone of Northeastern China. Forests 2023, 14, 2292. https://doi.org/10.3390/f14122292
Wang L, Jia Z, Li Q, He L, Tian J, Ding W, Liu T, Gao Y, Zhang J, Han D, et al. Grazing Impacts on Soil Enzyme Activities Vary with Vegetation Types in the Forest-Steppe Ecotone of Northeastern China. Forests. 2023; 14(12):2292. https://doi.org/10.3390/f14122292
Chicago/Turabian StyleWang, Long, Zhiqing Jia, Qingxue Li, Lingxianzi He, Jiapeng Tian, Wei Ding, Tao Liu, Ya Gao, Jiapeng Zhang, Dong Han, and et al. 2023. "Grazing Impacts on Soil Enzyme Activities Vary with Vegetation Types in the Forest-Steppe Ecotone of Northeastern China" Forests 14, no. 12: 2292. https://doi.org/10.3390/f14122292