
Genome-Wide Analysis and Expression Profiling of YUCCA Gene Family in Developmental and Environmental Stress Conditions in Tea Plant (Camellia sinensis)
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Tea CsYUC Genes
2.2. Multiple Sequence Alignment and Phylogenetic Analysis
2.3. Gene Structure and Conserved Motifs Analysis of CsYUC Numbers
2.4. Physical and Chemical Properties Analysis of CsYUCs
2.5. Analysis of Cis-Acting Elements in the Promoter Regions of CsYUC Genes
2.6. Open Available RNA-Seq Data Analysis
2.7. RNA-Seq and qRT-PCR Detection
2.8. Statistical Analysis
3. Results and Discussion
3.1. Identification and Bioinformatics Analysis of the CsYUC Gene Family in Tea Plants
3.2. Multiple Sequence Alignment and Phylogenetic Analysis of the CsYUC Gene Family
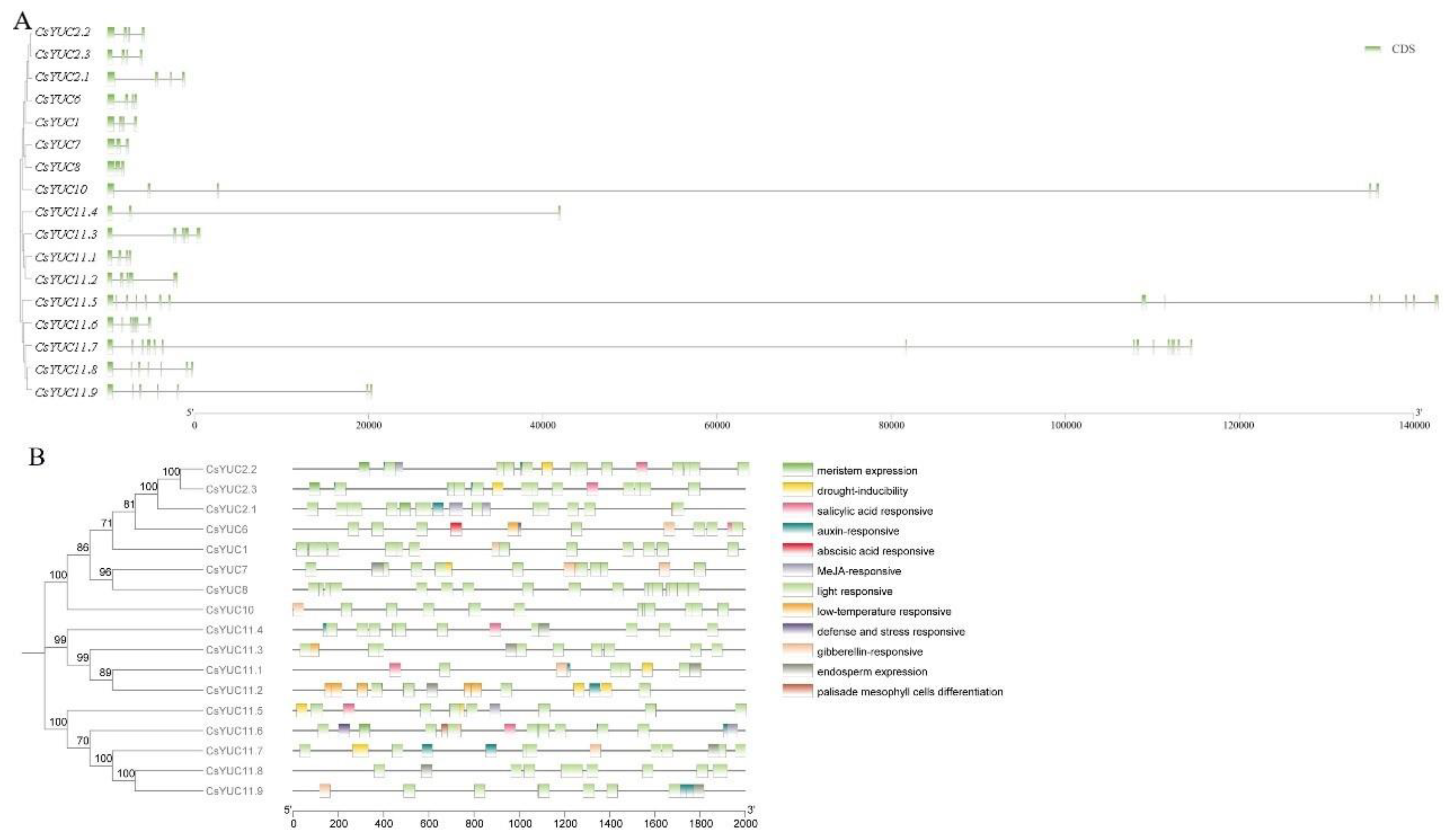
3.3. Gene Structure and Conserved Domain Analysis of CsYUC Genes
3.4. The Cis-Acting Elements of CsYUC Genes
3.5. Expression Profiles of the CsYUC Family Numbers in Tea Plants
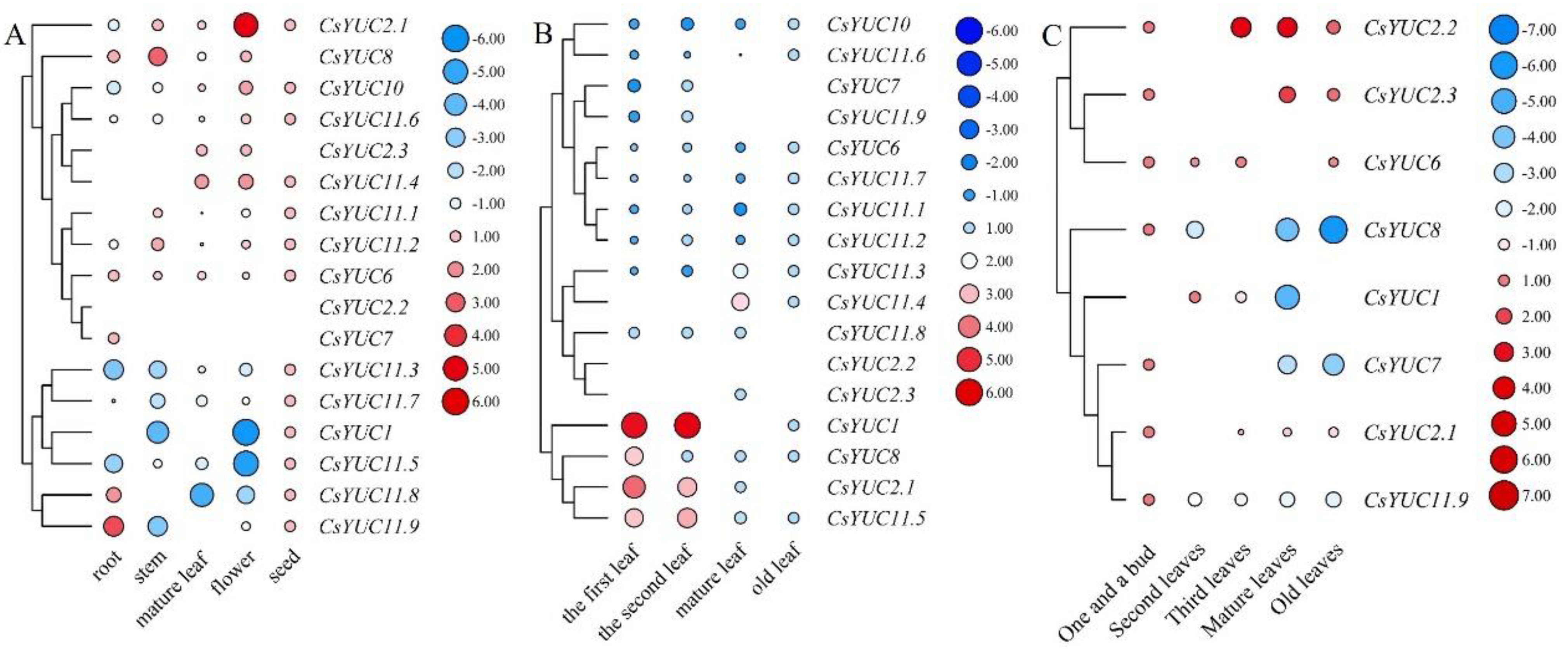
3.5.1. CsYUC Gene Expression Analysis during the Development Processes of Tea Plants
Expression Profiles of CsYUCs in Different Tissues
The Regulatory Roles of CsYUC Genes in Tea Leaf Development
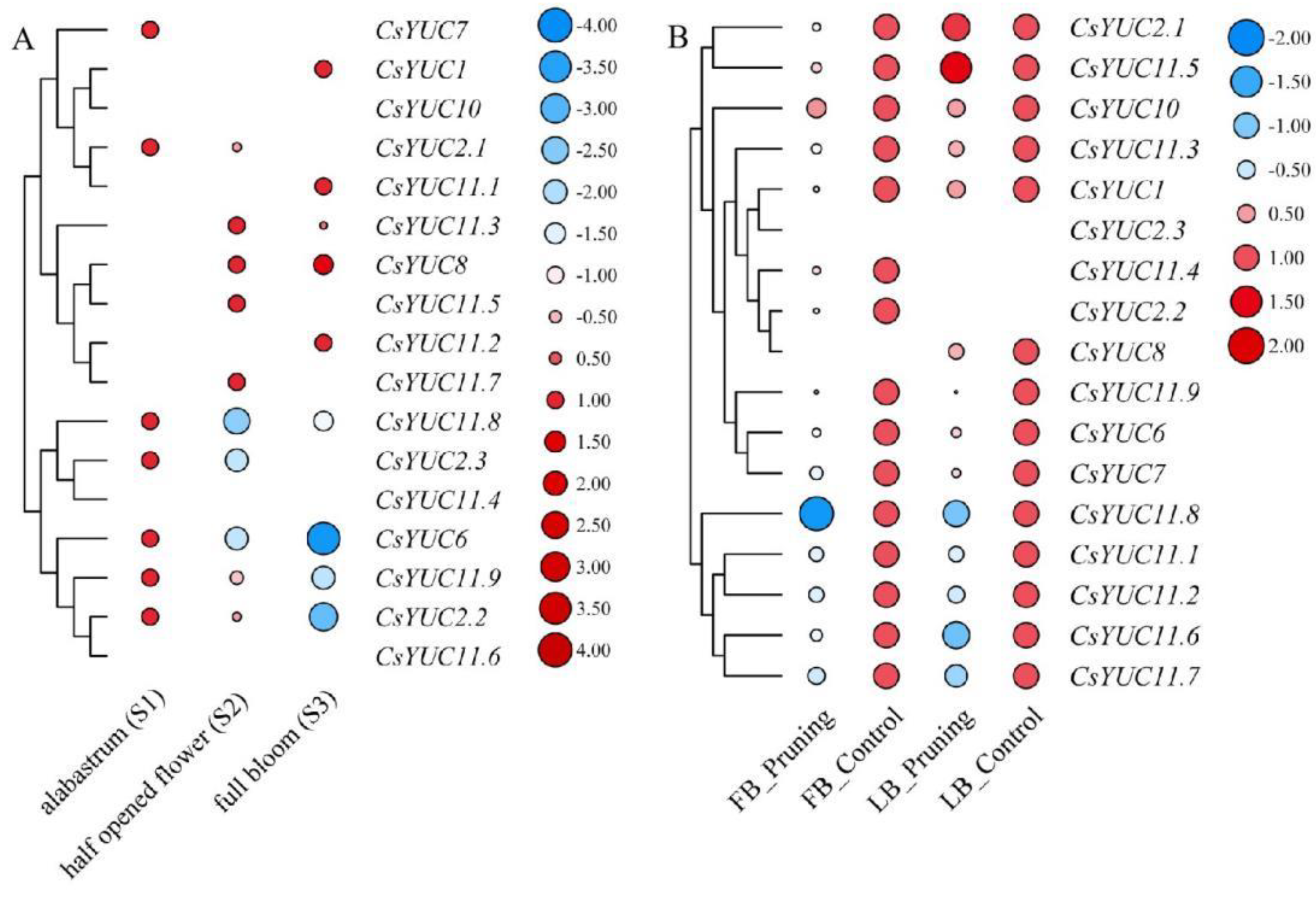
The Roles of CsYUC Genes in Tea Flower Development
The Regulatory Roles of CsYUCs in the Lateral Bud Development of Tea Plants Induced by Pruning
3.5.2. The Expression of CsYUC Numbers under Several Environmental Stress Conditions
The Expression of CsYUC Numbers under Temperature Stresses
- The Roles of CsYUC Genes in Response to High Temperature
- 2.
- The Regulatory Roles of CsYUC Genes in Response to Low Temperature
The Roles of CsYUC Genes in Response to Drought Stress
The Regulatory Roles of CsYUC Genes in Response to NaCl Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.; Xu, T.; Wang, H.; Feng, D. Genome-wide identification and expression analysis of the TaYUCCA gene family in wheat. Mol. Biol. Rep. 2021, 48, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, H.; Li, H.; Teng, R.; Zhuang, J. Genome-based identification and analysis of the genes involved in auxin biosynthesis and signal transduction during tea plant leaf development. Sci. Hortic. 2020, 261, 109030. [Google Scholar] [CrossRef]
- Song, C.; Zhang, D.; Zheng, L.; Shen, Y.; Zuo, X.; Mao, J.; Meng, Y.; Wu, H.; Zhang, Y.; Liu, X.; et al. Genome-wide identification and expression profiling of the YUCCA gene family in Malus domestica. Sci. Rep. 2020, 10, 10866. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The roles of auxin biosynthesis YUCCA gene family in plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef]
- Yan, S.; Che, G.; Ding, L.; Chen, Z.; Liu, X.; Wang, H.; Zhao, W.; Ning, K.; Zhao, J.; Tesfamichael, K.; et al. Different cucumber CsYUC genes regulate response to abiotic stresses and flower development. Sci. Rep. 2016, 6, 20760. [Google Scholar] [PubMed]
- Park, H.C.; Cha, J.-Y.; Yun, D.-J. Roles of YUCCAs in auxin biosynthesis and drought stress responses in plants. Plant Signal Behav. 2013, 6, 337–349. [Google Scholar]
- Blakeslee, J.J.; Rossi, T.S.; Kriechbaumer, V. Auxin biosynthesis: Spatial regulation and adaptation to stress. J. Exp. Bot. 2019, 70, 5041–5049. [Google Scholar] [CrossRef]
- Uc-Chuc, M.A.; Kú-González, A.F.; Jiménez-Ramírez, I.A.; Loyola-Vargas, V.M. Identification, analysis, and modeling of the YUCCA protein family genome-wide in Coffea canephora. Proteins 2022, 90, 1005–1024. [Google Scholar] [CrossRef]
- Pérez-Alonso, M.M.; Sánchez-Parra, B.; Ortiz-García, P.; Santamaría, M.E.; Díaz, I.; Pollmann, S. Jasmonic acid-dependent MYC transcription factors bind to a tandem G-Box motif in the YUCCA8 and YUCCA9 promoters to regulate biotic stress responses. Int. J. Mol. Sci. 2021, 22, 9768. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin biosynthesis by the YUCCA genes in rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef]
- Poulet, A.; Kriechbaumer, V. Bioinformatics Analysis of phylogeny and transcription of TAA/YUC auxin biosynthetic genes. Int. J. Mol. Sci. 2017, 18, 1791. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Zhu, Y.; Yu, Y.; Zhao, Q.Y.; Wang, S.J.; Wang, X.C.; Yao, M.Z.; Luo, D.; Li, X.; Chen, L.; et al. Global transcriptome and gene regulation network for secondary metabolite biosynthesis of tea plant (Camellia sinensis). BMC Genom. 2015, 16, 560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S. Genome-wide analysis and expression patterns of the YUCCA genes in maize. J. Genet. Genomics 2015, 42, 707–710. [Google Scholar]
- Wang, Y.; Liu, H.; Wang, S.; Li, H. Genome-wide identification and expression analysis of the YUCCA gene family in soybean (Glycine max L.). Plant Growth Regul. 2017, 81, 265–275. [Google Scholar] [CrossRef]
- Kim, J.I.; Baek, D.; Park, H.C.; Chun, H.J.; Oh, D.-H.; Lee, M.K.; Cha, J.-Y.; Kim, W.-Y.; Kim, M.C.; Chung, W.S.; et al. Overexpression of Arabidopsis YUCCA6 in potato results in high-auxin developmental phenotypes and enhanced resistance to water deficit. Mol. Plant 2013, 6, 337–349. [Google Scholar] [CrossRef]
- Qin, M.; Wang, J.; Zhang, T.; Hu, X.; Liu, R.; Gao, T.; Zhao, S.; Yuan, Y.; Zheng, J.; Wang, Z.; et al. Genome-wide identification and analysis on YUCCA gene family in Isatisindigotica Fort. and IiYUCCA6-1 functional exploration. Int. J. Mol. Sci. 2020, 21, 2188. [Google Scholar] [CrossRef]
- Liu, H.; Xie, W.-F.; Zhang, L.; Valpuesta, V.; Ye, Z.-W.; Gao, Q.-H.; Duan, K. Auxin biosynthesis by the YUCCA6 flavin monooxygenase gene in woodland strawberry. J. Integr. Plant Biol. 2014, 56, 350–363. [Google Scholar] [CrossRef]
- Wang, Y.X.; Liu, Z.W.; Wu, Z.J.; Li, H.; Wang, W.L.; Cui, X.; Zhuang, J. Genome-wide identification and expression analysis of GRAS family transcription factors in tea plant (Camellia sinensis). Sci. Rep. 2018, 8, 3949. [Google Scholar] [CrossRef]
- Wang, X.; Feng, H.; Chang, Y.; Ma, C.; Wang, L.; Hao, X.; Li, A.; Cheng, H.; Wang, L.; Cui, P.; et al. Population sequencing enhances understanding of tea plant evolution. Nat. Commun. 2020, 11, 4447. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, Y.; Zhang, M.; Liu, Y.; Kong, L.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Identification, expression, and comparative genomicanalysis of the IPT and CKX gene families in Chinese cabbage (Brassica raga ssp. pekinensis). BMC Genom. 2013, 14, 594. [Google Scholar]
- Zhang, L.; Li, M.; Fu, J.; Huang, X.; Yan, P.; Ge, S.; Li, Z.; Bai, P.; Zhang, L.; Han, W.; et al. Genome-wide identification and expression analysis of isopentenyl transferase family genes during development and resistance to abiotic stresses in tea plant (Camellia sinensis). Plants 2022, 11, 2243. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Byung-guk, K.; Lori, D.O.; Yi, L.; Cheng, Z.-M. Identification of the flavin-dependent monooxygenase-encoding YUCCA gene family in Populus trichocarpa and their expression in vegetative tissues and in response to hormone and environmental stresses. Plant Cell. Tiss. Organ Cult. 2009, 97, 271–283. [Google Scholar]
- Liu, F.; Wang, Y.; Ding, Z.; Zhao, L.; Xiao, J.; Wang, L.; Ding, S. Transcriptomic analysis of flower development in tea (Camelliasinensis (L.)). Gene 2017, 631, 39–51. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, L.; Duan, K.; Zhu, Z.-P.; Ye, Z.-W.; Gao, Q.-H. YUCCA type auxin biosynthesis genes encoding flavin monooxygenases in melon: Genome-wide identification and developmental expression analysis. S. Afr. J. Bot. 2016, 102, 142–152. [Google Scholar] [CrossRef]
- Kim, J.I.; Murphy, A.S.; Baek, D.; Lee, S.-W.; Yun, D.-J.; Bressan, R.A.; Narasimhan, M.L. YUCCA6 over-expression demonstrates auxin function in delaying leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 3981–3992. [Google Scholar] [CrossRef]
- Gallavotti, A.; Barazesh, S.; Malcomber, S.; Hall, D.; Jackson, D.; Schmidt, R.J.; Steen, P.M. sparse inflorescence1 encodes a monocot-specific YUCCA-like gene required for vegetative and eproductive development in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 15196–15201. [Google Scholar] [CrossRef]
- Zheng, C.; Zhao, L.; Wang, Y.; Shen, J.; Zhang, Y.; Jia, S.; Li, Y.; Ding, Z. Integrated RNA-Seq and sRNA-Seq analysis identifies chilling and freezing responsive key molecular players and pathways in tea plant (Camellia sinensis). PLoS ONE 2015, 10, e0125031. [Google Scholar] [CrossRef]
- Li, N.; Yue, C.; Cao, H.; Qian, W.; Hao, X.; Wang, Y.; Wang, L.; Ding, C.; Wang, X.; Yang, Y. Transcriptome sequencing dissection of the mechanisms underlying differential cold sensitivity in young and mature leaves of the tea plant (Camellia sinensis). J. Plant Physiol. 2018, 224–225, 144–155. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, D.; Zhou, L.; Zhang, X.; Liao, J.; Duan, Y.; Wen, B.; Ma, Y.; Wang, Y.; Fang, W.; et al. Transcriptomic and metabolomic profiling of Camellia sinensis L. cv. ‘Suchazao’ exposed to temperature stresses reveals modification in proteinsynthesis and photosynthetic andanthocyanin biosynthetic pathways. Tree Physiol. 2019, 39, 1583–1599. [Google Scholar] [CrossRef] [PubMed]
- Sakata, T.; Yagihashi, N.; Higashitani, A. Tissue-specific auxin signaling in response to temperature fluctuation. Plant Signal Behav. 2010, 5, 1510–1512. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Li, C.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Asami, T.; Sabir, J.S.M.; Park, H.C.; Lee, S.Y.; Bohnert, H.J.; Bressan, R.A.; Pardo, J.M.; Yun, D.-J. A novel thiol-reductase activity of Arabidopsis YUC6 confers drought tolerance independently of auxin biosynthesis. Nat. Commun. 2015, 6, 8041. [Google Scholar]
- Cha, J.Y.; Kim, W.Y.; Kang, S.B.; Kim, J.; Baek, D.; Jung, I.J.; Kim, M.R.; Li, N.; Kim, H.J.; Nakajima, M.; et al. Transcriptomic analysis of the effects of three different light treatments on the biosynthesis of characteristic compounds in the tea plant by RNA-Seq. Tree Genet. Genomes 2016, 12, 118. [Google Scholar]
- Lee, M.; Jung, J.H.; Han, D.Y.; Seo, P.J.; Park, W.J.; Park, C.M. Activation of a flavin monooxygenase gene YUCCA7 enhances drought resistance in Arabidopsis. Planta 2012, 235, 923–938. [Google Scholar] [CrossRef]
- Wan, S.; Wang, W.; Zhou, T.; Zhang, Y.; Chen, J.; Xiao, B.; Yang, Y.; Yu, Y. Transcriptomic analysis reveals the molecular mechanisms of Camellia sinensis in response to salt stress. Plant Growth Regul. 2018, 84, 481–492. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | CDS Length (bp) | Protein | |||
|---|---|---|---|---|---|
| Name | Locus ID | Length (aa) | pI | MS (kDa) | |
| CsYUC1 | GWHPACFB006323 | 1251 | 416 | 8.94 | 46.80 |
| CsYUC2.1 | GWHPACFB009508 | 1314 | 437 | 8.68 | 48.96 |
| CsYUC2.2 | GWHPACFB020502 | 1278 | 425 | 8.74 | 47.68 |
| CsYUC2.3 | GWHPACFB020503 | 1062 | 353 | 8.02 | 39.32 |
| CsYUC6 | GWHPACFB006291 | 1266 | 421 | 8.85 | 46.65 |
| CsYUC7 | GWHPACFB026074 | 1269 | 422 | 9.10 | 46.85 |
| CsYUC8 | GWHPACFB005110 | 1344 | 447 | 8.44 | 49.89 |
| CsYUC10 | GWHPACFB005354 | 1353 | 450 | 8.77 | 50.53 |
| CsYUC11.1 | GWHPACFB020329 | 1020 | 339 | 6.31 | 38.66 |
| CsYUC11.2 | GWHPACFB020327 | 1560 | 519 | 6.34 | 59.35 |
| CsYUC11.3 | GWHPACFB019218 | 1587 | 528 | 6.51 | 60.09 |
| CsYUC11.4 | GWHPACFB013041 | 885 | 295 | 7.07 | 33.36 |
| CsYUC11.5 | GWHPACFB010117 | 2685 | 894 | 5.98 | 102.27 |
| CsYUC11.6 | GWHPACFB005904 | 1416 | 471 | 5.79 | 52.86 |
| CsYUC11.7 | GWHPACFB019879 | 2580 | 859 | 6.34 | 98.91 |
| CsYUC11.8 | GWHPACFB011461 | 1203 | 400 | 6.02 | 45.61 |
| CsYUC11.9 | GWHPACFB011462 | 1302 | 433 | 6.07 | 49.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Jin, S.; Bai, P.; Ge, S.; Yan, P.; Li, Z.; Zhang, L.; Han, W.; Zeng, J.; Li, X. Genome-Wide Analysis and Expression Profiling of YUCCA Gene Family in Developmental and Environmental Stress Conditions in Tea Plant (Camellia sinensis). Forests 2023, 14, 2185. https://doi.org/10.3390/f14112185
Zhang L, Jin S, Bai P, Ge S, Yan P, Li Z, Zhang L, Han W, Zeng J, Li X. Genome-Wide Analysis and Expression Profiling of YUCCA Gene Family in Developmental and Environmental Stress Conditions in Tea Plant (Camellia sinensis). Forests. 2023; 14(11):2185. https://doi.org/10.3390/f14112185
Chicago/Turabian StyleZhang, Liping, Shan Jin, Peixian Bai, Shibei Ge, Peng Yan, Zhengzhen Li, Lan Zhang, Wenyan Han, Jianming Zeng, and Xin Li. 2023. "Genome-Wide Analysis and Expression Profiling of YUCCA Gene Family in Developmental and Environmental Stress Conditions in Tea Plant (Camellia sinensis)" Forests 14, no. 11: 2185. https://doi.org/10.3390/f14112185