
Changes in Leaf Functional Traits Driven by Environmental Filtration in Different Monsoon Tropical Forest Types

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Setting
2.2. Data Collection
2.2.1. Vegetation Survey
2.2.2. Collection of Environmental Factors
2.2.3. Collection of Functional Traits
2.3. Statistical Analysis
3. Results
3.1. Comparisons Species Composition across Three Forest Types
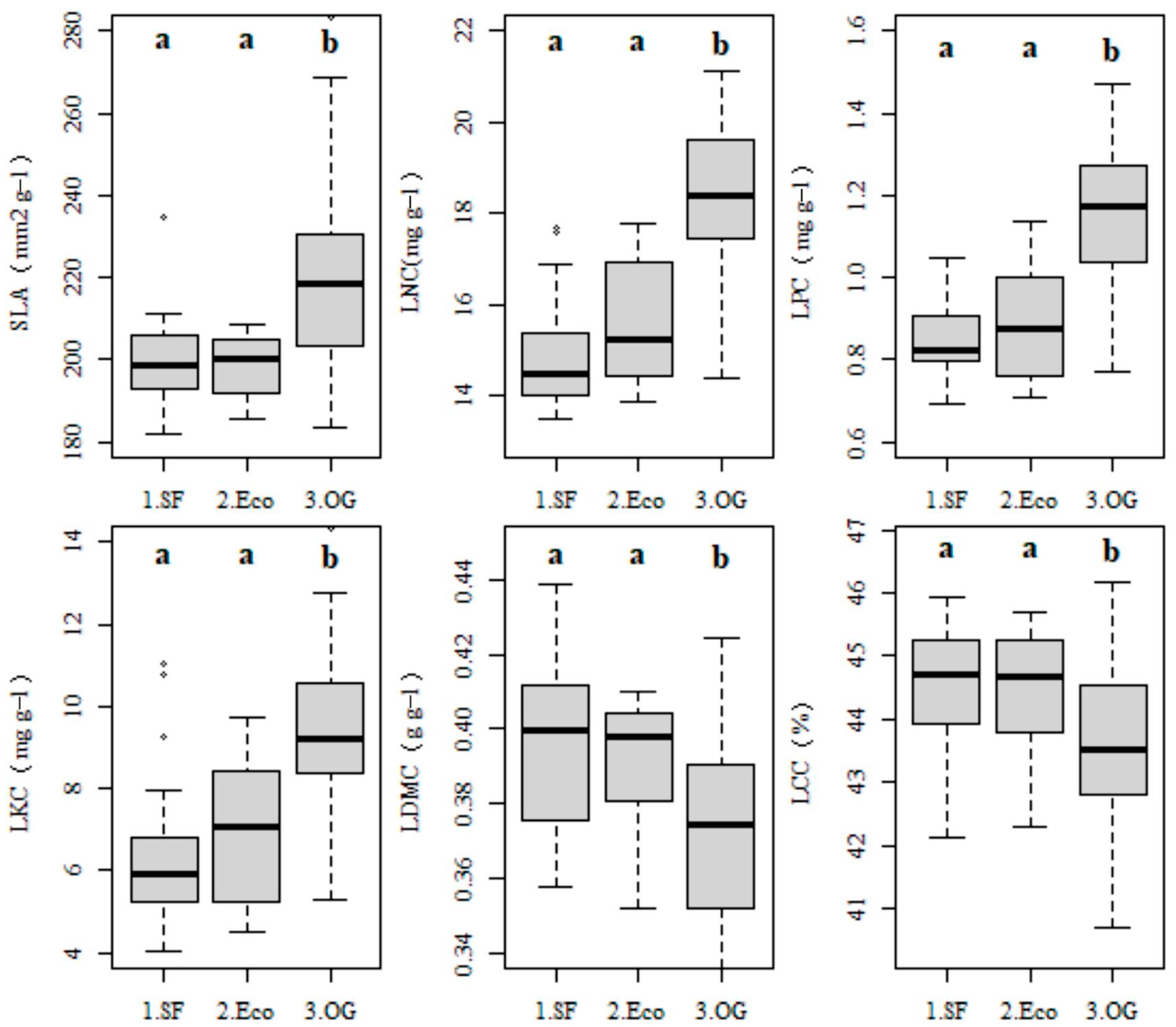
3.2. Forest Types Shaped Leaf Functional Trait Variations
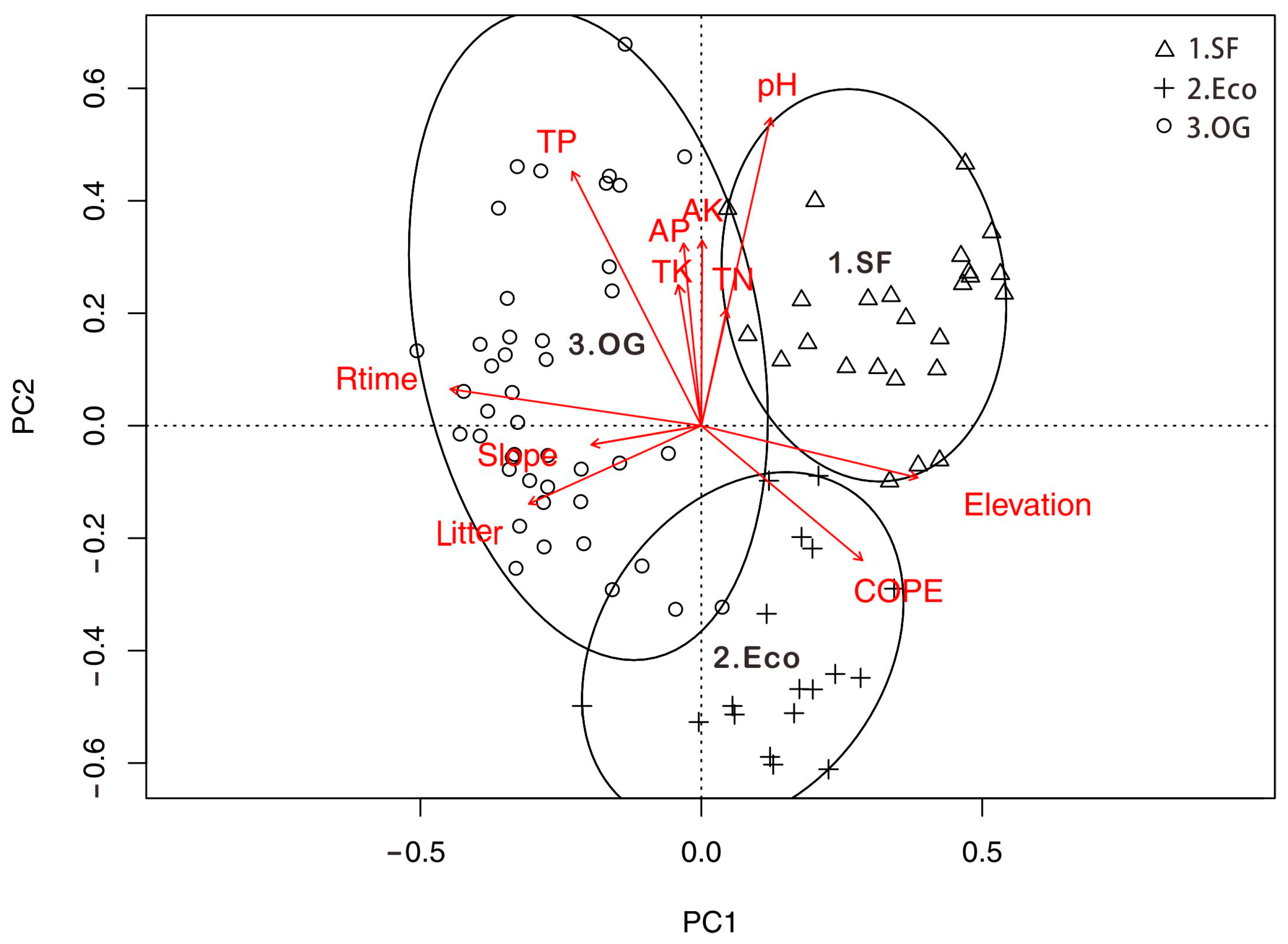
3.3. Relationship between Environmental Factors and Leaf Functional Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Li, S.; Wang, H.; Gou, W.; White, J.F.; Kingsley, K.L.; Wu, G.; Su, P. Leaf functional traits of dominant desert plants in the Hexi Corridor, Northwestern China: Trade-off relationships and adversity strategies. Glob. Ecol. Conserv. 2021, 28, e01666. [Google Scholar] [CrossRef]
- Oktavia, D.; Jin, G. Variations in leaf morphological and chemical traits in response to life stages, plant functional types, and habitat types in an old-growth temperate forest. Basic Appl. Ecol. 2020, 49, 22–33. [Google Scholar] [CrossRef]
- Duan, X.; Jia, Z.; Li, J.; Wu, S. The influencing factors of leaf functional traits variation of Pinus densiflora Sieb. et Zucc. Glob. Ecol. Conserv. 2022, 38, e02177. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, K.; Zang, R.; Ding, Y. Environmental filtering of species with different functional traits into plant assemblages across a tropical coniferous-broadleaved forest ecotone. Plant Soil 2014, 380, 361–374. [Google Scholar] [CrossRef]
- Khan, A.; Yan, L.; Mahadi Hasan, M.; Wang, W.; Xu, K.; Zou, G.; Liu, X.-D.; Fang, X.-W. Leaf traits and leaf nitrogen shift photosynthesis adaptive strategies among functional groups and diverse biomes. Ecol. Indic. 2022, 141, 109098. [Google Scholar] [CrossRef]
- Charles, B.; Chase, M.H.; Pociask, G.; Bhattarai, R.; Matthews, J.W. Can functional leaf traits be used for monitoring wetland restoration? A comparison between commonly used monitoring metrics and functional leaf traits. Ecol. Indic. 2022, 140, 109032. [Google Scholar] [CrossRef]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Díaz, S.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef]
- Mahayani, N.P.D.; Slik, F.J.W.; Webb, E.L.; Savini, T.; Gale, G.A. Changes in tree functional composition and forest functioning ten years after logging and thinning interventions in Bornean tropical forests. For. Ecol. Manag. 2022, 506, 119948. [Google Scholar] [CrossRef]
- Botha, M.; Greve, M. How do functional traits change during plant succession? A review. S. Afr. J. Bot. 2018, 115, 281. [Google Scholar] [CrossRef]
- Ling-Ling, S.; Qing, T.; Guang, L.; Zong-Xing, L.; Xiaoying, L.; Juan, G.; Yuchen, L.; Qiao, C.; Yue, Z. Variation in characteristics of leaf functional traits of alpine vegetation in the Three-River Headwaters Region, China. Ecol. Indic. 2022, 145, 109557. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S., III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1999; Volume 30, pp. 1–67. [Google Scholar]
- Li, C.; Yu, H.; Xu, Y.; Zhu, W.; Wang, P.; Huang, J. Close linkages between leaf functional traits and soil and leaf C:N:P stoichiometry under altered precipitation in a desert steppe in northwestern China. Plant Ecol. 2022, 223, 407–421. [Google Scholar] [CrossRef]
- Tor-ngern, P.; Chart-asa, C.; Chanthorn, W.; Rodtassana, C.; Yampum, S.; Unawong, W.; Nathalang, A.; Brockelman, W.; Srinoppawan, K.; Chen, Y.; et al. Variation of leaf-level gas exchange rates and leaf functional traits of dominant trees across three successional stages in a Southeast Asian tropical forest. For. Ecol. Manag. 2021, 489, 119101. [Google Scholar] [CrossRef]
- Chanthorn, W.; Hartig, F.; Brockelman, W.Y. Structure and community composition in a tropical forest suggest a change of ecological processes during stand development. For. Ecol. Manag. 2017, 404, 100–107. [Google Scholar] [CrossRef]
- Zhang, H.; John, R.; Peng, Z.; Yuan, J.; Chu, C.; Du, G.; Zhou, S. The Relationship between Species Richness and Evenness in Plant Communities along a Successional Gradient: A Study from Sub-Alpine Meadows of the Eastern Qinghai-Tibetan Plateau, China. PLoS ONE 2012, 7, e49024. [Google Scholar] [CrossRef]
- Ordoñez, J.C.; Van Bodegom, P.M.; Witte, J.-P.M.; Wright, I.J.; Reich, P.B.; Aerts, R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 2009, 18, 137–149. [Google Scholar] [CrossRef]
- De la Riva, E.; Villar, R.; Perez-Ramos, I.; Quero, J.; Matías, L.; Poorter, L.; Marañón, T. Relationships between leaf mass per area and nutrient concentrations in 98 Mediterranean woody species are determined by phylogeny, habitat and leaf habit. Trees 2018, 32, 497–510. [Google Scholar] [CrossRef]
- Zirbel, C.R.; Grman, E.; Bassett, T.; Brudvig, L.A. Landscape context explains ecosystem multifunctionality in restored grasslands better than plant diversity. Ecology 2019, 100, e02634. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Romero-Pérez, I.E.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef]
- Buzzard, V.; Hulshof, C.M.; Birt, T.; Violle, C.; Enquist, B.J. Re-growing a tropical dry forest: Functional plant trait composition and community assembly during succession. Funct. Ecol. 2016, 30, 1006–1013. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Bongers, F.; Poorter, L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Guariguata, M.R.; Ostertag, R. Neotropical secondary forest succession: Changes in structural and functional characteristics. For. Ecol. Manag. 2001, 148, 185–206. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Lohbeck, M.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Poorter, L.; Bongers, F. Functional trait strategies of trees in dry and wet tropical forests are similar but differ in their consequences for succession. PLoS ONE 2015, 10, e0123741. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L.; Rozendaal, D.M.A.; Bongers, F.; de Almeida-Cortez, J.S.; Almeyda Zambrano, A.M.; Álvarez, F.S.; Andrade, J.L.; Villa, L.F.A.; Balvanera, P.; Becknell, J.M.; et al. Wet and dry tropical forests show opposite successional pathways in wood density but converge over time. Nat. Ecol. Evol. 2019, 3, 928–934. [Google Scholar] [CrossRef]
- Faccion, G.; Alves, A.M.; do Espírito-Santo, M.M.; Silva, J.O.; Sanchez-Azofeifa, A.; Ferreira, K.F. Intra- and interspecific variations on plant functional traits along a successional gradient in a Brazilian tropical dry forest. Flora 2021, 279, 151815. [Google Scholar] [CrossRef]
- Vogel, A.; Manning, P.; Cadotte, M.W.; Cowles, J.; Isbell, F.; Jousset, A.L.C.; Kimmel, K.; Meyer, S.T.; Reich, P.B.; Roscher, C.; et al. Chapter Three—Lost in trait space: Species-poor communities are inflexible in properties that drive ecosystem functioning. In Advances in Ecological Research; Eisenhauer, N., Bohan, D.A., Dumbrell, A.J., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 61, pp. 91–131. [Google Scholar]
- Wu, Z.Y. Vegetation of China; Science Press: Beijing, China, 1980; pp. 363–397. [Google Scholar]
- Peng, J.B.; Liu, T.; Chen, J.X.; Li, Z.Y.; Ling, Y.A.; De Wulf, A.; De Maeyer, P. The conflicts of agricultural water supply and demand under climate change in a typical arid land watershed of Central Asia. J. Hydrol. Reg. Stud. 2023, 47, 101384. [Google Scholar] [CrossRef]
- Chen, J.X.; Zhang, J.H.; Peng, J.B.; Zou, L.; Fan, Y.J.; Yang, F.R.; Hu, Z.W. Alp-valley and elevation effects on the reference evapotranspiration and the dominant climate controls in Red River Basin, China: Insights from geographical differentiation. J. Hydrol. 2023, 620, 129397. [Google Scholar] [CrossRef]
- Zhang, J.; Zuo, X.; Lv, P.; Zhao, S.; Zhao, X. Plant functional trait response to habitat change and grazing in a semiarid grassland: Unravelling species turnover and intraspecific variation effects. Pol. J. Ecol. 2019, 67, 62–74. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Poorter, L.; Bongers, F. Environmental changes during secondary succession in a tropical dry forest in Mexico. J. Trop. Ecol. 2011, 27, 477–489. [Google Scholar] [CrossRef]
- Cao, M.; Zou, X.; Warren, M.; Zhu, H. Tropical Forests of Xishuangbanna, China1. Biotropica 2006, 38, 306–309. [Google Scholar] [CrossRef]
- Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/ (accessed on 9 May 2022).
- Chianucci, F.; Cutini, A. Estimation of canopy properties in deciduous forests with digital hemispherical and cover photography. Agric. For. Meteorol. 2013, 168, 130–139. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Steege, H.t.; Morgan, H.D.; Heijden, M.G.A.V.D.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Lenza, E.; Klink, C.A. Phenological behavior of woody species in a “cerrado” sensu stricto of Brasília, DF. Braz. J. Bot. 2006, 29, 627–638. [Google Scholar] [CrossRef]
- Araújo, I.; Morandi, P.S.; Müller, A.O.; Mariano, L.H.; Alvarez, F.; da Silva, I.V.; Marimon Junior, B.H.; Marimon, B.S. Leaf functional traits and monodominance in Southern Amazonia tropical forests. Plant Ecol. 2022, 223, 185–200. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Chemical Analysis; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Zhao, N.; Yu, G.; He, N.; Wang, Q.; Guo, D.; Zhang, X.; Wang, R.; Xu, Z.; Jiao, C.; Li, N.; et al. Coordinated pattern of multi-element variability in leaves and roots across Chinese forest biomes. Glob. Ecol. Biogeogr. 2016, 25, 359–367. [Google Scholar] [CrossRef]
- Maeshiro, R.; Kusumoto, B.; Fujii, S.; Shiono, T.; Kubota, Y. Using tree functional diversity to evaluate management impacts in a subtropical forest. Ecosphere 2013, 4, art70. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for Spatial Analysis in Macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Liu, S.; He, F.; Letcher, S.G. Recovery of woody plant diversity in tropical rain forests in southern China after logging and shifting cultivation. Biol. Conserv. 2012, 145, 225–233. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Aiba, S.-I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental correlates of tree biomass, basal area, wood specific gravity and stem density gradients in Borneo’s tropical forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Sattistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Feng, Y.-L.; Fu, G.-L.; Zheng, Y.-L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners. Planta 2008, 228, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W.A.; Geiger, E.L.; Gotsch, S.G.; Rossatto, D.R.; Silva, L.C.R.; Lau, O.L.; Haridasan, M.; Franco, A.C. Ecological thresholds at the savanna-forest boundary: How plant traits, resources and fire govern the distribution of tropical biomes. Ecol. Lett. 2012, 15, 759–768. [Google Scholar] [CrossRef]
- Díaz, S.; Hodgson, J.G.; Thompson, K.; Cabido, M.; Cornelissen, J.H.C.; Jalili, A.; Montserrat-Martí, G.; Grime, J.P.; Zarrinkamar, F.; Asri, Y.; et al. The plant traits that drive ecosystems: Evidence from three continents. J. Veg. Sci. 2004, 15, 295–304. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Orthen, B.; Franco, A.C. Constraints to seedling success of savanna and forest trees across the savanna-forest boundary. Oecologia 2004, 140, 252–260. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E.; Schwilk, D.W. Flammability as an ecological and evolutionary driver. J. Ecol. 2017, 105, 289–297. [Google Scholar] [CrossRef]
- Takashima, T.; Hikosaka, K.; Hirose, T. Photosynthesis or persistence: Nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant Cell Environ. 2004, 27, 1047–1054. [Google Scholar] [CrossRef]
- Kitajima, K.; Llorens, A.M.; Stefanescu, C.; Timchenko, M.V.; Lucas, P.W.; Wright, S.J. How cellulose-based leaf toughness and lamina density contribute to long leaf lifespans of shade-tolerant species. New Phytol. 2012, 195, 640–652. [Google Scholar] [CrossRef]
- Cunningham, S.A.; Summerhayes, B.; Westoby, M. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecol. Monogr. 1999, 69, 569–588. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Derroire, G.; Powers, J.S.; Hulshof, C.M.; Cárdenas Varela, L.E.; Healey, J.R. Contrasting patterns of leaf trait variation among and within species during tropical dry forest succession in Costa Rica. Sci. Rep. 2018, 8, 285. [Google Scholar] [CrossRef]
- Larjavaara, M.; Muller-Landau, H.C. Rethinking the value of high wood density. Funct. Ecol. 2010, 24, 701–705. [Google Scholar] [CrossRef]
- Plourde, B.T.; Boukili, V.K.; Chazdon, R.L. Radial changes in wood specific gravity of tropical trees: Inter- and intraspecific variation during secondary succession. Funct. Ecol. 2015, 29, 111–120. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, D.S.; Reich, P.B. Photosynthesis and Leaf Nitrogen in Five Amazonian Tree Species During Early Secondary Succession. Ecology 1996, 77, 581–594. [Google Scholar] [CrossRef]
- Raaimakers, D.; Boot, R.G.A.; Dijkstra, P.; Pot, S. Photosynthetic rates in relation to leaf phosphorus content in pioneer versus climax tropical rainforest trees. Oecologia 1995, 102, 120–125. [Google Scholar] [CrossRef]
- Pacala, S.W.; Rees, M.A. Models Suggesting Field Experiments to Test Two Hypotheses Explaining Successional Diversity. Am. Nat. 1998, 152, 729–737. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Aide, T.M.; Erickson, D.L.; Forero, M.; Kress, W.J.; Swenson, N.G.; Zimmerman, J.K. Functional convergence and phylogenetic divergence during secondary succession of subtropical wet forests in Puerto Rico. J. Veg. Sci. 2016, 27, 283–294. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R. Interactions of drought and shade effects on seedlings of four Quercus species: Physiological and structural leaf responses. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Rada, F.; Sarmiento, L.; García-Varela, S. Plant functional traits along an old-field succession in the high tropical andes. Acta Oecologica 2021, 111, 103738. [Google Scholar] [CrossRef]
- Gulías, J.; Flexas, J.; Mus, M.; Cifre, J.; Lefi, E.; Medrano, H. Relationship between maximum leaf photosynthesis, nitrogen content and specific leaf area in Balearic endemic and non-endemic Mediterranean species. Ann. Bot. 2003, 92, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, D.; Leuschner, C.; Bohman, K.; Hagemeier, M.; Juhrbandt, J.; Tjitrosemito, S. Leaf gas exchange of trees in old-growth and young secondary forest stands in Sulawesi, Indonesia. Trees Struct. Funct. 2006, 20, 278–285. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, L.; Wen, D. Photosynthesis of subtropical forest species from different successional status in relation to foliar nutrients and phosphorus fractions. Sci. Rep. 2018, 8, 10455. [Google Scholar] [CrossRef]
- Asefa, M.; Cao, M.; Zhang, G.; Ci, X.; Li, J.; Yang, J. Environmental filtering structures tree functional traits combination and lineages across space in tropical tree assemblages. Sci. Rep. 2017, 7, 132. [Google Scholar] [CrossRef]
- Quisehuatl-Medina, A.; Averett, J.P.; Endress, B.A.; Lopez-Toledo, L. Removal of cattle accelerates tropical dry forest succession in Northwestern Mexico. Biotropica 2020, 52, 457–469. [Google Scholar] [CrossRef]
- Maza-Villalobos, S.; García-Ramírez, P.; Endress, B.A.; Lopez-Toledo, L. Plant functional traits under cattle grazing and fallow age scenarios in a tropical dry forest of Northwestern Mexico. Basic Appl. Ecol. 2022, 64, 30–44. [Google Scholar] [CrossRef]
- Ayala-Orozco, B.; Gavito, M.E.; Mora, F.; Siddique, I.; Balvanera, P.; Jaramillo, V.J.; Cotler, H.; Romero-Duque, L.P.; Martínez-Meyer, E. Resilience of Soil Properties to Land-Use Change in a Tropical Dry Forest Ecosystem. Land Degrad. Dev. 2018, 29, 315–325. [Google Scholar] [CrossRef]
- Gavito, M.E.; Sandoval-Pérez, A.L.; del Castillo, K.; Cohen-Salgado, D.; Colarte-Avilés, M.E.; Mora, F.; Santibáñez-Rentería, A.; Siddique, I.; Urquijo-Ramos, C. Resilience of soil nutrient availability and organic matter decomposition to hurricane impact in a tropical dry forest ecosystem. For. Ecol. Manag. 2018, 426, 81–90. [Google Scholar] [CrossRef]
- Campo, J.; Maass, M.; Jaramillo, V.J.; Martínez-Yrízar, A.; Sarukhán, J. Phosphorus cycling in a Mexican tropical dry forest ecosystem. Biogeochemistry 2001, 53, 161–179. [Google Scholar] [CrossRef]
- Niklas, K.L. Differences between Acer saccharum Leaves from Open and Wind-Protected Sites. Ann. Bot. 1996, 78, 61–66. [Google Scholar] [CrossRef]
- Ohdo, T.; Takahashi, K. Plantspecies richness and community assembly along gradients of elevation and soil nitrogen availability. Aob Plants 2020, 12, plaa014. [Google Scholar] [CrossRef] [PubMed]
- Llambí, L.D.; Fontaine, M.; Rada, F.; Saugier, B.; Sarmiento, L. Ecophysiology of Dominant Plant Species during Old-Field Succession in a High Tropical Andean Ecosystem. Arct. Antarct. Alp. Res. 2003, 35, 447–453. [Google Scholar] [CrossRef]
- Llambí, L.; Sarmiento, L. Biomasa microbiana y otros parámetros edáficos en una sucesión secundaria en el páramo. Ecotropicos 1998, 11, 1–14. [Google Scholar]
- Monasterio, M.; Sarmiento, L. Adaptive radiation of Espeletia in the cold andean tropics. Trends Ecol. Evol. 1991, 6, 387–391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Forest Types | Indicator Species | Indicator Values | p |
|---|---|---|---|
| Secondary Forests | Saprosma ternatum Hook.f. | 32.6 | 0.0382 |
| Baccaurea ramiflora Lour. | 27.8 | 0.3955 | |
| Garcinia yunnanensis Hu | 23.2 | 0.0684 | |
| Ficus langkokensis Drake | 23 | 0.1336 | |
| Dysoxylum excelsum BI. | 21.7 | 0.2144 | |
| Antidesma montanum BI. | 20.8 | 0.0442 | |
| Dichapetalum gelonioides (Roxb.) Engl. | 18.8 | 0.4733 | |
| Beilschmiedia purpurascens H.W.Li | 18.1 | 0.4011 | |
| Polyalthia cheliensis Hu | 16.9 | 0.3311 | |
| Cinnamomum tenuipilum Kosterm. | 16.6 | 0.0234 | |
| Aidia yunnanensis (Hutchins.) Yamazaki | 16.3 | 0.5047 | |
| Ecotone Forests | Ardisia tenera Mez | 38.9 | 0.0496 |
| Litsea pierrei Lec. var. szemois Liou | 38 | 0.0016 | |
| Syzygium polypetaloideum Merr. et Perry | 37.2 | 0.0016 | |
| Castanopsis echinocarpa Miq. | 36.8 | 0.0188 | |
| Memecylon polyanthum H.L.Li | 35.8 | 0.0022 | |
| Litsea verticillata Hance | 35.8 | 0.0014 | |
| Garcinia cowa Roxb. | 35.7 | 0.0658 | |
| Canarium album (Lour.) Raeusch. | 35.6 | 0.0174 | |
| Machilus tenuipila H. W. Li | 35 | 0.1196 | |
| Old-growth Forests | Litsea pittosporifolia Yang et P. H. Huang | 34.4 | 0.0086 |
| Phoebe lanceolata (Wall. ex Nees) Nees | 27.6 | 0.7285 | |
| Pittosporopsis kerrii Craib | 27.1 | 0.94 | |
| Parashorea chinensis Wang Hsie | 23.4 | 0.1416 | |
| Knema linifolia (Roxb.) Warb. | 21.4 | 0.6629 | |
| Schima crenata Korth. | 21.4 | 0.028 | |
| Barringtonia racemosa (L.) Spreng. | 18.6 | 0.4775 | |
| Knema erratica (Hook. f. et Thoms.) J. Sincl. | 18.5 | 0.4813 | |
| Gironniera subaequalis Planch. | 18.3 | 0.0882 |
| Environmental Factors | Functional Traits | |||||
|---|---|---|---|---|---|---|
| SLA | LNC | LPC | LKC | LDMC | LCC | |
| Models | #31,682 | #11,259 | #15,354 | #15,354 | #15,989 | #7044 |
| r² | 0.206 | 0.364 | 0.452 | 0.417 | 0.229 | 0.341 |
| AICc | −253.825 | −220.177 | −162.143 | −85.444 | −302.118 | −404.049 |
| Rtime | 0.354 | 0.627 | 0.837 | 0.644 | −0.458 | −0.799 |
| Aspect | −0.256 | −0.302 | −0.238 | −0.225 | 0.275 | −0.808 |
| Slope | −0.312 | 0.515 | 0.298 | 0.301 | 0.229 | −0.378 |
| Litter | 0.248 | 0.534 | 0.343 | 0.586 | −0.248 | −0.293 |
| Elevation | −0.616 | −0.859 | −0.928 | −0.761 | −0.251 | 0.964 |
| SWC | −0.245 | 0.352 | 0.238 | 0.368 | −0.254 | 0.232 |
| pH | 0.279 | −0.249 | −0.293 | −0.243 | 0.718 | −0.258 |
| SOM | −0.351 | −0.252 | −0.387 | −0.281 | 0.771 | 0.243 |
| TN | 0.398 | −0.234 | −0.548 | −0.445 | 0.392 | −0.696 |
| TP | 0.739 | 0.238 | 0.378 | 0.237 | −0.433 | −0.416 |
| TK | −0.26 | 0.34 | 0.607 | 0.33 | −0.369 | −0.291 |
| AN | 0.277 | 0.851 | 0.857 | 0.789 | −0.488 | −0.307 |
| AP | 0.329 | −0.283 | 0.318 | 0.253 | −0.515 | −0.231 |
| AK | 0.287 | 0.231 | −0.376 | −0.227 | 0.401 | 0.238 |
| COPE | 0.362 | −0.735 | −0.711 | −0.964 | −0.243 | 0.394 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, F.; Xiao, L.; Huang, J.; Luo, H.; Zang, R. Changes in Leaf Functional Traits Driven by Environmental Filtration in Different Monsoon Tropical Forest Types. Forests 2023, 14, 2101. https://doi.org/10.3390/f14102101
Deng F, Xiao L, Huang J, Luo H, Zang R. Changes in Leaf Functional Traits Driven by Environmental Filtration in Different Monsoon Tropical Forest Types. Forests. 2023; 14(10):2101. https://doi.org/10.3390/f14102101
Chicago/Turabian StyleDeng, Fuying, Lijun Xiao, Jing Huang, Honghao Luo, and Runguo Zang. 2023. "Changes in Leaf Functional Traits Driven by Environmental Filtration in Different Monsoon Tropical Forest Types" Forests 14, no. 10: 2101. https://doi.org/10.3390/f14102101