
Effects of Truffle Inoculation on Root Physiology and Mycorrhizosphere Microbial Communities of Carya illinoinensis Seedlings
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seedling and Tuber Inoculation of Pecan
2.2. Sample Collection
2.3. Colonization Detection and Infestation Rate Statistics of Tubercle Bacteria
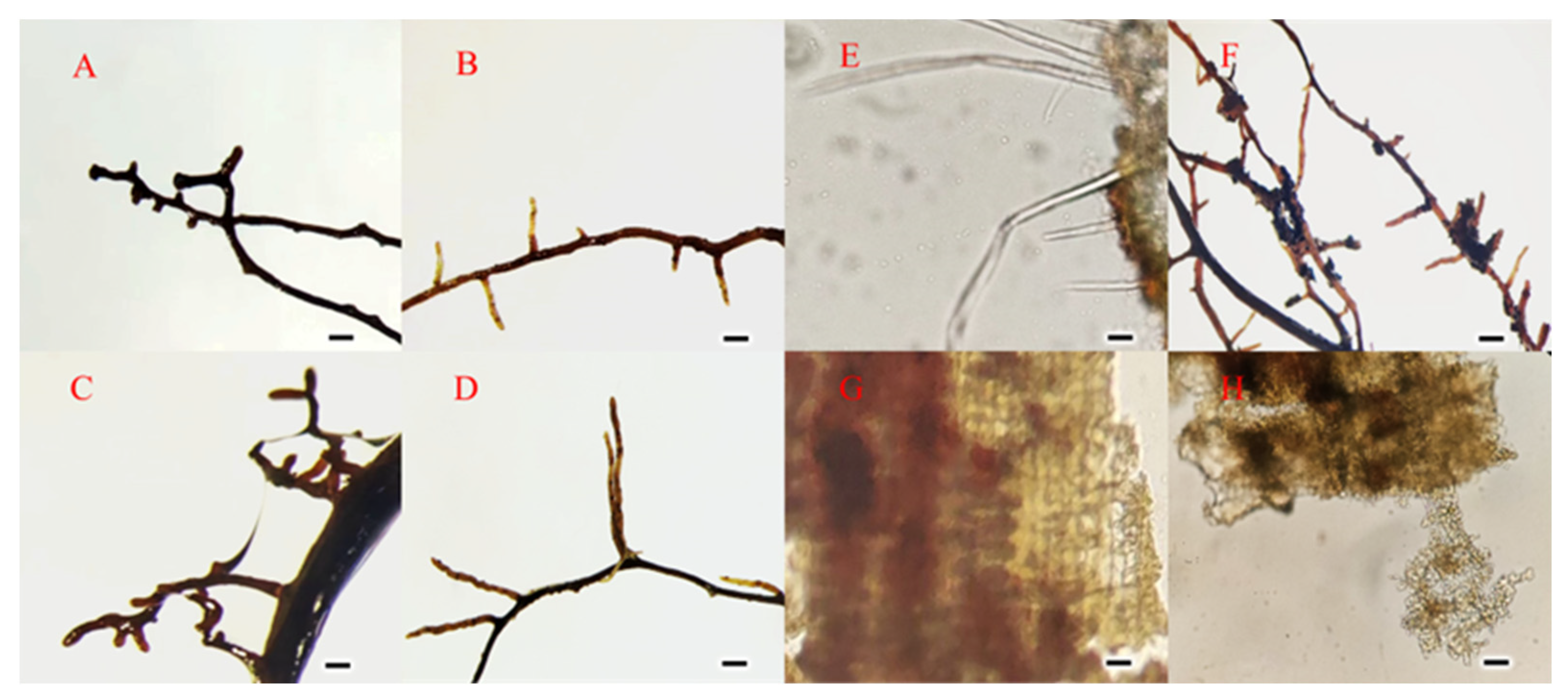
2.3.1. Morphological Detection of ECM
2.3.2. Molecular Identification of Mycorrhizal Fungi
2.3.3. Calculation of Infestation Rate
2.4. Determination of Sample Indicators
2.4.1. Steps for the Determination of Microbial Indicators
2.4.2. Measurement Steps of Soil Physicochemical Indexes
2.4.3. Steps for the Determination of Physiological Indices of the Root System
2.5. Data Analysis and Visualization
3. Results
3.1. Morphological Characteristics of Ectomycorrhizae
3.2. Physiological Characteristics of Seedling Root System
3.3. Characterization of Mycorrhizosphere Soil Chemical Properties
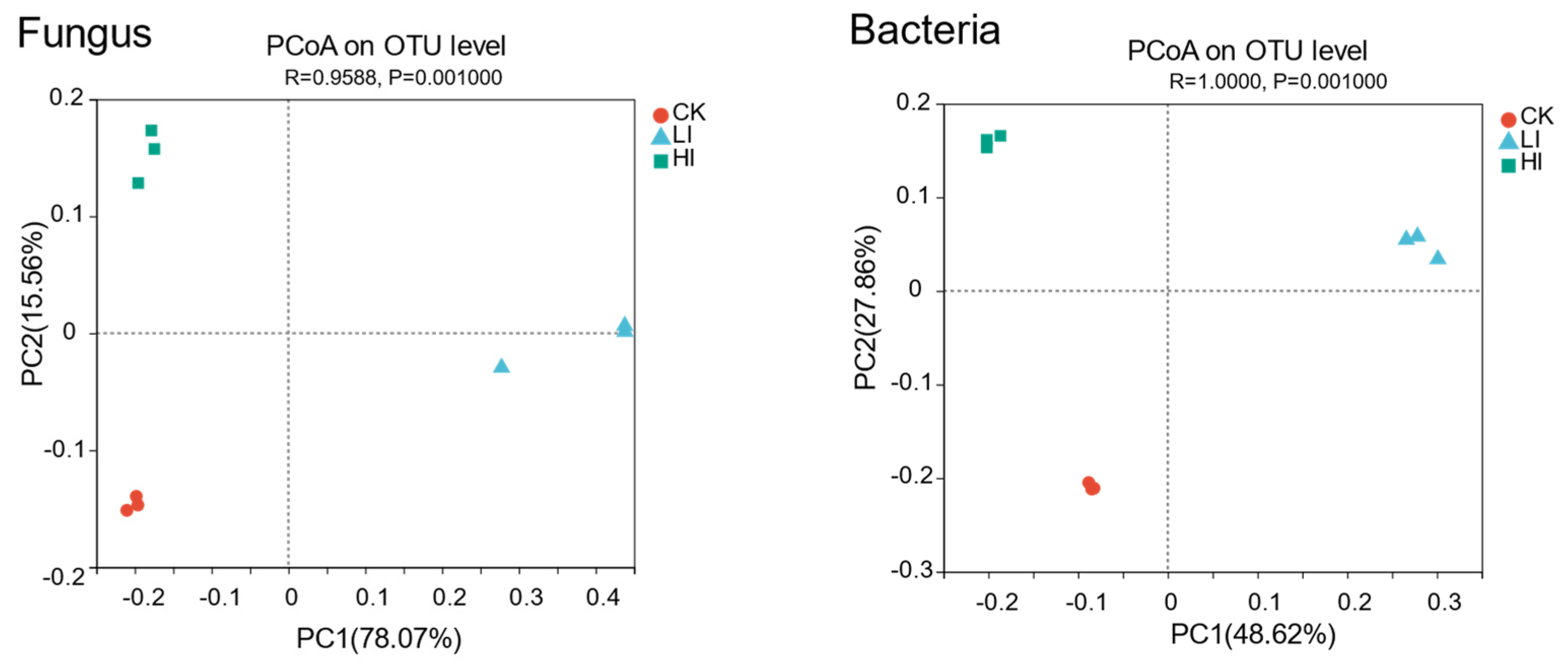
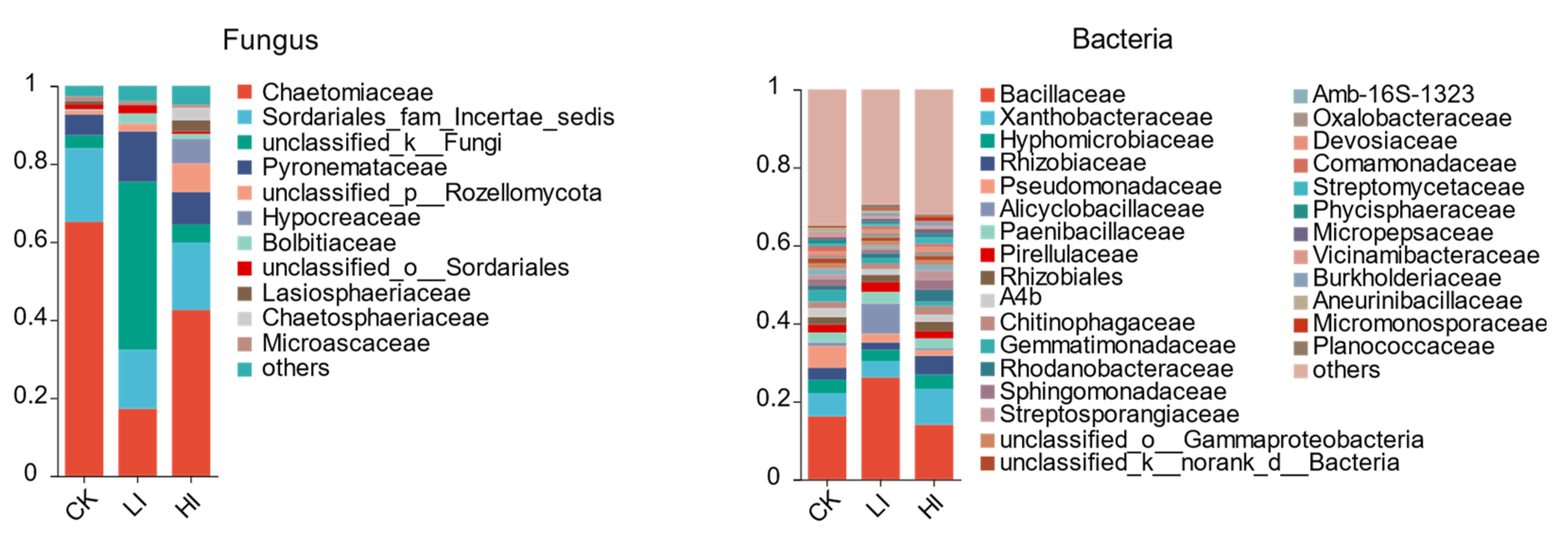
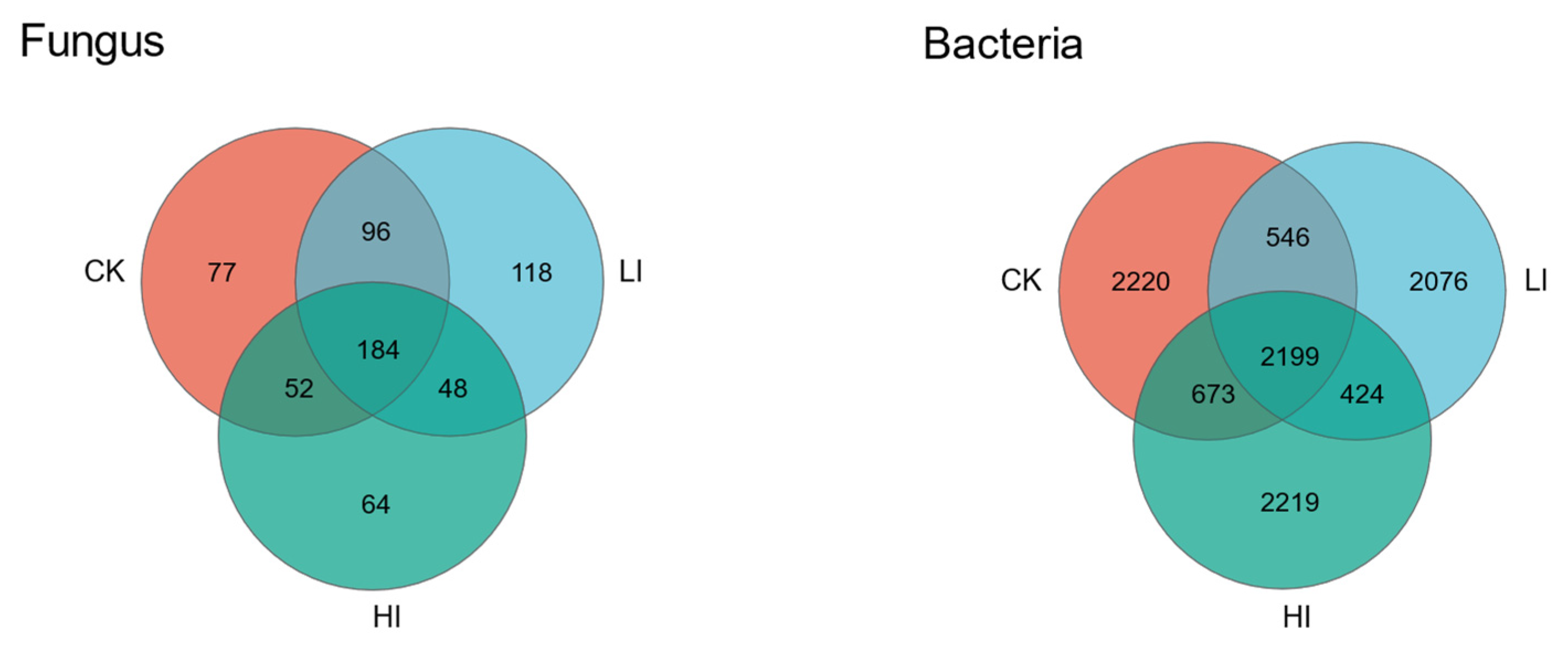
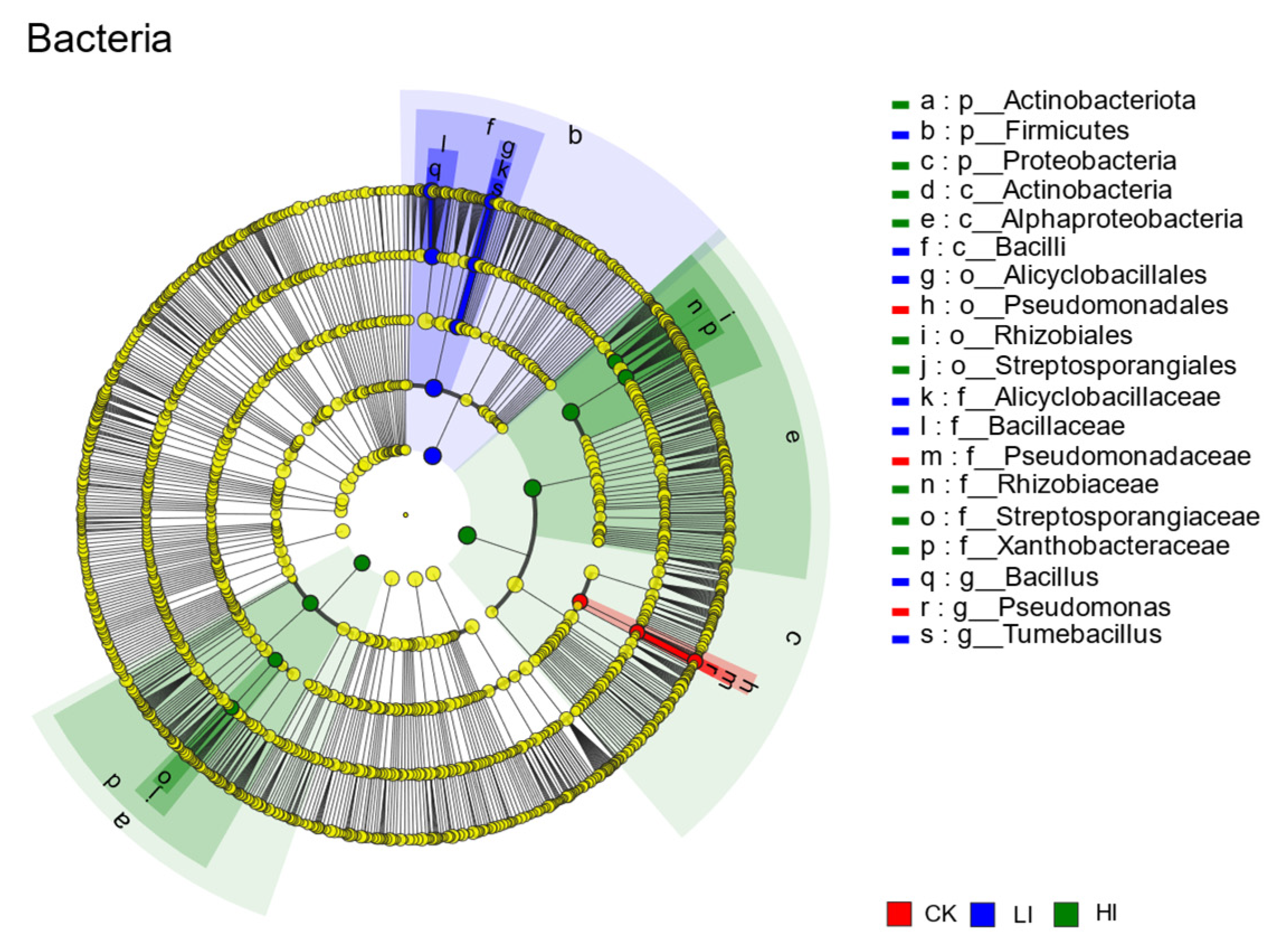
3.4. Characterization of Mycorrhizosphere Soil Microbial Communities
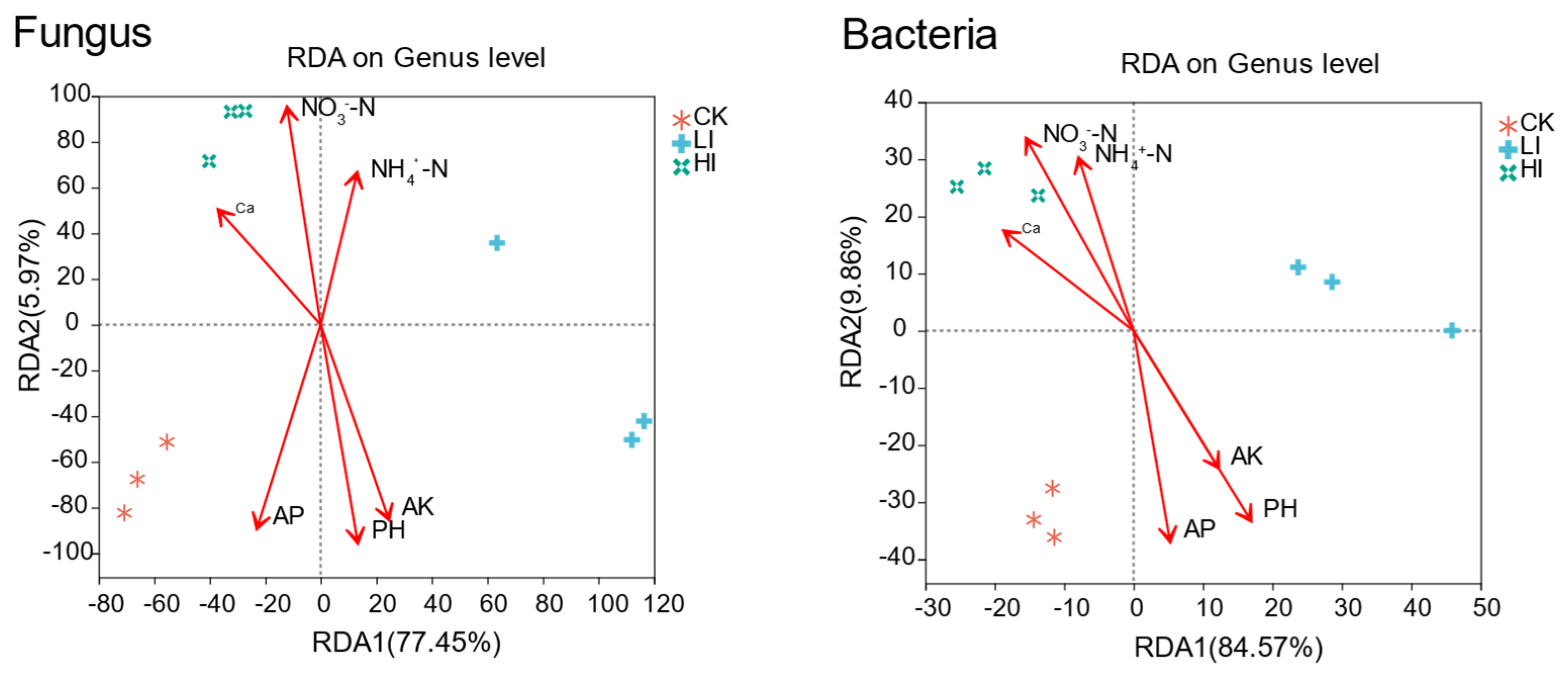
3.5. Association of Mycorrhizosphere Soil Microbial Community Composition with Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Phong, W.N.; Gibberd, M.R.; Payne, A.D.; Dykes, G.A.; Coorey, R. Methods used for extraction of plant volatiles have potential to preserve truffle aroma: A review. Compr. Rev. Food. Sci. Food Saf. 2022, 21, 1677–1701. [Google Scholar] [CrossRef] [PubMed]
- Rubini, A.; Riccioni, C.; Belfiori, B.; Paolocci, F. Impact of the competition between mating types on the cultivation of Tuber melanosporum: Romeo and Juliet and the matter of space and time. Mycorrhiza 2014, 24, S19–S27. [Google Scholar] [CrossRef]
- Rubini, A.; Belfiori, B.; Riccioni, C.; Arcioni, S.; Martin, F.; Paolocci, F. Tuber melanosporum: Mating type distribution in a natural plantation and dynamics of strains of different mating types on the roots of nursery-inoculated host plants. New Phytol. 2011, 189, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Murat, C. Forty years of inoculating seedlings with truffle fungi: Past and future perspectives. Mycorrhiza 2015, 25, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Benucci, G.M.N.; Raggi, L.; Albertini, E.; Grebenc, T.; Bencivenga, M.; Falcinelli, M.; Di Massimo, G. Ectomycorrhizal communities in a productive Tuber aestivum Vittad. orchard: Composition, host influence and species replacement. FEMS Microbiol. Ecol. 2011, 76, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Bonito, G.; Smith, M.E.; Nowak, M.; Healy, R.A.; Guevara, G.; Cázares, E.; Kinoshita, A.; Nouhra, E.R.; Domínguez, L.S.; Tedersoo, L.; et al. Historical Biogeography and Diversification of Truffles in the Tuberaceae and Their Newly Identified Southern Hemisphere Sister Lineage. PLoS ONE 2013, 8, e52765. [Google Scholar] [CrossRef]
- Iotti, M.; Piattoni, F.; Leonardi, P.; Hall, I.R.; Zambonelli, A. First evidence for truffle production from plants inoculated with mycelial pure cultures. Mycorrhiza 2016, 26, 793–798. [Google Scholar] [CrossRef]
- Bonfante, P.; Anca, I.A. Plants, Mycorrhizal Fungi, and Bacteria: A Network of Interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef]
- Palacios, Y.M.; Winfrey, B.K. Three mechanisms of mycorrhizae that may improve stormwater biofilter performance. Ecol. Eng. 2021, 159, 8. [Google Scholar] [CrossRef]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Dominguez, J.A.; Martin, A.; Anriquez, A.; Albanesi, A. The combined effects of Pseudomonas fluorescens and Tuber melanosporum on the quality of Pinus halepensis seedlings. Mycorrhiza 2012, 22, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Le Tacon, F.; Zeller, B.; Plain, C.; Hossann, C.; Brechet, C.; Robin, C. Carbon Transfer from the Host to Tuber melanosporum Mycorrhizas and Ascocarps Followed Using a C-13 Pulse-Labeling Technique. PLoS ONE 2013, 8, e64626. [Google Scholar] [CrossRef]
- Bonito, G.; Brenneman, T.; Vilgalys, R. Ectomycorrhizal fungal diversity in orchards of cultivated pecan (Carya illinoinensis; Juglandaceae). Mycorrhiza 2011, 21, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Marozzi, G.; Sanchez, S.; Benucci, G.M.N.; Bonito, G.; Falini, L.B.; Albertini, E.; Donnini, D. Mycorrhization of pecan (Carya illinoinensis) with black truffles: Tuber melanosporum and Tuber brumale. Mycorrhiza 2017, 27, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Cejka, T.; Trnka, M.; Krusic, P.J.; Stobbe, U.; Oliach, D.; Vaclavik, T.; Tegel, W.; Buntgen, U. Predicted climate change will increase the truffle cultivation potential in central Europe. Sci. Rep. 2020, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Liu, Y.J.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Rigamonte, T.A.; Pylro, V.S.; Duarte, G.F. The Role of Mycorrhization Helper Bacteria in the Establishment and Action of Ectomycorrhizae Associations. Braz. J. Microbiol. 2010, 41, 832–840. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Wu, X.Q.; Hou, L.L.; Sheng, J.M.; Ren, J.H.; Zheng, L.; Chen, D.; Ye, J.R. Effects of ectomycorrhizal fungus Boletus edulis and mycorrhiza helper Bacillus cereus on the growth and nutrient uptake by Pinus thunbergii. Biol. Fertil. Soils 2012, 48, 385–391. [Google Scholar] [CrossRef]
- Mediavilla, O.; Olaizola, J.; Santos-del-Blanco, L.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Mycorrhization between Cistus ladanifer L. and Boletus edulis Bull is enhanced by the mycorrhiza helper bacteria Pseudomonas fluorescens Migula. Mycorrhiza 2016, 26, 161–168. [Google Scholar] [CrossRef]
- Sabella, E.; Nutricati, E.; Aprile, A.; Miceli, A.; Sorce, C.; Lorenzi, R.; De Bellis, L. Arthrinium phaeospermum isolated from Tuber borchii ascomata: The first evidence for a “Mycorrhization Helper Fungus”? Mycol. Prog. 2015, 14, 9. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Li, Y.H.; Xu, W.H.; Chen, W.J.; Hu, Y.L.; Wang, Z.G. Different genotypes regulate the microbial community structure in the soybean rhizosphere. J. Integr. Agric. 2023, 22, 585–597. [Google Scholar] [CrossRef]
- Kang, Z.J.; Li, X.L.; Li, Y.; Ye, L.; Zhang, B.; Zhang, X.P.; Penttinen, P.; Gu, Y.F. Black Truffles Affect Quercus aliena Physiology and Root-Associated nirK- and nirS-Type Denitrifying Bacterial Communities in the Initial Stage of Inoculation. Front. Microbiol. 2022, 13, 11. [Google Scholar] [CrossRef]
- Agerer, R. Fungal relationships and structural identity of their ectomycorrhizae. Mycol. Prog. 2006, 5, 67–107. [Google Scholar] [CrossRef]
- Donnini, D.; Benucci, G.M.N.; Bencivenga, M.; Baciarelli-Falini, L. Quality assessment of truffle-inoculated seedlings in Italy: Proposing revised parameters for certification. For. Syst. 2014, 23, 385–393. [Google Scholar] [CrossRef]
- Doane, T.A.; Horwath, W.R. Spectrophotometric determination of nitrate with a single reagent. Anal. Lett. 2003, 36, 2713–2722. [Google Scholar] [CrossRef]
- Liu, J.P.; Tang, Y.J.; Bao, J.S.; Wang, H.K.; Peng, F.R.; Chen, M.Y.; Tan, P.P. Pecan plantation age influences the structures, ecological networks, and functions of soil microbial communities. Land Degrad. Dev. 2022, 33, 3294–3309. [Google Scholar] [CrossRef]
- Moore, T.R.; Bubier, J.L.; Frolking, S.E.; Lafleur, P.M.; Roulet, N.T. Plant biomass and production and CO2 exchange in an ombrotrophic bog. J. Ecol. 2010, 90, 25–36. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, G.; Bian, X.; Zhao, Q. Effects of root interaction and nitrogen fertilization on the chlorophyll content, root activity, photosynthetic characteristics of intercropped soybean and microbial quantity in the rhizosphere. Plant Soil Environ. 2013, 59, 80–88. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases: Anti-Versus Pro- Oxidants? Anti-Cancer Agents Med. Chem. 2011, 11, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glockner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Benucci, G.M.N.; Bonito, G.; Falini, L.B.; Bencivenga, M. Mycorrhization of Pecan trees (Carya illinoinensis) with commercial truffle species: Tuber aestivum Vittad. and Tuber borchii Vittad. Mycorrhiza 2012, 22, 383–392. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant-microbiome interactions under a changing world: Responses, consequences and perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef]
- Xiong, W.; Song, Y.Q.; Yang, K.M.; Gu, Y.; Wei, Z.; Kowalchuk, G.A.; Xu, Y.C.; Jousset, A.; Shen, Q.; Geisen, S. Rhizosphere protists are key determinants of plant health. Microbiome 2020, 8, 9. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Li, Y.X.; Wei, P.F.; Sun, N.N.; Wen, X.J.; Liu, Z.K.; Li, D.Z.; Feng, Y.Q.; Zhang, X.Y. Differences Between Microbial Communities of Pinus Species Having Differing Level of Resistance to the Pine Wood Nematode. Microb. Ecol. 2022, 84, 1245–1255. [Google Scholar] [CrossRef]
- Adair, K.L.; Douglas, A.E. Making a microbiome: The many determinants of host-associated microbial community composition. Curr. Opin. Microbiol. 2017, 35, 23–29. [Google Scholar] [CrossRef]
- Kennedy, P.G.; Bruns, T.D. Priority effects determine the outcome of ectomycorrhizal competition between two Rhizopogon species colonizing Pinus muricata seedlings. New Phytol. 2005, 166, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.; Peay, K.G.; Bruns, T.D. Root tip competition among ectomycorrhizal fungi: Are priority effects a rule or an exception? Ecology 2009, 90, 2098–2107. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, J.; Xiong, C.; Li, X.L.; Chen, Z.Q.; Li, P.; Huang, W.L. Tuber indicum shapes the microbial communities of ectomycorhizosphere soil and ectomycorrhizae of an indigenous tree (Pinus armandii). PLoS ONE 2017, 12, e0175720. [Google Scholar] [CrossRef] [PubMed]
- Deveau, A.; Antony-Babu, S.; Le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Temporal changes of bacterial communities in the Tuber melanosporum ectomycorrhizosphere during ascocarp development. Mycorrhiza 2016, 26, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Antony-Babu, S.; Deveau, A.; Van Nostrand, J.D.; Zhou, J.Z.; Le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle-associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbiol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef]
- Gryndler, M.; Soukupova, L.; Hrselova, H.; Gryndlerova, H.; Borovicka, J.; Streiblova, E.; Jansa, J. A quest for indigenous truffle helper prokaryotes. Environ. Microbiol. Rep. 2013, 5, 346–352. [Google Scholar] [CrossRef]
- Barea, J.M.; Azcon, R.; Azcon-Aguilar, C. Mycorrhizosphere interactions to improve plant fitness and soil quality. Antonie Van Leeuwenhoek 2002, 81, 343–351. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Akhtar, M.S.; Futai, K. Mycorrhizae: Sustainable Agriculture and Forestry; Springer: Dordrecht, The Netherlands, 2008; pp. 228–229. [Google Scholar]
- Dietrich, M.; Montesinos-Navarro, A.; Gabriel, R.; Strasser, F.; Meier, D.V.; Mayerhofer, W.; Gorka, S.; Wiesenbauer, J.; Martin, V.; Weidinger, M.; et al. Both abundant and rare fungi colonizing Fagus sylvatica ectomycorrhizal root-tips shape associated bacterial communities. Commun. Biol. 2022, 5, 14. [Google Scholar] [CrossRef]
- Rasmann, S.; Bennett, A.; Biere, A.; Karley, A.; Guerrieri, E. Root symbionts: Powerful drivers of plant above- and belowground indirect defenses. Insect Sci. 2017, 24, 947–960. [Google Scholar] [CrossRef]
- Barto, E.K.; Weidenhamer, J.D.; Cipollini, D.; Rillig, M.C. Fungal superhighways: Do common mycorrhizal networks enhance below ground communication? Trends Plant Sci. 2012, 17, 633–637. [Google Scholar] [CrossRef]
- Gao, P.; Guo, Y.N.; Li, Y.D.; Duan, T.Y. Effects of dual inoculation of AMF and rhizobium on alfalfa (Medicago sativa) root rot caused by Microdochium tabacinum. Austral. Plant Pathol. 2018, 47, 195–203. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef] [PubMed]
- Pavic, A.; Stankovic, S.; Marjanovic, Z. Biochemical Characterization of a Sphingomonad Isolate from the Ascocarp of White Truffle (Tuber Magnatum Pico). Arch. Biol. Sci. 2011, 63, 697–704. [Google Scholar] [CrossRef]
- Becquer, A.; Guerrero-Galan, C.; Eibensteiner, J.L.; Houdinet, G.; Bucking, H.; Zimmermann, S.D.; Garcia, K. The ectomycorrhizal contribution to tree nutrition. In Molecular Physiology and Biotechnology of Trees; Canovas, F.M., Ed.; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2019; pp. 77–126. [Google Scholar]
- Liu, S.B.; He, F.K.; Kuzyakov, Y.; Xiao, H.X.; Hoang, D.T.T.; Pu, S.Y.; Razavi, B.S. Nutrients in the rhizosphere: A meta-analysis of content, availability, and influencing factors. Sci. Total Environ. 2022, 826, 14. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, eaba0196. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Infection Rate | Seedling Index | Root Activity mg TTF/(g·h) | SOD Activity (U/mg·prot) | POD Activity (U/mg·prot) |
|---|---|---|---|---|---|
| CK | 0 | 11.80 ± 5.95 a | 2.50 ± 1.35 a | 363.28 ± 44.73 a | 46.67 ± 11.55 b |
| LI | 3.28% | 9.15 ± 4.24 a | 3.05 ± 0.50 a | 369.70 ± 56.48 a | 37.78 ± 3.85 b |
| HI | 56.25% | 9.86 ± 3.29 a | 4.99 ± 2.61 a | 364.75 ± 71.68 a | 62.22 ± 3.85 a |
| p-value | 0.777 | 0.277 | 0.990 | 0.018 | |
| F-value | 0.263 | 1.600 | 0.010 | 8.455 |
| Group | PH | NH4+-N (mg/kg) | NO3−-N (mg/kg) | AP (mg/kg) | AK (mg/kg) | Ca2+ (mg/kg) |
|---|---|---|---|---|---|---|
| CK | 6.33 ± 0.01 a | 21.98 ± 5.30 b | 0.42 ± 0.05 c | 24.38 ± 1.25 a | 103.59 ± 11.29 a | 1026.16 ± 54.19 a |
| LI | 6.27 ± 0.06 b | 26.40 ± 3.82 ab | 1.53 ± 0.26 b | 20.55 ± 0.71 b | 99.80 ± 12.40 ab | 977.11 ± 68.44 a |
| HI | 6.08 ± 0.03 c | 32.73 ± 4.73 a | 5.19 ± 0.27 a | 16.96 ± 0.73 c | 77.40 ± 11.77 b | 1259.58 ± 226.07 a |
| p-value | <0.0001 | 0.077 | <0.0001 | <0.0001 | 0.069 | 0.099 |
| F-value | 141.848 | 4.044 | 383.220 | 47.458 | 4.300 | 3.491 |
| Fungal Samples | Species Richness | Species Diversity | |
|---|---|---|---|
| Classification | OTUs Observed | Shannon | Simpson |
| CK | 275.00 ± 21.52 a | 2.19 ± 0.05 b | 0.2384 ± 0.01711 b |
| LI | 308.33 ± 35.13 a | 2.37 ± 0.13 b | 0.2023 ± 0.02885 b |
| HI | 256.33 ± 20.03 a | 2.59 ± 0.96 a | 0.1517 ± 0.01504 a |
| p-value | 0.127 | 0.008 | 0.010 |
| F-value | 2.976 | 12.014 | 10.937 |
| Bacteria Samples | Species Richness | Species Diversity | |
|---|---|---|---|
| Classification | OTUs Observed | Shannon | Simpson |
| CK | 3169.00 ± 17.69 a | 6.62 ± 0.04 a | 0.0064 ± 0.00057 b |
| LI | 2965.00 ± 305.50 a | 6.15 ± 0.18 b | 0.01669 ± 0.00280 a |
| HI | 3069.67 ± 140.32 a | 6.15 ± 0.18 b | 0.00554 ± 0.00007 b |
| p-value | 0.462 | 0.003 | <0.0001 |
| F-value | 0.826 | 17.175 | 42.500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Wu, J.; Liu, J.; Tan, P.; Zhu, K.; Peng, F. Effects of Truffle Inoculation on Root Physiology and Mycorrhizosphere Microbial Communities of Carya illinoinensis Seedlings. Forests 2023, 14, 2078. https://doi.org/10.3390/f14102078
Chen H, Wu J, Liu J, Tan P, Zhu K, Peng F. Effects of Truffle Inoculation on Root Physiology and Mycorrhizosphere Microbial Communities of Carya illinoinensis Seedlings. Forests. 2023; 14(10):2078. https://doi.org/10.3390/f14102078
Chicago/Turabian StyleChen, Haoyu, Jiawei Wu, Junping Liu, Pengpeng Tan, Kaikai Zhu, and Fangren Peng. 2023. "Effects of Truffle Inoculation on Root Physiology and Mycorrhizosphere Microbial Communities of Carya illinoinensis Seedlings" Forests 14, no. 10: 2078. https://doi.org/10.3390/f14102078