
Resistance to Bark Beetle Outbreak in Norway Spruce: Population Structure Analysis and Comparative Genomic Assessment of Surviving (LTS) and Randomly Selected Reference Trees
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Plant Samples
2.2. DNA Extraction and Genotyping
2.3. Data Analysis
2.4. The Population–Genetic Structure
2.5. Candidate Gene Mining
3. Results
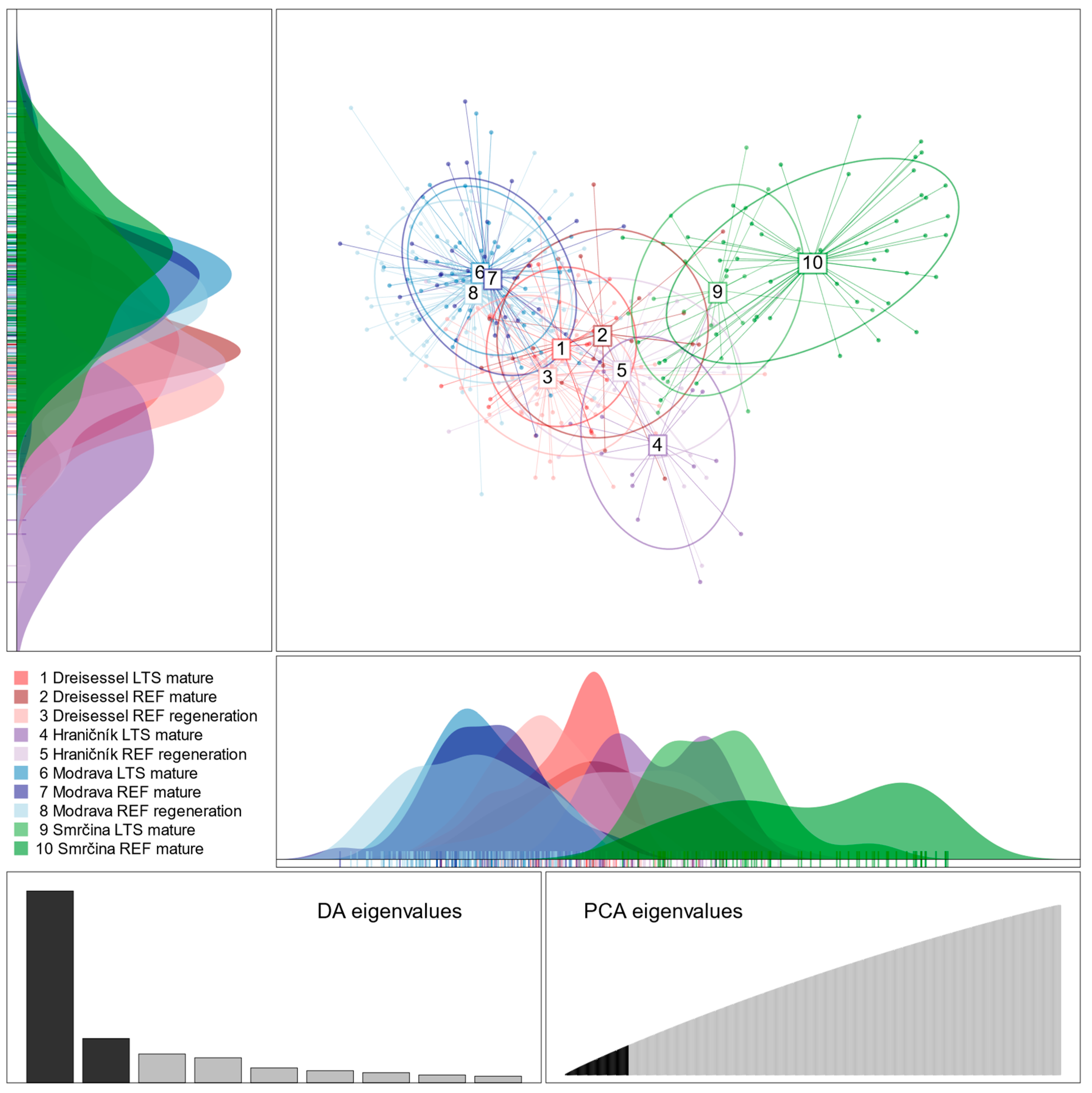
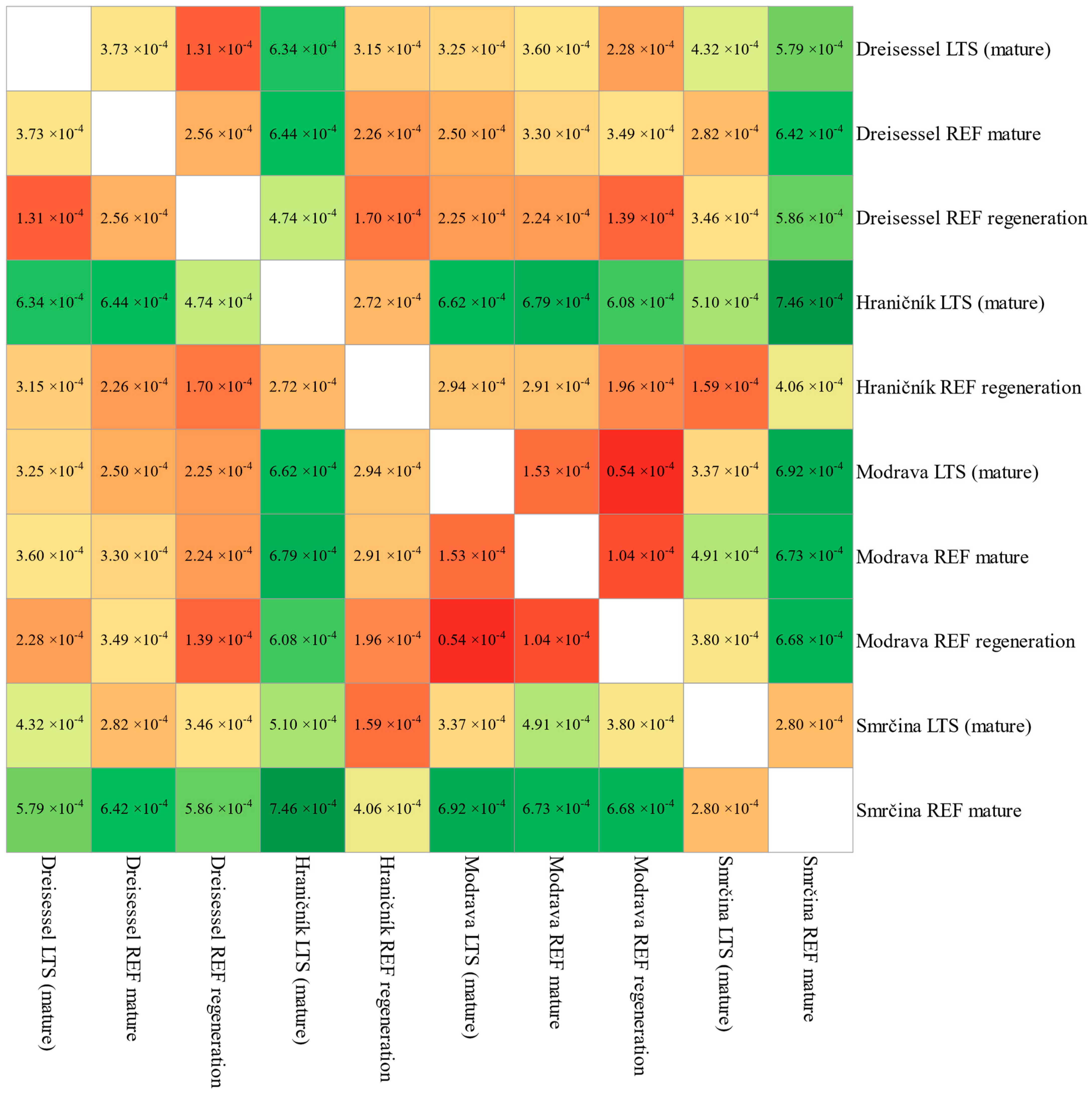
3.1. Population Structure
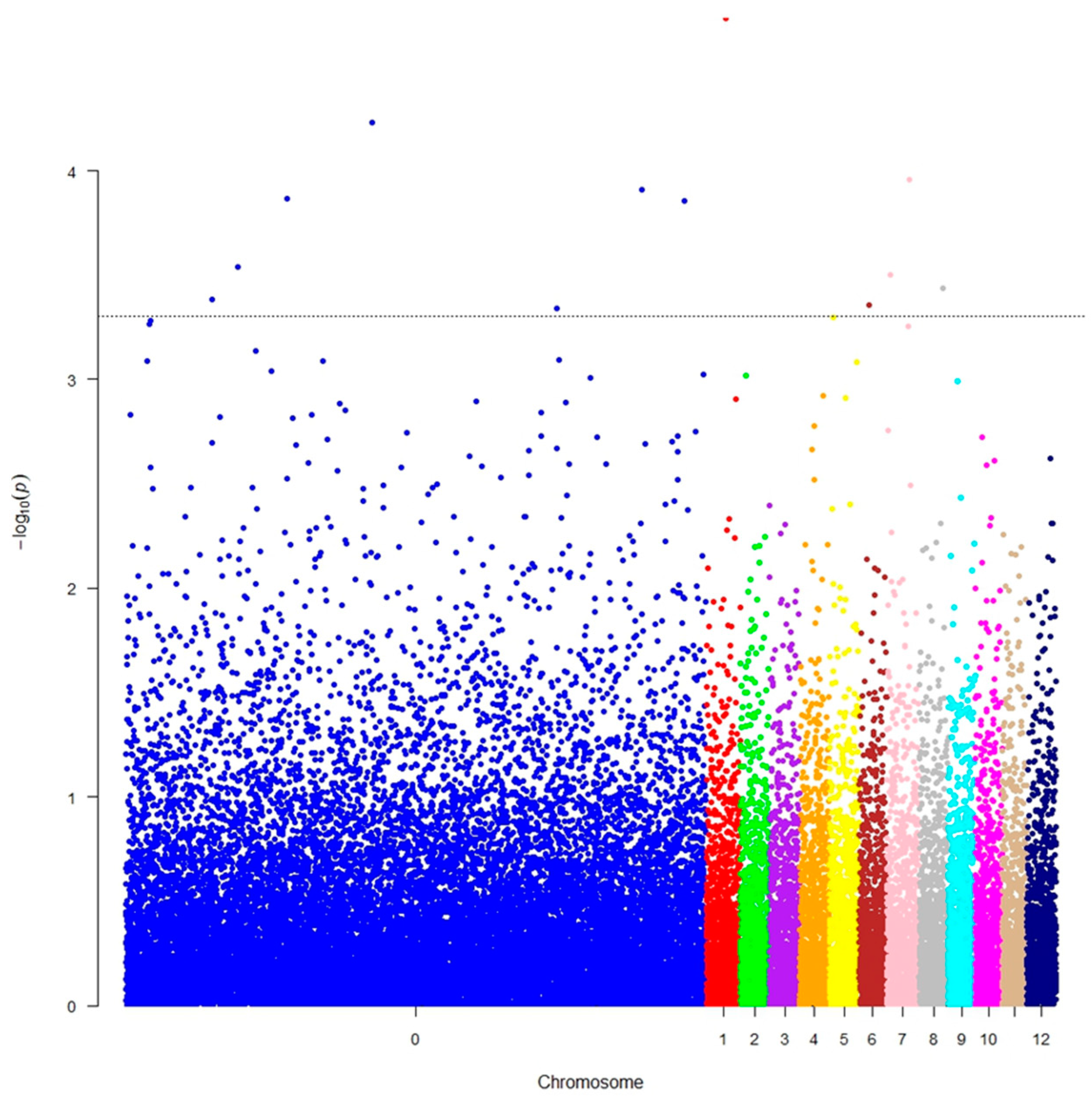
3.2. GWAS Analysis and Gene Identification
4. Discussion
4.1. Population Genetic Structure
4.2. GWAS Analysis and Gene Identification
4.3. Tree Survival—Last Trees Standing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wermelinger, B. Ecology and Management of the Spruce Bark Beetle Ips Typographus—A Review of Recent Research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Vega, F.E.; Hofstetter, R.W. Bark Beetles: Biology and Ecology of Native and Invasive Species; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Ranger, C.M. Bark Beetles: Biology and Ecology of Native and Invasive Species. Am. Entomol. 2016, 62, 196–198. [Google Scholar] [CrossRef]
- Stauffer, C.; Lakatos, F.; Hewitt, G.M. Phylogeography and Postglacial Colonization Routes of Ips Typographus L. (Coleoptera, Scolytidae). Mol. Ecol. 1999, 8, 763–773. [Google Scholar] [CrossRef]
- Nardi, D.; Jactel, H.; Pagot, E.; Samalens, J.; Marini, L. Drought and Stand Susceptibility to Attacks by the European Spruce Bark Beetle: A Remote Sensing Approach. Agric. For. Entomol. 2023, 25, 119–129. [Google Scholar] [CrossRef]
- Cooper, L.A.; Reed, C.C.; Ballantyne, A.P. Mountain Pine Beetle Attack Faster Growing Lodgepole Pine at Low Elevations in Western Montana, USA. For. Ecol. Manag. 2018, 427, 200–207. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing Forest Disturbances in Europe and Their Impact on Carbon Storage. Nat. Clim. Change 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Långström, B.; Lindelöw, Å.; Schroeder, M.; Björklund, N.; Öhrn, P. The Spruce Bark Beetle Outbreak in Sweden Following the January-Storms in 2005 and 2007. 2009. Available online: https://pub.epsilon.slu.se/5076/1/langstrom_b_etal_100823.pdf (accessed on 10 April 2023).
- Nohrstedt, D.; Parker, C. The Public Policy Dimension of Resilience in Natural Disaster Management: Sweden’s Gudrun and Per Storms. In Disaster and Development. Environmental Hazards; Springer: Cham, Switzerland, 2014; pp. 235–253. [Google Scholar]
- Toth, D.; Maitah, M.; Maitah, K.; Jarolínová, V. The Impacts of Calamity Logging on the Development of Spruce Wood Prices in Czech Forestry. Forests 2020, 11, 283. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-Scale Drivers of Natural Disturbances Prone to Anthropogenic Amplification: The Dynamics of Bark Beetle Eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Weed, A.S.; Ayres, M.P.; Bentz, B.J. Chapter 4—Population Dynamics of Bark Beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 157–176. ISBN 978-0-12-417156-5. [Google Scholar]
- Biedermann, P.H.W.; Müller, J.; Grégoire, J.-C.; Gruppe, A.; Hagge, J.; Hammerbacher, A.; Hofstetter, R.W.; Kandasamy, D.; Kolarik, M.; Kostovcik, M.; et al. Bark Beetle Population Dynamics in the Anthropocene: Challenges and Solutions. Trends Ecol. Evol. 2019, 34, 914–924. [Google Scholar] [CrossRef]
- Schiebe, C.; Hammerbacher, A.; Birgersson, G.; Witzell, J.; Brodelius, P.E.; Gershenzon, J.; Hansson, B.S.; Krokene, P.; Schlyter, F. Inducibility of Chemical Defenses in Norway Spruce Bark Is Correlated with Unsuccessful Mass Attacks by the Spruce Bark Beetle. Oecologia 2012, 170, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Brignolas, F.; Lacroix, B.; Lieutier, F.; Sauvard, D.; Drouet, A.; Claudot, A.C.; Yart, A.; Berryman, A.A.; Christiansen, E. Induced Responses in Phenolic Metabolism in Two Norway Spruce Clones after Wounding and Inoculations with Ophiostoma Polonicum, a Bark Beetle-Associated Fungus. Plant Physiol. 1995, 109, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Brignolas, F.; Lieutier, F.; Sauvard, D.; Christiansen, E.; Berryman, A.A. Phenolic Predictors for Norway Spruce Resistance to the Bark Beetle Ips Typographus (Coleoptera: Scolytidae) and an Associated Fungus, Ceratocystis Polonica. Can. J. For. Res. 1998, 28, 720–728. [Google Scholar] [CrossRef]
- Lieutier, F.; Brignolas, F.; Sauvard, D.; Yart, A.; Galet, C.; Brunet, M.; van de Sype, H. Intra- and Inter-Provenance Variability in Phloem Phenols of Picea Abies and Relationship to a Bark Beetle-Associated Fungus. Tree Physiol. 2003, 23, 247–256. [Google Scholar] [CrossRef]
- Korolyova, N.; Buechling, A.; Lieutier, F.; Yart, A.; Cudlín, P.; Turčáni, M.; Jakuš, R. Primary and Secondary Host Selection by Ips Typographus Depends on Norway Spruce Crown Characteristics and Phenolic-Based Defenses. Plant Sci. 2022, 321, 111319. [Google Scholar] [CrossRef]
- Faccoli, M.; Schlyter, F. Conifer Phenolic Resistance Markers Are Bark Beetle Antifeedant Semiochemicals. Agric. For. Entomol. 2007, 9, 237–245. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and Chemical Defenses of Conifer Bark against Bark Beetles and Other Pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef]
- Christiansen, E.; Waring, R.H.; Berryman, A.A. Resistance of Conifers to Bark Beetle Attack: Searching for General Relationships. For. Ecol. Manag. 1987, 22, 89–106. [Google Scholar] [CrossRef]
- Hawkins, C.D.B.; Dhar, A.; Balliet, N.A.; Runzer, K.D. Residual Mature Trees and Secondary Stand Structure after Mountain Pine Beetle Attack in Central British Columbia. For. Ecol. Manag. 2012, 277, 107–115. [Google Scholar] [CrossRef]
- Six, D.L.; Vergobbi, C.; Cutter, M. Are Survivors Different? Genetic-Based Selection of Trees by Mountain Pine Beetle During a Climate Change-Driven Outbreak in a High-Elevation Pine Forest. Front. Plant Sci. 2018, 9, 993. [Google Scholar] [CrossRef]
- Korolyova, N.; Buechling, A.; Ďuračiová, R.; Zabihi, K.; Turčáni, M.; Svoboda, M.; Bláha, J.; Swarts, K.; Poláček, M.; Hradecký, J.; et al. The Last Trees Standing: Climate Modulates Tree Survival Factors during a Prolonged Bark Beetle Outbreak in Europe. Agric. For. Meteorol. 2022, 322, 109025. [Google Scholar] [CrossRef]
- Jakuš, R.; Edwards-Jonášová, M.; Cudlín, P.; Blaženec, M.; Ježík, M.; Havlíček, F.; Moravec, I. Characteristics of Norway Spruce Trees (Picea Abies) Surviving a Spruce Bark Beetle (Ips Typographus L.) Outbreak. Trees 2011, 25, 965–973. [Google Scholar] [CrossRef]
- Erbilgin, N.; Cale, J.A.; Hussain, A.; Ishangulyyeva, G.; Klutsch, J.G.; Najar, A.; Zhao, S. Weathering the Storm: How Lodgepole Pine Trees Survive Mountain Pine Beetle Outbreaks. Oecologia 2017, 184, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed]
- Azaiez, A.; Pavy, N.; Gérardi, S.; Laroche, J.; Boyle, B.; Gagnon, F.; Mottet, M.-J.; Beaulieu, J.; Bousquet, J. A Catalog of Annotated High-Confidence SNPs from Exome Capture and Sequencing Reveals Highly Polymorphic Genes in Norway Spruce (Picea Abies). BMC Genom. 2018, 19, 942. [Google Scholar] [CrossRef]
- Bernhardsson, C.; Zan, Y.; Chen, Z.; Ingvarsson, P.K.; Wu, H.X. Development of a Highly Efficient 50K Single Nucleotide Polymorphism Genotyping Array for the Large and Complex Genome of Norway Spruce (Picea Abies L. Karst) by Whole Genome Resequencing and Its Transferability to Other Spruce Species. Mol. Ecol. Resour. 2021, 21, 880–896. [Google Scholar] [CrossRef]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway Spruce Genome Sequence and Conifer Genome Evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; George, J.-P.; Fluch, S.; Geburek, T.; Grabner, M.; Karanitsch-Ackerl, S.; Konrad, H.; Mayer, K.; Sehr, E.M.; Wischnitzki, E.; et al. Drought Sensitivity of Norway Spruce at the Species’ Warmest Fringe: Quantitative and Molecular Analysis Reveals High Genetic Variation Among and Within Provenances. G3 Genes Genomes Genetics 2018, 8, 1225–1245. [Google Scholar] [CrossRef]
- Čepl, J.; Stejskal, J.; Korecký, J.; Hejtmánek, J.; Faltinová, Z.; Lstibůrek, M.; Gezan, S. The Dehydrins Gene Expression Differs across Ecotypes in Norway Spruce and Relates to Weather Fluctuations. Sci. Rep. 2020, 10, 20789. [Google Scholar] [CrossRef]
- Baison, J.; Vidalis, A.; Zhou, L.; Chen, Z.; Li, Z.; Sillanpää, M.J.; Bernhardsson, C.; Scofield, D.; Forsberg, N.; Grahn, T.; et al. Genome-wide Association Study Identified Novel Candidate Loci Affecting Wood Formation in Norway Spruce. Plant J. 2019, 100, 83–100. [Google Scholar] [CrossRef]
- Hrivnák, M.; Krajmerová, D.; Kurjak, D.; Konôpková, A.; Magni, F.; Scaglione, D.; Ditmarová, L.; Jamnická, G.; Marešová, J.; Gömöry, D. Differential Associations between Nucleotide Polymorphisms and Physiological Traits in Norway Spruce (Picea Abies Karst.) Plants under Contrasting Water Regimes. Forestry 2022, 95, 686–697. [Google Scholar] [CrossRef]
- Korecký, J.; Čepl, J.; Stejskal, J.; Faltinová, Z.; Dvořák, J.; Lstibůrek, M.; El-Kassaby, Y.A. Genetic Diversity of Norway Spruce Ecotypes Assessed by GBS-Derived SNPs. Sci. Rep. 2021, 11, 23119. [Google Scholar] [CrossRef]
- Chen, Z.-Q.; Zan, Y.; Milesi, P.; Zhou, L.; Chen, J.; Li, L.; Cui, B.; Niu, S.; Westin, J.; Karlsson, B.; et al. Leveraging Breeding Programs and Genomic Data in Norway Spruce (Picea abies L. Karst) for GWAS Analysis. Genome Biol. 2021, 22, 179. [Google Scholar] [CrossRef]
- Yanchuk, A.D.; Murphy, J.C.; Wallin, K.F. Evaluation of Genetic Variation of Attack and Resistance in Lodgepole Pine in the Early Stages of a Mountain Pine Beetle Outbreak. Tree Genet. Genomes 2008, 4, 171–180. [Google Scholar] [CrossRef]
- de la Mata, R.; Hood, S.; Sala, A. Insect Outbreak Shifts the Direction of Selection from Fast to Slow Growth Rates in the Long-Lived Conifer Pinus ponderosa. Proc. Natl. Acad. Sci. USA 2017, 114, 7391–7396. [Google Scholar] [CrossRef]
- Svoboda, M.; Janda, P.; Nagel, T.A.; Fraver, S.; Rejzek, J.; Bače, R. Disturbance History of an Old-Growth Sub-Alpine Picea Abies Stand in the Bohemian Forest, Czech Republic. J. Veg. Sci. 2012, 23, 86–97. [Google Scholar] [CrossRef]
- Křenová, Z.; Vrba, J. Just How Many Obstacles Are There to Creating a National Park? A Case Study from the Šumava National Park. Eur. J. Environ. Sci. 2014, 4. [Google Scholar] [CrossRef]
- Lausch, A.; Heurich, M.; Fahse, L. Spatio-Temporal Infestation Patterns of Ips typographus (L.) in the Bavarian Forest National Park, Germany. Ecol. Indic. 2013, 31, 73–81. [Google Scholar] [CrossRef]
- Massicotte, P.; South, A.; Hufkens, K. Rnaturalearth: World Map Data from Natural Earth 2023. Available online: https://github.com/ropensci/rnaturalearth (accessed on 30 April 2023).
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- VanRaden, P.M. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New Tools for the Analysis of Genome-Wide SNP Data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.J. Mmod: An R Library for the Calculation of Population Differentiation Statistics. Mol. Ecol. Resour. 2012, 12, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Sundell, D.; Mannapperuma, C.; Netotea, S.; Delhomme, N.; Lin, Y.-C.; Sjödin, A.; Van de Peer, Y.; Jansson, S.; Hvidsten, T.R.; Street, N.R. The Plant Genome Integrative Explorer Resource: PlantGenIE. Org. New Phytol. 2015, 208, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Van Bel, M.; Silvestri, F.; Weitz, E.M.; Kreft, L.; Botzki, A.; Coppens, F.; Vandepoele, K. PLAZA 5.0: Extending the Scope and Power of Comparative and Functional Genomics in Plants. Nucleic Acids Res. 2022, 50, D1468–D1474. [Google Scholar] [CrossRef] [PubMed]
- Meinke, D.W.; Cherry, J.M.; Dean, C.; Rounsley, S.D.; Koornneef, M. Arabidopsis Thaliana: A Model Plant for Genome Analysis. Science 1998, 282, 662–682. [Google Scholar] [CrossRef]
- Cieśla, M.; Boguta, M. Regulation of RNA Polymerase III Transcription by Maf1 Protein. Acta Biochim. Pol. 2008, 55, 215–225. [Google Scholar] [CrossRef]
- Heard, W.; Sklenář, J.; Tomé, D.F.A.; Robatzek, S.; Jones, A.M.E. Identification of Regulatory and Cargo Proteins of Endosomal and Secretory Pathways in Arabidopsis Thaliana by Proteomic Dissection *[S]. Mol. Cell. Proteomics 2015, 14, 1796–1813. [Google Scholar] [CrossRef]
- Mlynárová, L.; Nap, J.-P.; Bisseling, T. The SWI/SNF Chromatin-Remodeling Gene AtCHR12 Mediates Temporary Growth Arrest in Arabidopsis Thaliana upon Perceiving Environmental Stress: Chromatin Remodeling in Growth Response to Stress. Plant J. 2007, 51, 874–885. [Google Scholar] [CrossRef]
- Maghuly, F.; Pinsker, W.; Praznik, W.; Fluch, S. Genetic Diversity in Managed Subpopulations of Norway Spruce [Picea abies (L.) Karst.]. For. Ecol. Manag. 2006, 222, 266–271. [Google Scholar] [CrossRef]
- Meloni, M.; Perini, D.; Binelli, G. The Distribution of Genetic Variation in Norway Spruce (Picea Abies Karst.) Populations in the Western Alps. J. Biogeogr. 2007, 34, 929–938. [Google Scholar] [CrossRef]
- Tollefsrud, M.M.; Sønstebø, J.H.; Brochmann, C.; Johnsen, Ø.; Skrøppa, T.; Vendramin, G.G. Combined Analysis of Nuclear and Mitochondrial Markers Provide New Insight into the Genetic Structure of North European Picea Abies. Heredity 2009, 102, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Stojnić, S.; Avramidou, E.V.; Fussi, B.; Westergren, M.; Orlović, S.; Matović, B.; Trudić, B.; Kraigher, H.; Aravanopoulos, F.A.; Konnert, M. Assessment of Genetic Diversity and Population Genetic Structure of Norway Spruce (Picea abies (L.) Karsten) at Its Southern Lineage in Europe. Implications for Conservation of Forest Genetic Resources. Forests 2019, 10, 258. [Google Scholar] [CrossRef]
- Binova, Z.; Korecky, J.; Dvorak, J.; Bily, J.; Zadrapova, D.; Jansa, V.; Lstiburek, M. Genetic Structure of Norway Spruce Ecotypes Studied by SSR Markers. Forests 2020, 11, 110. [Google Scholar] [CrossRef]
- Chen, J.; Källman, T.; Ma, X.; Gyllenstrand, N.; Zaina, G.; Morgante, M.; Bousquet, J.; Eckert, A.; Wegrzyn, J.; Neale, D. Disentangling the Roles of History and Local Selection in Shaping Clinal Variation of Allele Frequencies and Gene Expression in Norway Spruce (Picea Abies). Genetics 2012, 191, 865–881. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bernhardsson, C.; Ingvarsson, P.K. Demography and Natural Selection Have Shaped Genetic Variation in the Widely Distributed Conifer Norway Spruce (Picea abies). Genome Biol. Evol. 2020, 12, 3803–3817. [Google Scholar] [CrossRef] [PubMed]
- Di-Giovanni, F.; Kevan, P.G.; Arnold, J. Lower Planetary Boundary Layer Profiles of Atmospheric Conifer Pollen above a Seed Orchard in Northern Ontario, Canada. For. Ecol. Manag. 1996, 83, 87–97. [Google Scholar] [CrossRef]
- Burczyk, J.; Lewandowski, A.; Chalupka, W. Local Pollen Dispersal and Distant Gene Flow in Norway Spruce (Picea abies [L.] Karst.). For. Ecol. Manag. 2004, 197, 39–48. [Google Scholar] [CrossRef]
- Jansen, S.; Konrad, H.; Geburek, T. The Extent of Historic Translocation of Norway Spruce Forest Reproductive Material in Europe. Ann. For. Sci. 2017, 74, 56. [Google Scholar] [CrossRef]
- Spiecker, H. Silvicultural Management in Maintaining Biodiversity and Resistance of Forests in Europe—Temperate Zone. J. Environ. Manag. 2003, 67, 55–65. [Google Scholar] [CrossRef]
- Brůna, J.; Wild, J.; Svoboda, M.; Heurich, M.; Müllerová, J. Impacts and Underlying Factors of Landscape-Scale, Historical Disturbance of Mountain Forest Identified Using Archival Documents. For. Ecol. Manag. 2013, 305, 294–306. [Google Scholar] [CrossRef]
- Čada, V.; Morrissey, R.C.; Michalová, Z.; Bače, R.; Janda, P.; Svoboda, M. Frequent Severe Natural Disturbances and Non-Equilibrium Landscape Dynamics Shaped the Mountain Spruce Forest in Central Europe. For. Ecol. Manag. 2016, 363, 169–178. [Google Scholar] [CrossRef]
- Průša, E. Die Bohmischen Und Mahrischen Urwalder-Ihre Struktur Und Okologie/The Bohmian and Mahrian Primeval Forests-Their Structure and Ecology; Tschechoslowakischen Akademie der Wissenschaften: Bratislava, Slovakia, 1985. [Google Scholar]
- Armstrong, R.A. When to Use the Bonferroni Correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Rothman, K.J. No Adjustments Are Needed for Multiple Comparisons. Epidemiology 1990, 1, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Perneger, T.V. What’s Wrong with Bonferroni Adjustments. BMJ 1998, 316, 1236–1238. [Google Scholar] [CrossRef]
- Graczyk, D.; Cieśla, M.; Boguta, M. Regulation of tRNA Synthesis by the General Transcription Factors of RNA Polymerase III—TFIIIB and TFIIIC, and by the MAF1 Protein. Biochim. Biophys. Acta BBA—Gene Regul. Mech. 2018, 1861, 320–329. [Google Scholar] [CrossRef]
- Blayney, J.; Geary, J.; Chrisp, R.; Violet, J.; Barratt, L.; Tavukçu, L.; Paine, K.; Vaistij, F.E.; Graham, I.A.; Denby, K.J.; et al. Impact on Arabidopsis Growth and Stress Resistance of Depleting the Maf1 Repressor of RNA Polymerase III. Gene 2022, 815, 146130. [Google Scholar] [CrossRef]
- Soprano, A.S.; Abe, V.Y.; Smetana, J.H.C.; Benedetti, C.E. Citrus MAF1, a Repressor of RNA Polymerase III, Binds the Xanthomonas Citri Canker Elicitor PthA4 and Suppresses Citrus Canker Development. Plant Physiol. 2013, 163, 232–242. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Kovac, K.A. Plant DNA Methyltransferases. In Plant Gene Silencing; Matzke, M.A., Matzke, A.J.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 69–81. ISBN 978-94-011-4183-3. [Google Scholar]
- Liu, J.; He, Z. Small DNA Methylation, Big Player in Plant Abiotic Stress Responses and Memory. Front. Plant Sci. 2020, 11, 595603. [Google Scholar] [CrossRef]
- Jiang, C.; Mithani, A.; Belfield, E.J.; Mott, R.; Hurst, L.D.; Harberd, N.P. Environmentally Responsive Genome-Wide Accumulation of de Novo Arabidopsis Thaliana Mutations and Epimutations. Genome Res. 2014, 24, 1821–1829. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Paszkowski, J. Heat-Induced Release of Epigenetic Silencing Reveals the Concealed Role of an Imprinted Plant Gene. PLoS Genet. 2014, 10, e1004806. [Google Scholar] [CrossRef]
- Wibowo, A.; Becker, C.; Marconi, G.; Durr, J.; Price, J.; Hagmann, J.; Papareddy, R.; Putra, H.; Kageyama, J.; Becker, J. Hyperosmotic Stress Memory in Arabidopsis Is Mediated by Distinct Epigenetically Labile Sites in the Genome and Is Restricted in the Male Germline by DNA Glycosylase Activity. Elife 2016, 5, e13546. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.-K. Molecular and Genetic Aspects of Plant Responses to Osmotic Stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Millar, C.I.; Westfall, R.D.; Delany, D.L.; Bokach, M.J.; Flint, A.L.; Flint, L.E. Forest Mortality in High-Elevation Whitebark Pine (Pinus Albicaulis) Forests of Eastern California, USA; Influence of Environmental Context, Bark Beetles, Climatic Water Deficit, and Warming. Can. J. For. Res. 2012, 42, 749–765. [Google Scholar] [CrossRef]
- Knapp, E.E.; Bernal, A.A.; Kane, J.M.; Fettig, C.J.; North, M.P. Variable Thinning and Prescribed Fire Influence Tree Mortality and Growth during and after a Severe Drought. For. Ecol. Manag. 2021, 479, 118595. [Google Scholar] [CrossRef]
- Ferrenberg, S.; Kane, J.M.; Mitton, J.B. Resin Duct Characteristics Associated with Tree Resistance to Bark Beetles across Lodgepole and Limber Pines. Oecologia 2014, 174, 1283–1292. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Linares, J.C.; Camarero, J.J. Reduced Growth Sensitivity to Climate in Bark-Beetle Infested Aleppo Pines: Connecting Climatic and Biotic Drivers of Forest Dieback. For. Ecol. Manag. 2015, 357, 126–137. [Google Scholar] [CrossRef]
- Reed, C.C.; Hood, S.M. Few Generalizable Patterns of Tree-Level Mortality during Extreme Drought and Concurrent Bark Beetle Outbreaks. Sci. Total Environ. 2021, 750, 141306. [Google Scholar] [CrossRef]
- Rhoades, D.F. Herbivore Population Dynamics and Plant Chemistry. In Variable Plants and Herbivores in Natural and Managed Systems; Academic Press: New York, NY, USA, 1983; Volume 6, pp. 155–220. [Google Scholar]
- Waring, R.H.; Pitman, G.B. Modifying Lodgepole Pine Stands to Change Susceptibility to Mountain Pine Beetle Attack. Ecology 1985, 66, 889–897. [Google Scholar] [CrossRef]
- Mattson, W.J.; Haack, R.A. The Role of Drought in Outbreaks of Plant-Eating Insects. Bioscience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The Dilemma of Plants: To Grow or Defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Landis, R.M.; Peart, D.R. Early Performance Predicts Canopy Attainment across Life Histories in Subalpine Forest Trees. Ecology 2005, 86, 63–72. [Google Scholar] [CrossRef]
- Petit, R.J.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef]
- Reed, D.E.; Ewers, B.E.; Pendall, E.; Frank, J.; Kelly, R. Bark Beetle-Induced Tree Mortality Alters Stand Energy Budgets Due to Water Budget Changes. Theor. Appl. Climatol. 2018, 131, 153–165. [Google Scholar] [CrossRef]
- Koontz, M.J.; Latimer, A.M.; Mortenson, L.A.; Fettig, C.J.; North, M.P. Cross-Scale Interaction of Host Tree Size and Climatic Water Deficit Governs Bark Beetle-Induced Tree Mortality. Nat. Commun. 2021, 12, 129. [Google Scholar] [CrossRef]
- Huang, J.; Kautz, M.; Trowbridge, A.M.; Hammerbacher, A.; Raffa, K.F.; Adams, H.D.; Goodsman, D.W.; Xu, C.; Meddens, A.J.H.; Kandasamy, D.; et al. Tree Defence and Bark Beetles in a Drying World: Carbon Partitioning, Functioning and Modelling. New Phytol. 2020, 225, 26–36. [Google Scholar] [CrossRef]
- Mageroy, M.H.; Christiansen, E.; Långström, B.; Borg-Karlson, A.-K.; Solheim, H.; Björklund, N.; Zhao, T.; Schmidt, A.; Fossdal, C.G.; Krokene, P. Priming of Inducible Defenses Protects Norway Spruce against Tree-Killing Bark Beetles. Plant Cell Environ. 2020, 43, 420–430. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | GPS Coordinates | Total Number of Trees | |||
|---|---|---|---|---|---|
| LTS Trees | Reference Trees | ||||
| Mature | Juvenile | ||||
| Dreisessel | 48°47′ N 13°48′ E | 92 | 20 | 18 | 54 |
| Hraničník | 48°45′ N 13°55′ E | 56 | 15 | 0 | 41 |
| Modrava | 48°59′ N 13°26′ E | 168 | 48 | 33 | 87 |
| Smrčina | 48°44′ N 13°56′ E | 67 | 19 | 48 | 0 |
| Comparison | Site | Jost’s D | Confidence Interval | |
|---|---|---|---|---|
| Lower | Upper | |||
| (a) LTS and reference trees | Dreisessel | 1.42 × 10−4 | 0.25 × 10−4 | 2.58 × 10−4 |
| Hraničník | 2.72 × 10−4 | 1.09 × 10−4 | 4.35 × 10−4 | |
| Modrava | 0.64 × 10−4 | 0.03 × 10−4 | 1.24 × 10−4 | |
| Smrčina | 2.80 × 10−4 | 1.36 × 10−4 | 4.24 × 10−4 | |
| (b) LTS between sites | Smrčina × Modrava | 3.37 × 10−4 | 1.80 × 10−4 | 4.94 × 10−4 |
| Smrčina × Hraničník | 5.10 × 10−4 | 2.43 × 10−4 | 7.77 × 10−4 | |
| Smrčina × Dreisessel | 4.32 × 10−4 | 2.41 × 10−4 | 6.24 × 10−4 | |
| Modrava × Hraničník | 6.62 × 10−4 | 4.62 × 10−4 | 8.63 × 10−4 | |
| Modrava × Dreisessel | 3.25 × 10−4 | 1.63 × 10−4 | 4.87 × 10−4 | |
| Hraničník × Dreisessel | 6.34 × 10−4 | 3.70 × 10−4 | 8.99 × 10−4 | |
| (c) Reference trees between sites | Smrčina × Modrava | 6.47 × 10−4 | 5.80 × 10−4 | 7.14 × 10−4 |
| Smrčina × Hraničník | 4.06 × 10−4 | 3.01 × 10−4 | 5.11 × 10−4 | |
| Smrčina × Dreisessel | 5.48 × 10−4 | 4.72 × 10−4 | 6.25 × 10−4 | |
| Modrava × Hraničník | 2.01 × 10−4 | 1.35 × 10−4 | 2.67 × 10−4 | |
| Modrava × Dreisessel | 1.38 × 10−4 | 0.87 × 10−4 | 1.89 × 10−4 | |
| Hraničník × Dreisessel | 1.35 × 10−4 | 0.42 × 10−4 | 2.28 × 10−4 | |
| Contig ID | Marker ID | Gene (Plantgenie.org) | Gene (PLAZA) | LG | p-Value |
|---|---|---|---|---|---|
| MA_139355 | AX-302167819 | --- | --- | NA | 4.15 × 10−4 |
| MA_17088 | AX-305072589 | MA_17088g0010 | PAB00027290 | 7 | 2.85 × 10−4 |
| MA_35335 | AX-305188807 | MA_35335g0010 | PAB00038352 | 5 | 4.90 × 10−4 |
| MA_466244 | AX-306784220 | MA_466244g0010 | PAB00043107 | 1 | 1.72 × 10−5 |
| MA_496531 | AX-305623507 | MA_496531g0010 | PAB00044508 | 6 | 4.34 × 10−4 |
| MA_51088 | AX-308536830 | MA_51088g0010 | PAB00045124 | 7 | 1.13 × 10−4 |
| MA_538811 | AX-306985564 | --- | --- | NA | 4.62 × 10−4 |
| MA_539 | AX-309072172 | --- | --- | NA | 1.29 × 10−4 |
| MA_77097 | AX-308569646 | MA_77097g0010 | PAB00054584 | NA | 1.16 × 10−4 |
| MA_818649 | AX-308742628 | MA_818649g0010 | PAB00056594 | 8 | 2.93 × 10−4 |
| MA_8764366 | AX-303041489 | --- | --- | NA | 1.38 × 10−4 |
| MA_914090 | AX-304622666 | --- | --- | NA | 5.76 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korecký, J.; Čepl, J.; Korolyova, N.; Stejskal, J.; Turčáni, M.; Jakuš, R. Resistance to Bark Beetle Outbreak in Norway Spruce: Population Structure Analysis and Comparative Genomic Assessment of Surviving (LTS) and Randomly Selected Reference Trees. Forests 2023, 14, 2074. https://doi.org/10.3390/f14102074
Korecký J, Čepl J, Korolyova N, Stejskal J, Turčáni M, Jakuš R. Resistance to Bark Beetle Outbreak in Norway Spruce: Population Structure Analysis and Comparative Genomic Assessment of Surviving (LTS) and Randomly Selected Reference Trees. Forests. 2023; 14(10):2074. https://doi.org/10.3390/f14102074
Chicago/Turabian StyleKorecký, Jiří, Jaroslav Čepl, Nataliya Korolyova, Jan Stejskal, Marek Turčáni, and Rastislav Jakuš. 2023. "Resistance to Bark Beetle Outbreak in Norway Spruce: Population Structure Analysis and Comparative Genomic Assessment of Surviving (LTS) and Randomly Selected Reference Trees" Forests 14, no. 10: 2074. https://doi.org/10.3390/f14102074