
Mycobiome of Post-Agricultural Soils 20 Years after Application of Organic Substrates and Planting of Pine Seedlings
 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mulching of the Soil
2.2. DNA Extraction
2.3. Libraries Preparation
2.4. Processing and Analysis of Sequencing Data
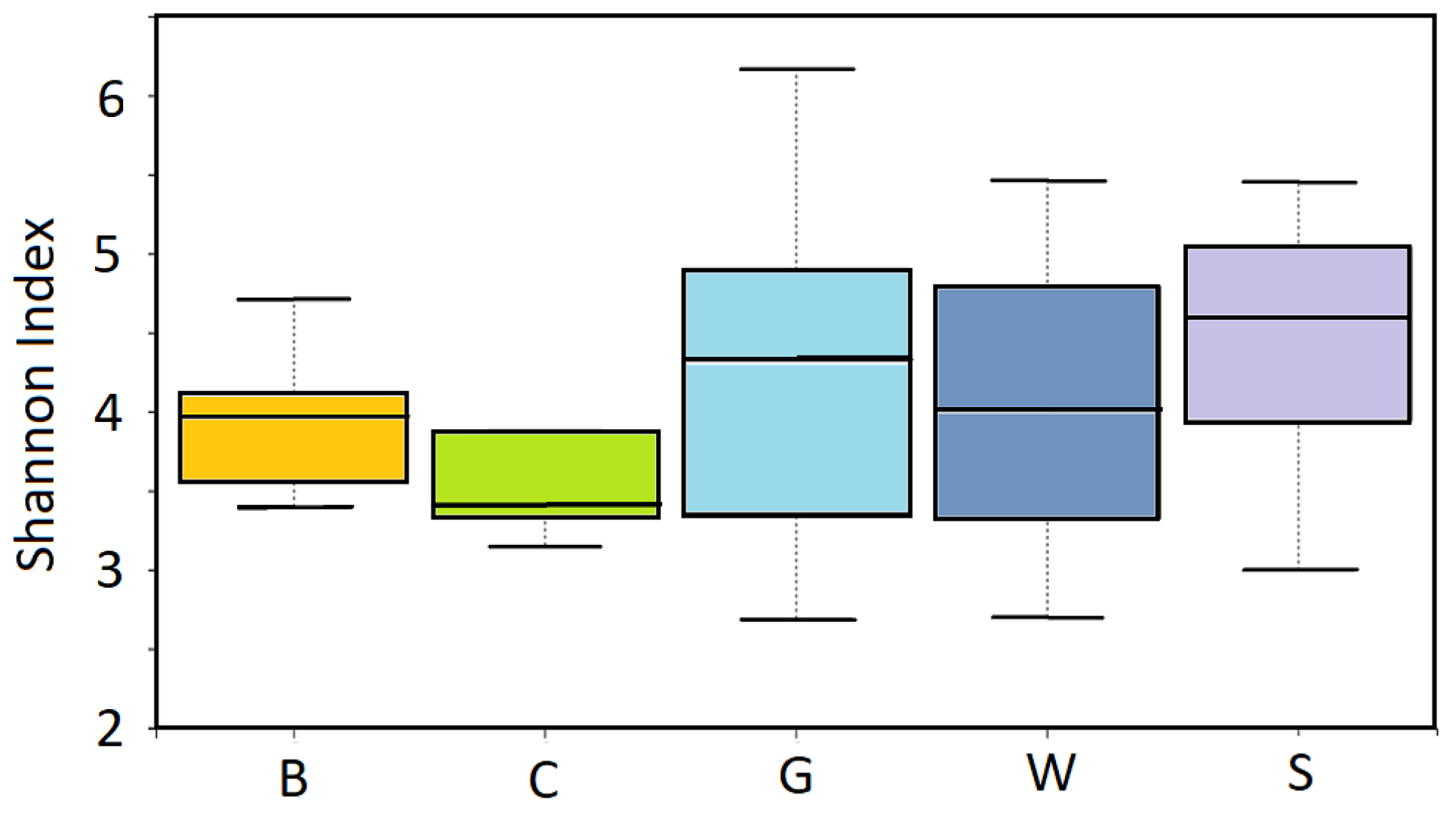
2.5. Analysis of Fungal Biodiversity
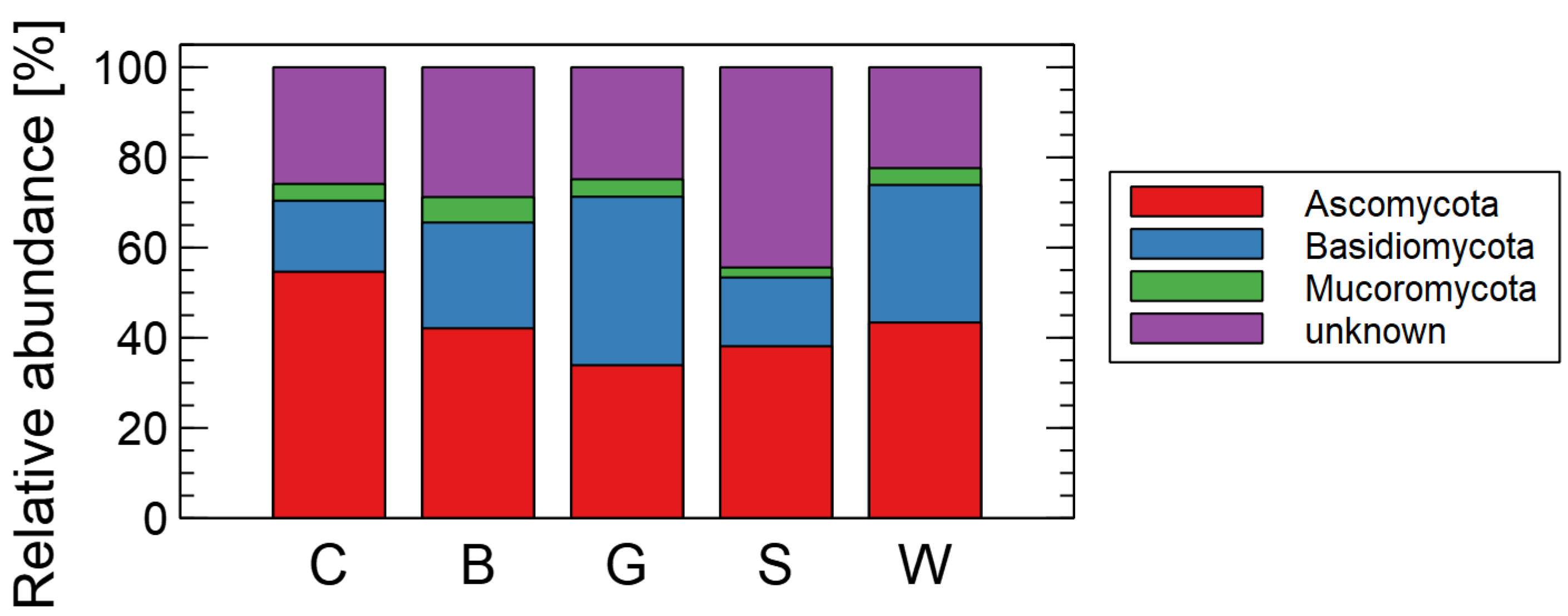
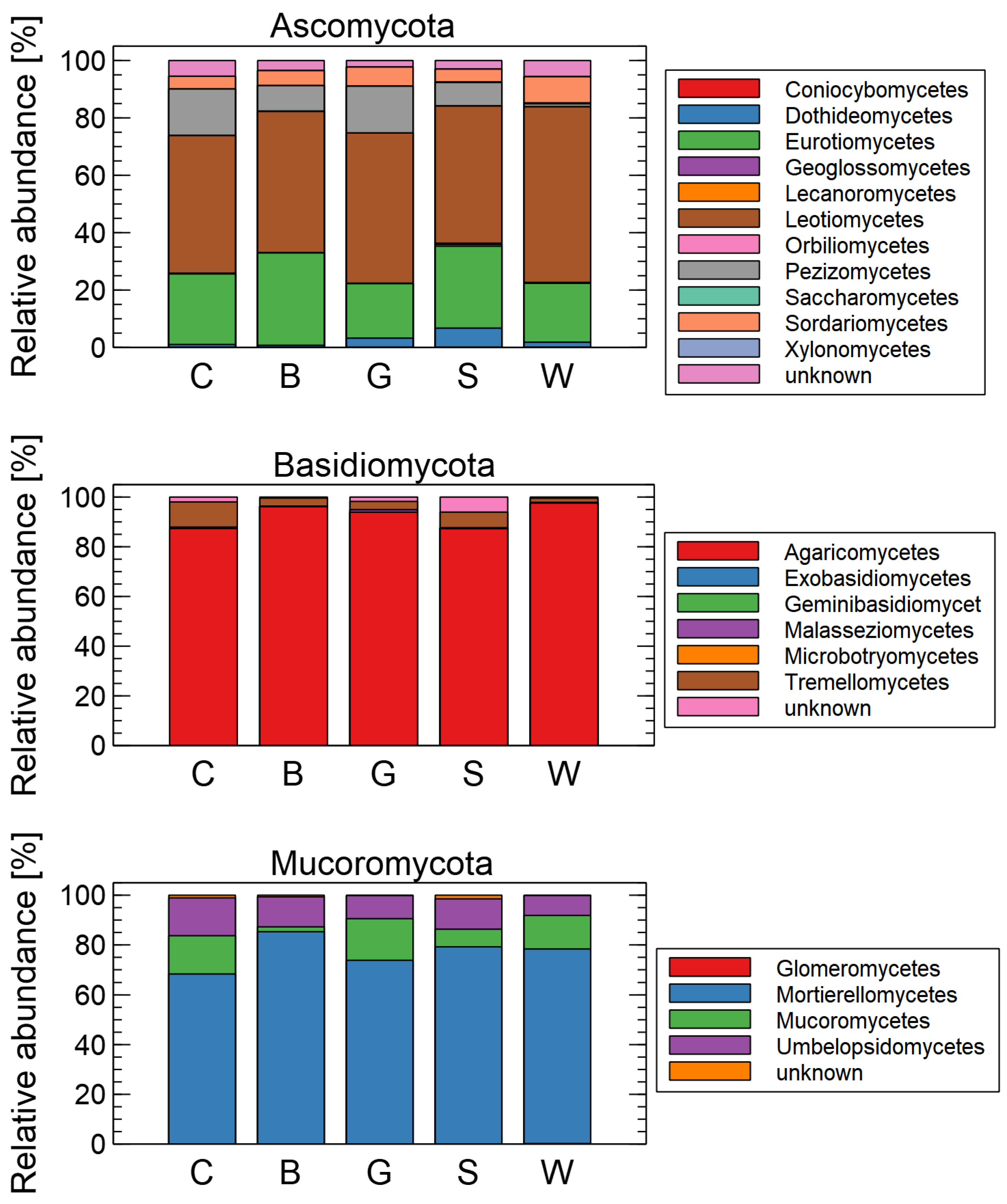
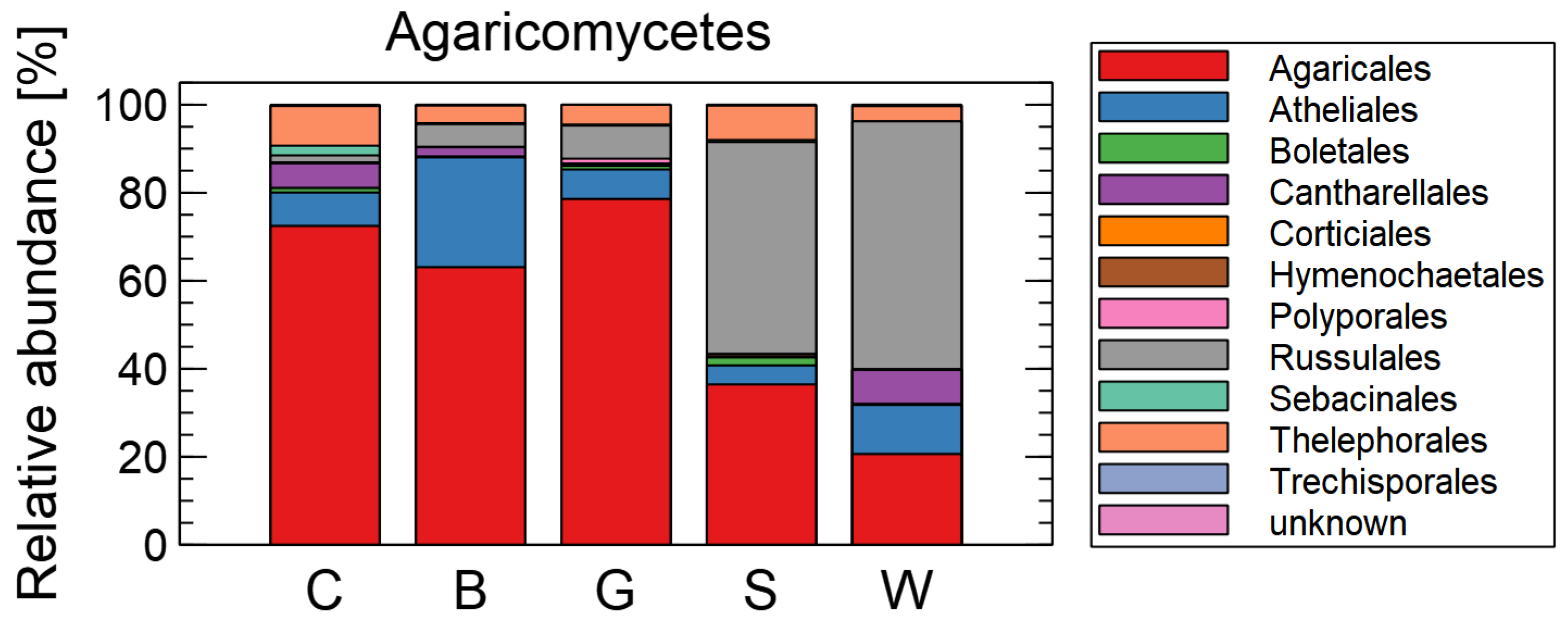
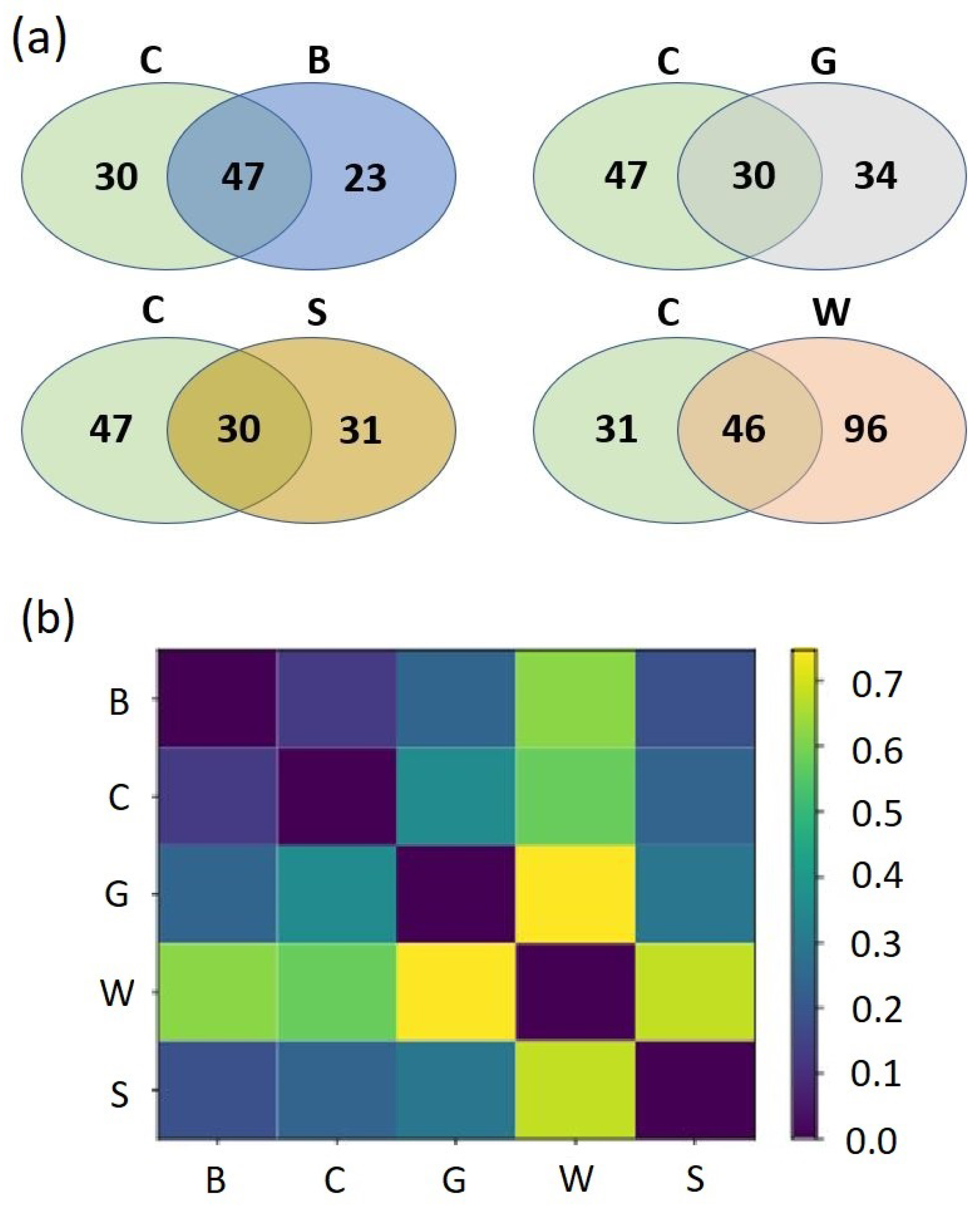
3. Results
Effects of Mulching on the Soil Mycobiome
4. Discussion
4.1. Advantages of Adding Organic Substances
4.2. Protective Effects of Identified Mycorrhizae against Heterobasidion spp. and Other Pathogens
4.3. Protective Effect of Identified Mycorrhizal Fungi on Plants against Drought
4.4. Identified Fungi as a Potential Source of Bioactive Molecules
4.5. Antioxidant Capacity and Antimicrobial Activities of Some Identified Fungi
4.6. The Role of Mycorrhizal Fungi in the Nutrient Cycle of Pine Trees
4.7. The Role of Micorrhizal Fungi in the Nutrient Cycle of Pine Trees
4.8. The Role of Antagonistic Fungi in Biological Protection of Pines from Pathogens
4.9. Fungi Identified as Pathogens for Animals and Humans
4.10. Seedborne Fungi
4.11. Edible Ectomycorrhizal Fungi
4.12. Identified Fungi Occurred as Pathogenic to Various Plant Species
4.13. Other Identified Fungi Specific for Different Environments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sierota, Z.; Malecka, M. Ocena zmian w drzewostanie sosnowym na gruncie porolnym po 30 latach od wykonania pierwszych ciec pielegnacyjnych bez zabiegu ochronnego przeciw hubie korzeni. Sylwan 2003, 147, 19–26. [Google Scholar]
- Małecka, M.; Sierota, Z. Ocena zagrożenia i ryzyka rozwoju huby korzeni w drzewostanie na gruncie porolnym. Sylwan 2003, 147, 12–25. [Google Scholar]
- Sierota, Z.; Kwaśna, H. Changes in fungal communities in abandoned farmland soil enriched with pine sawdust. Folia For. Pol. Ser. A For. 1998, 40, 85–94. [Google Scholar]
- Sierota, Z.; Kwaśna, H. Ocena mikologiczna zmian zachodzących w glebie gruntu porolnego po dodaniu trocin iglastych. Sylwan 1999, 143, 57–66. [Google Scholar]
- Kwaśna, H.; Brzeski, M.; Sierota, Z. Mikroorganizmy środowiska glebowego odłogujących gruntów porolnych–zmiany w zbiorowiskach grzybów i nicieni po dodaniu trocin iglastych. In Drobnoustroje Środowiska Glebowego–Aspekty Fizjologiczne, Biochemiczne, Genetyczne; Dahm, W.H., Pokojska, A., Eds.; Wyd. A. Marszałek: Toruń, Poland, 2001; pp. 57–66. [Google Scholar]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.; Khanna, P. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Wall, A.; Hytönen, J. Soil fertility of afforested arable land compared to continuously. Plant Soil 2005, 275, 247–260. [Google Scholar] [CrossRef]
- Olejarski, I. Wplyw zabiegow agrotechnicznych na niektore wlasciwosci gleb oraz stan upraw sosnowych na pozarzyskach wielkoobszarowych. Pr. Inst. Badaw. LeŚNictwa Ser. A 2003, 2, 47–77. [Google Scholar]
- Kwaśna, H.; Sierota, Z.; Bateman, G.L. Fungal communities in fallow soil before and after amending with pine sawdust. Appl. Soil Ecol. 2000, 14, 177–182. [Google Scholar] [CrossRef]
- Duda, B.; Sierota, Z. Survival of Scots pine seedlings after biological and chemical control of damping-off in plastic greenhouses. Eur. J. For. Pathol. 1987, 17, 110–117. [Google Scholar] [CrossRef]
- Retter, A.; Nilsson, R.H.; Bourlat, S.J. Exploring the taxonomic composition of two fungal communities on the Swedish west coast through metabarcoding. Biodivers. Data J. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Põlme, S.; Riit, T.; Liiv, I.; Kõljalg, U.; Kisand, V.; Nilsson, H.; Hildebrand, F.; et al. Shotgun metagenomes and multiple primer pair-barcode combinations of amplicons reveal biases in metabarcoding analyses of fungi. MycoKeys 2015, 10, 1–43. [Google Scholar] [CrossRef]
- Wurzbacher, C.; Nilsson, R.H.; Rautio, M.; Peura, S. Poorly known microbial taxa dominate the microbiome of permafrost thaw ponds. ISME J. 2017, 11, 1938–1941. [Google Scholar] [CrossRef] [Green Version]
- FastQC Project. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 November 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Marcelino, V.R.; Clausen, P.T.L.C.; Buchmann, J.P.; Wille, M.; Iredell, J.R.; Meyer, W.; Lund, O.; Sorrell, T.C.; Holmes, E.C. CCMetagen: Comprehensive and accurate identification of eukaryotes and prokaryotes in metagenomic data. Genome Biol. 2020, 21. [Google Scholar] [CrossRef]
- Clausen, P.T.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 1–8. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.; Crous, P.; Boekhout, T.; et al. DNA barcoding analysis of more than 9 000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Calculate and Draw Custom Venn Diagrams. Available online: http://bioinformatics.psb.ugent.be/webtools/Venn (accessed on 1 November 2022).
- Scikit-Bio Software Package Project. Available online: http://scikit-bio.org/docs/0.5.8/ (accessed on 28 November 2022).
- Nguyen, D.; Boberg, J.; Ihrmark, K.; Stenström, E.; Stenlid, J. Do foliar fungal communities of Norway spruce shift along a tree species diversity gradient in mature European forests? Fungal Ecol. 2016, 23, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Małecka, M.; Hilszczańska, D. Changes in ectomycorrhizal structure of Scots pine growing on abandoned farmland soil enriched with organic substrates. Sylwan 2014, 158, 243–250. [Google Scholar]
- Olejarski, I.; Walendzik, J. Warunki wzrostu upraw lesnych w Bornem Sulinowie. Głos Lasu 2002, 08, 20–22. (In Polish) [Google Scholar]
- Olejarski, I. Wykorzystanie pozostalosci zrebowych do nawozenia organicznego gruntow (in Polish). Postępy Techniki w Leśnictwie 2005, 92, 20–24. [Google Scholar]
- Oszako, T.; Olejarski, I. Inicjowanie procesow przeksztalcenia gleb porolnych w gleby lesne poprzez wykorzystanie pozostalosci zrebowych, kompostow i trocin. Pr. Inst. Badaw. LeŚNictwa Ser. A 2003, 1, 76–79. (In Polish) [Google Scholar]
- Malecka, M.; Hilszczanska, D. Wpływ odłogowania i dodatku trocin iglastych do gleby porolnej na jej właściwości chemiczne i zbiorowisko grzybów ektomykoryzowych 15-letniej sosny zwyczajnej. Leśne Pr. Badaw. 2015, 76, 265–272. [Google Scholar]
- Kuhad, R.C.; Gupta, R.; Singh, A. Microbial cellulases and their industrial applications. Enzym. Res. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- de França Passos, D.; Pereira, N., Jr.; de Castro, A.M. A comparative review of recent advances in cellulases production by Aspergillus, Penicillium and Trichoderma strains and their use for lignocellulose deconstruction. Curr. Opin. Green Sustain. Chem. 2018, 14, 60–66. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Sierota, Z. Wpływ inokulum mikoryzowego grzyba Thelephora terrestri na wzrost sadzonek sosny zwyczajnej Pinus sylvestris L. II. Badania polowe. Sylwan 2006, 150, 20–28. [Google Scholar]
- Kwaśna, H.; Lakomy, P.; Gornowicz, R.; Mikicinski, A.; Behnke-Borowczyk, J.; Galazka, S. Struktura zbiorowisk grzybow i bakterii w glebie 1-rocznej uprawy i 10-letniego mlodnika w zaleznosci od sposobu przygotowania gleby. Sylwan 2015, 159, 71–81. [Google Scholar]
- Kwaśna, H.; Behnke-Borowczyk, J.; Gornowicz, R.; Łakomy, P. Effects of preparation of clearcut forest sites on the soil mycobiota with consequences for Scots pine growth and health. For. Pathol. 2019, 49, e12494. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Hilszczanska, D. Wplyw podlozy szkolkarskich na rozwoj mikoryz sosny Pinus sylvestris L. Sylwan 2000, 144, 93–97. [Google Scholar]
- Klimek, V.M.; Fircanis, S.; Maslak, P.; Guernah, I.; Baum, M.; Wu, N.; Panageas, K.; Wright, J.J.; Pandolfi, P.P.; Nimer, S.D. Tolerability, pharmacodynamics, and pharmacokinetics studies of depsipeptide (romidepsin) in patients with acute myelogenous leukemia or advanced myelodysplastic syndromes. Clin. Cancer Res. 2008, 14, 826–832. [Google Scholar] [CrossRef] [Green Version]
- Buscot, F.; Weber, G.; Oberwinkler, F. Interactions between Cylindrocarpon destructans and ectomycorrhizas of Picea abies with Laccaria laccata and Paxillus involutes. Trees 1992, 6, 83–90. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Pajares, J.; Díez, J. In vitro effects of four ectomycorrhizal fungi, Boletus edulis, Rhizopogon roseolus, Laccaria laccata and Lactarius deliciosus on Fusarium damping off in Pinus nigra seedlings. New For. 2006, 32, 323–334. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Tang, M.; Chen, H.; Tian, Z.Q. Effects of ectomycorrhizal fungi on damping-off and induction of pathogenesis-related proteins in Pinus tabulaeformis seedlings inoculated with Amanita vaginata. For. Pathol. 2011, 41, 262–269. [Google Scholar] [CrossRef]
- Velmala, S.; Vuorinen, I.; Uimari, A.; Piri, T.; Pennanen, T. Ectomycorrhizal fungi increase the vitality of Norway spruce seedlings under the pressure of Heterobasidion root rot in vitro but may increase susceptibility to foliar necrotrophs. Fungal Biol. 2018, 122, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.L.; Massicotte, H.B.; Melville, L.H. Mycorrhizas: Anatomy and Cell Biology; NRC Research Press: Ottawa, ON, Canada, 2004. [Google Scholar]
- Wilcox, H.; Wang, C. Ectomycorrhizal and ectendomycorrhizal associations of Phialophora finlandia with Pinus resinosa, Picea rubens, and Betula alleghaniensis. Can. J. For. Res. 1987, 17, 976–990. [Google Scholar] [CrossRef]
- Agerer, R. Studies on ectomycorrhizae. X. Mycorrhizae formed by Cortinarius obtusus and C. venetus on spruce. Mycologia 1987, 79, 524–539. [Google Scholar] [CrossRef]
- Grelet, G.A.; Johnson, D.; Paterson, E.; Anderson, I.C.; Alexander, I.J. Reciprocal carbon and nitrogen transfer between an ericaceous dwarf shrub and fungi isolated from Piceirhiza bicolorata ectomycorrhizas. New Phytol. 2009, 182, 359–366. [Google Scholar] [CrossRef]
- Wen, Z.; Shi, L.; Tang, Y.; Hong, L.; Xue, J.; Xing, J.; Chen, Y.; Nara, K. Soil spore bank communities of ectomycorrhizal fungi in endangered Chinese Douglas-fir forests. Mycorrhiza 2018, 28, 49–58. [Google Scholar] [CrossRef]
- Furtak, K.; Grządziel, J.; Gałązka, A.; Niedźwiecki, J. Analysis of soil properties, bacterial community composition, and metabolic diversity in fluvisols of a floodplain area. Sustainability 2019, 11, 3929. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Chang, Z.z.; Zhao, J.t.; Zhou, M.g. Antifungal activity of Penicillium striatisporum Pst10 and its biocontrol effect on Phytophthora root rot of chilli pepper. Biol. Control 2008, 44, 24–31. [Google Scholar] [CrossRef]
- Rusman, Y.; Wilson, M.B.; Williams, J.M.; Held, B.W.; Blanchette, R.A.; Anderson, B.N.; Lupfer, C.R.; Salomon, C.E. Antifungal norditerpene oidiolactones from the fungus Oidiodendron truncatum, a potential biocontrol agent for White-Nose Syndrome in bats. J. Nat. Prod. 2020, 83, 344–353. [Google Scholar] [CrossRef]
- Ohtani, K.; Fujioka, S.; Shimada, A.; Kimura, Y. Nematicidal activities of 4-hydroxyphenylacetic acid and oidiolactone D produced by the fungus Oidiodendron sp. Z. Naturforschung C 2011, 66, 31–34. [Google Scholar] [CrossRef]
- Lou, H.; Guo, C.; Fan, B.; Fu, R.; Su, H.; Zhang, J.; Sun, L. Lingonberry (Vaccinium vitis-idaea L.) Interact With Lachnum pygmaeum to Mitigate Drought and Promote Growth. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Kwaśna, H.; Kotynska, U.; Łakomy, P.; Mallett, K. Stimulation of Armillaria rhizomorph formation by oak root fungi. Acta Mycol. 2001, 36, 257–272. [Google Scholar] [CrossRef] [Green Version]
- Schild, D.E.; Kennedy, A.; Stuart, M. Isolation of symbiont and associated fungi from ectomycorrhizas of Sitka spruce. For. Pathol. 1988, 18, 51–61. [Google Scholar] [CrossRef]
- Ważny, R.; Jędrzejczyk, R.J.; Rozpądek, P.; Domka, A.; Turnau, K. Biotization of highbush blueberry with ericoid mycorrhizal and endophytic fungi improves plant growth and vitality. Appl. Microbiol. Biotechnol. 2022, 106, 4775–4786. [Google Scholar] [CrossRef]
- Trappe, J.M. Mycorrhizal host and distribution of Cenococcum graniforme. Lloydia 1964, 27, 100–106. [Google Scholar]
- Molina, R.; Trappe, J.M. Patterns of ectomycorrhizal host specificity and potential among Pacific Northwest conifers and fungi. For. Sci. 1982, 28, 423–458. [Google Scholar]
- Trappe, J.M. Cenococcum Graniforme—Its Distribution, Ecology, Mycorrhiza Formation, and Inherent Variation; University of Washington: Seattle, WI, USA, 1962. [Google Scholar]
- Pigott, C. Fine structure of mycorrhiza formed by Cenococcum geophilum Fr. on Tilia cordata Mill. New Phytol. 1982, 92, 501–512. [Google Scholar] [CrossRef]
- Coleman, M.D.; Bledsoe, C.S.; Lopushinsky, W. Pure culture response of ectomycorrhizal fungi to imposed water stress. Can. J. Bot. 1989, 67, 29–39. [Google Scholar] [CrossRef]
- de Román, M.; de Miguel, A.M. Post-fire, seasonal and annual dynamics of the ectomycorrhizal community in a Quercus ilex L. forest over a 3-year period. Mycorrhiza 2005, 15, 471–482. [Google Scholar] [CrossRef]
- Torres, P.; Honrubia, M. Changes and effects of a natural fire on ectomycorrhizal inoculum potential of soil in a Pinus halepensis forest. For. Ecol. Manag. 1997, 96, 189–196. [Google Scholar] [CrossRef]
- Shaw, C.G., III; Sidle, R.C. Evaluation of planting sites common to a southeast Alaska clear-cut. II. Available inoculum of the ectomycorrhizal fungus Cenococcum geophilum. Can. J. For. Res. 1983, 13, 9–11. [Google Scholar] [CrossRef]
- Fernández-Toirán, L.; Águeda, B. Fruitbodies of Cenococcum geophilum. Mycotaxon 2007, 100, 109–114. [Google Scholar]
- Pigott, C. Survival of mycorrhiza formed by Cenococcum geophilum Fr. in dry soils. New Phytol. 1982, 92, 513–517. [Google Scholar] [CrossRef]
- Clericuzio, M.; Bivona, M.; Gamalero, E.; Bona, E.; Novello, G.; Massa, N.; Dovana, F.; Marengo, E.; Robotti, E. A Systematic Study of the Antibacterial Activity of Basidiomycota Crude Extracts. Antibiotics 2021, 10, 1424. [Google Scholar] [CrossRef]
- Beattie, K.D.; Rouf, R.; Gander, L.; May, T.W.; Ratkowsky, D.; Donner, C.D.; Gill, M.; Grice, I.D.; Tiralongo, E. Antibacterial metabolites from Australian macrofungi from the genus Cortinarius. Phytochemistry 2010, 71, 948–955. [Google Scholar] [CrossRef]
- Türkoğlu, A.; Duru, M.E.; Mercan, N. Antioxidant and antimicrobial activity of Russula delica Fr: An edidle wild mushroom. Eurasian J. Anal. Chem. 2007, 2, 54–67. [Google Scholar] [CrossRef]
- Chen, X.H.; Xia, L.X.; Zhou, H.B.; Qiu, G.Z. Chemical composition and antioxidant activities of Russula griseocarnosa sp. nov. J. Agric. Food Chem. 2010, 58, 6966–6971. [Google Scholar] [CrossRef]
- Nandi, A.K.; Samanta, S.; Maity, S.; Sen, I.K.; Khatua, S.; Devi, K.S.P.; Acharya, K.; Maiti, T.K.; Islam, S.S. Antioxidant and immunostimulant β-glucan from edible mushroom Russula albonigra (Krombh.) Fr. Carbohydr. Polym. 2014, 99, 774–782. [Google Scholar] [CrossRef]
- Zhou, J.; Gui, H.; Yang, S.; Yang, X.; Shi, L. Fungal interactions matter: Tricholoma matsutake domination affect fungal diversity and function in mountain forest soils. Biology 2021, 10, 1051. [Google Scholar] [CrossRef]
- Ohara, H.; Hamada, M. Disappearance of bacteria from the zone of active mycorrhizas in Tricholoma matsutake (S. Ito et Imai) Singer. Nature 1967, 213, 528–529. [Google Scholar] [CrossRef]
- Ogawa, M. Microbial ecology of mycorrhizal fungus, Tricholoma matsutake Sing. in pine forest 1. Fungal colony (“Shiro”) of T. matsutake. Bull. For. For. Prod. Res. Inst. 1975, 272, 79–121. [Google Scholar]
- Eto, S. Cultivation of the pine seedlings infected with Tricholoma matsutake by use of in vitro mycorrhizal synthesis. Bull. Hiroshima Prefect. For. Exp. Stn. 1990, 24, 1–6. [Google Scholar]
- Eto, S. Cultivation of the pine seedlings formed ectomycorrhizae with Tricholoma matsutake in plant culture flasks. Bull. Hiroshima Prefect. For. Exp. Stn. 1999, 24, 1–6. (In Japanese) [Google Scholar]
- Yamada, A.; Maeda, K.; Ohmasa, M. Ectomycorrhiza formation of Tricholoma matsutake isolates on seedlings of Pinus densiflora in vitro. Mycoscience 1999, 40, 455–463. [Google Scholar] [CrossRef]
- Guerin-Laguette, A.; Shindo, K.; Matsushita, N.; Suzuki, K.; Lapeyrie, F. The mycorrhizal fungus Tricholoma matsutake stimulates Pinus densiflora seedling growth in vitro. Mycorrhiza 2004, 14, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Yamada, A.; Furukawa, H. Advances in the cultivation of the highly-prized ectomycorrhizal mushroom Tricholoma matsutake. Mycoscience 2020, 61, 49–57. [Google Scholar] [CrossRef]
- Velmala, S.M.; Rajala, T.; Heinonsalo, J.; Taylor, A.F.; Pennanen, T. Profiling functions of ectomycorrhizal diversity and root structuring in seedlings of Norway spruce (Picea abies) with fast-and slow-growing phenotypes. New Phytol. 2014, 201, 610–622. [Google Scholar] [CrossRef]
- Jones, M.D.; Grenon, F.; Peat, H.; Fitzgerald, M.; Holt, L.; Philip, L.J.; Bradley, R. Differences in 15N uptake amongst spruce seedlings colonized by three pioneer ectomycorrhizal fungi in the field. Fungal Ecol. 2009, 2, 110–120. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Arhipova, N.; Gaitnieks, T.; Vulfa, L.; Nikolajeva, V.; Balasova, I. Estimation of factors influencing development of Heterobasidion annosum in Spruce stands. Proc. Latv. Univ. Agric. 2008. [Google Scholar]
- Gené, J.; Blanco, J.L.; Cano, J.; García, M.E.; Guarro, J. New filamentous fungus Sagenomella chlamydospora responsible for a disseminated infection in a dog. J. Clin. Microbiol. 2003, 41, 1722–1725. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Caballero, J.; Toni, P.; Garcia, I.; Martinez de Merlo, E.; Rollan, E.; Gonzalez, M.; Blanco, J. Disseminated mycoses in a dog by Paecilomyces sp. J. Vet. Med. Ser. A 2000, 47, 243–249. [Google Scholar] [CrossRef]
- Christen-Zaech, S.; Patel, S.; Mancini, A.J. Recurrent cutaneous Geomyces pannorum infection in three brothers with ichthyosis. J. Am. Acad. Dermatol. 2008, 58, S112–S113. [Google Scholar] [CrossRef]
- Gianni, C.; Caretta, G.; Romano, C. Skin infection due to Geomyces pannorum var. pannorum. Mycoses 2003, 46, 430–432. [Google Scholar] [CrossRef]
- Lorch, J.M.; Lindner, D.L.; Gargas, A.; Muller, L.K.; Minnis, A.M.; Blehert, D.S. A culture-based survey of fungi in soil from bat hibernacula in the eastern United States and its implications for detection of Geomyces destructans, the causal agent of bat white-nose syndrome. Mycologia 2013, 105, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Kokurewicz, T.; Ogórek, R.; Pusz, W.; Matkowski, K. Bats Increase the Number of Cultivable Airborne Fungi in the “Nietoperek” Bat Reserve in Western Poland. Microb. Ecol. 2016, 72, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Boekhout, T.; Akova, M.; Meis, J.F.; Cornely, O.A.; Lortholary, O.; study group, E.E.; ECMM. ESCMID and ECMM joint clinical guidelines for the diagnosis and management of rare invasive yeast infections. Clin. Microbiol. Infect. 2014, 20, 76–98. [Google Scholar] [CrossRef]
- Correa-Galeote, D.; Argiz, L.; Mosquera-Corral, A.; Del Rio, A.V.; Juarez-Jimenez, B.; Gonzalez-Lopez, J.; Rodelas, B. Structure of fungal communities in sequencing batch reactors operated at different salinities for the selection of triacylglyceride-producers from a fish-canning lipid-rich waste stream. New Biotechnol. 2022, 71, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Agger, W.A.; Andes, D.; Burgess, J.W. Exophiala jeanselmei infection in a heart transplant recipient successfully treated with oral terbinafine. Clin. Infect. Dis. 2004, 38, e112–e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcio, N.; Tiyomi, A.; Gloria, B.; Fernanda, S.; Revankar, S.G.; Brian, L.W.; Sutton, D.A.; Patterson, T.F. Nosocomial outbreak of Exophiala jeanselmei fungemia associated with contamination of hospital water. Clin. Infect. Dis. 2002, 34, 1475–1480. [Google Scholar]
- Badali, H.; Najafzadeh, M.; Esbroeck, M.v.; Enden, E.v.d.; Tarazooie, B.; Meis, J.; Hoog, G.d. The clinical spectrum of Exophiala jeanselmei, with a case report and in vitro antifungal susceptibility of the species. Med. Mycol. 2010, 48, 318–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, T.; Farris, S.; Sutherland, J. Penetration of Sitka spruce by the pathogenic fungus Caloscypha fulgens. Can. J. Bot. 1982, 60, 544–553. [Google Scholar] [CrossRef]
- Feng, B.; Wang, X.H.; Ratkowsky, D.; Gates, G.; Lee, S.S.; Grebenc, T.; Yang, Z.L. Multilocus phylogenetic analyses reveal unexpected abundant diversity and significant disjunct distribution pattern of the Hedgehog Mushrooms (Hydnum L.). Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Zhang, Y.Z.; Zhang, Y.H.; Guan, G.Y.; Zhang, K.P.; Li, H.J.; Wang, J.J. Mushroom poisoning from Inocybe serotina: A case report from Ningxia, northwest China with exact species identification and muscarine detection. Toxicon 2020, 179, 72–75. [Google Scholar] [CrossRef]
- Bas, C.; Kuyper, T.W.; Noordeloos, M.; Vellinga, E. Flora Agaricina Neerlandica; AA Balkema Publishers: Rotterdam, The Netherlands; Brookfield, VT, USA, 1988; Volume 1. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Huang, H.Y.; Yang, S.D.; Zeng, N.K.; Zhang, G.L.; Hu, Y.; Tang, L.P. Hygrophorus parvirussula sp. nov., a new edible mushroom from southwestern China. Phytotaxa 2018, 373, 139–146. [Google Scholar] [CrossRef]
- Naseer, A.; Khalid, A.N.; Healy, R.; Smith, M.E. Two new species of Hygrophorus from temperate Himalayan Oak forests of Pakistan. MycoKeys 2019, 56, 33. [Google Scholar] [CrossRef]
- Karadžić, D.; Milijašević, T. The most important parasitic and saprophytic fungi in Austrian pine and Scots pine plantations in Serbia. Glas. Šumar. Fak. 2008, 97, 147–170. [Google Scholar] [CrossRef]
- Santamaria, O.; Tejerina, L.; Pajares, J.; Diez, J. Effects of associated fungi Sclerophoma pythiophila and Cenangium ferruginosum on Gremmeniella abietina dieback in Spain. For. Pathol. 2007, 37, 121–128. [Google Scholar] [CrossRef]
- Kunca, A.; Leontovyč, R. Pines dieback caused by Cenangium ferruginosum Fr. in Slovakia in 2012. Folia Oecol. 2013, 40, 220–224. [Google Scholar]
- Stosiek, N.; Terebieniec, A.; Ząbek, A.; Młynarz, P.; Cieśliński, H.; Klimek-Ochab, M. N-phosphonomethylglycine utilization by the psychrotolerant yeast Solicoccozyma terricola M 3.1. 4. Bioorg. Chem. 2019, 93, 102866. [Google Scholar] [CrossRef]
- Aiello, D.; Sannino, C.; Giannoni, T.; Fabbrizi, G.; Gelosia, M.; Nicolini, A.; Turchetti, B.; Cotana, F.; Buzzini, P. Triacyl Glycerols from Yeast-Catalyzed Batch and Fed-Batch Bioconversion of Hydrolyzed Lignocellulose from Cardoon Stalks. Fermentation 2021, 7, 315. [Google Scholar] [CrossRef]
- Berning, B.; Jones, M.E.S.; Vieira, L.M. Revision of the European species of the genus Hincksina Norman, 1903 (Bryozoa, Cheilostomatida, Flustridae). Zootaxa 2021, 5081, 333–352. [Google Scholar] [CrossRef]
- Audouin, J. Explication sommaire des planches de polypes de I’Egypte et de la Syrie, publieers par Jules-Cesar Savigny. Descr. Égypte Hist. Nat. 1826, 1, 225–244. [Google Scholar]
- Harmelin, J.G.; Bishop, J.D.; Madurell, T.; Souto, J.; Jones, M.E.S.; Zabala, M. Unexpected diversity of the genus Collarina Jullien, 1886 (Bryozoa, Cheilostomatida) in the NE Atlantic-Mediterranean region: New species and reappraisal of C. balzaci (Audouin, 1826) and C. fayalensis Harmelin, 1978. Zoosystema 2019, 40, 385–418. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| ITS3-Mix2 | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGAACCAWCGATGAAGAACGCAG |
| ITS4-cwmix1 | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAATCCTCCGCTTAYTGATATGC |
| ITS4-cwmix2 | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAATCCTCCGCTTATTRATATGC |
| Genus | Fungi Relative Abundance (%) | ||||
|---|---|---|---|---|---|
| Control | Bark | Wood | Sawdust | Bark Compost | |
| Compost | Residues | Gravel | |||
| Amphinema | 0.00 | 5.01 | 0.00 | 0.00 | 0.00 |
| Apiotrichum | 0.00 | 0.10 | 0.08 | 0.00 | 1.02 |
| Cenangium | 0.10 | 0.09 | 0.40 | 0.10 | 2.01 |
| Cenococcum | 0.00 | 0.00 | 0.00 | 2.01 | 0.00 |
| Collarina | 0.06 | 0.00 | 1.01 | 0.00 | 0.00 |
| Cortinarius | 3.01 | 2.03 | 0.01 | 0.08 | 0.00 |
| Exophiala | 0.70 | 0.30 | 1.02 | 0.70 | 0.30 |
| Geomyces | 1.01 | 0.30 | 0.80 | 0.09 | 2.01 |
| Hyaloscypha | 1.10 | 4.00 | 0.21 | 1.03 | 0.20 |
| Hydnum | 0.30 | 0.20 | 2.03 | 0.00 | 0.00 |
| Hygrophorus | 0.00 | 0.10 | 0.90 | 0.00 | 1.04 |
| Inocybe | 0.90 | 5.02 | 2.00 | 0.00 | 7.01 |
| Meliniomyces | 6.10 | 0.40 | 1.01 | 8.02 | 2.00 |
| Mortierella | 2.03 | 2.00 | 0.80 | 1.01 | 0.52 |
| Oidiodendron | 6.00 | 5.00 | 7.00 | 5.03 | 2.03 |
| Penicillium | 5.02 | 10.01 | 6.03 | 7.01 | 4.04 |
| Russula | 0.10 | 0.10 | 9.04 | 5.04 | 1.00 |
| Sagenomella | 6.04 | 2.00 | 0.70 | 2.00 | 0.81 |
| Solicoccozyma | 1.03 | 0.60 | 0.30 | 0.51 | 0.40 |
| Tricholoma | 2.00 | 1.02 | 1.02 | 1.02 | 4.02 |
| Wilcoxina | 6.01 | 0.10 | 0.04 | 0.11 | 2.00 |
| Functional Group | |||
|---|---|---|---|
| Soil Condition | Mycorrizal | Saprotroph | Plant Pathogen |
| Control | 11 | 20 | 4 |
| Bark compost | 24 | 15 | 3 |
| Bark compost gravel | 16 | 14 | 2 |
| Sawdust | 12 | 14 | 3 |
| Wood residues | 21 | 31 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malewski, T.; Borowik, P.; Olejarski, I.; Berezovska, D.; Dyshko, V.; Behnke-Borowczyk, J.; Pusz, W.; Matic, S.; Oszako, T. Mycobiome of Post-Agricultural Soils 20 Years after Application of Organic Substrates and Planting of Pine Seedlings. Forests 2023, 14, 36. https://doi.org/10.3390/f14010036
Malewski T, Borowik P, Olejarski I, Berezovska D, Dyshko V, Behnke-Borowczyk J, Pusz W, Matic S, Oszako T. Mycobiome of Post-Agricultural Soils 20 Years after Application of Organic Substrates and Planting of Pine Seedlings. Forests. 2023; 14(1):36. https://doi.org/10.3390/f14010036
Chicago/Turabian StyleMalewski, Tadeusz, Piotr Borowik, Ireneusz Olejarski, Daria Berezovska, Valentyna Dyshko, Jolanta Behnke-Borowczyk, Wojciech Pusz, Slavica Matic, and Tomasz Oszako. 2023. "Mycobiome of Post-Agricultural Soils 20 Years after Application of Organic Substrates and Planting of Pine Seedlings" Forests 14, no. 1: 36. https://doi.org/10.3390/f14010036