

Occurrence Prediction of Western Conifer Seed Bug (Leptoglossus occidentalis: Coreidae) and Evaluation of the Effects of Climate Change on Its Distribution in South Korea Using Machine Learning Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ecological Characteristics of WCSB
2.2. Data Collection
2.3. Data Preprocessing and Analysis of Environmental Characteristics
2.4. Development of SDM for WCSB
3. Results
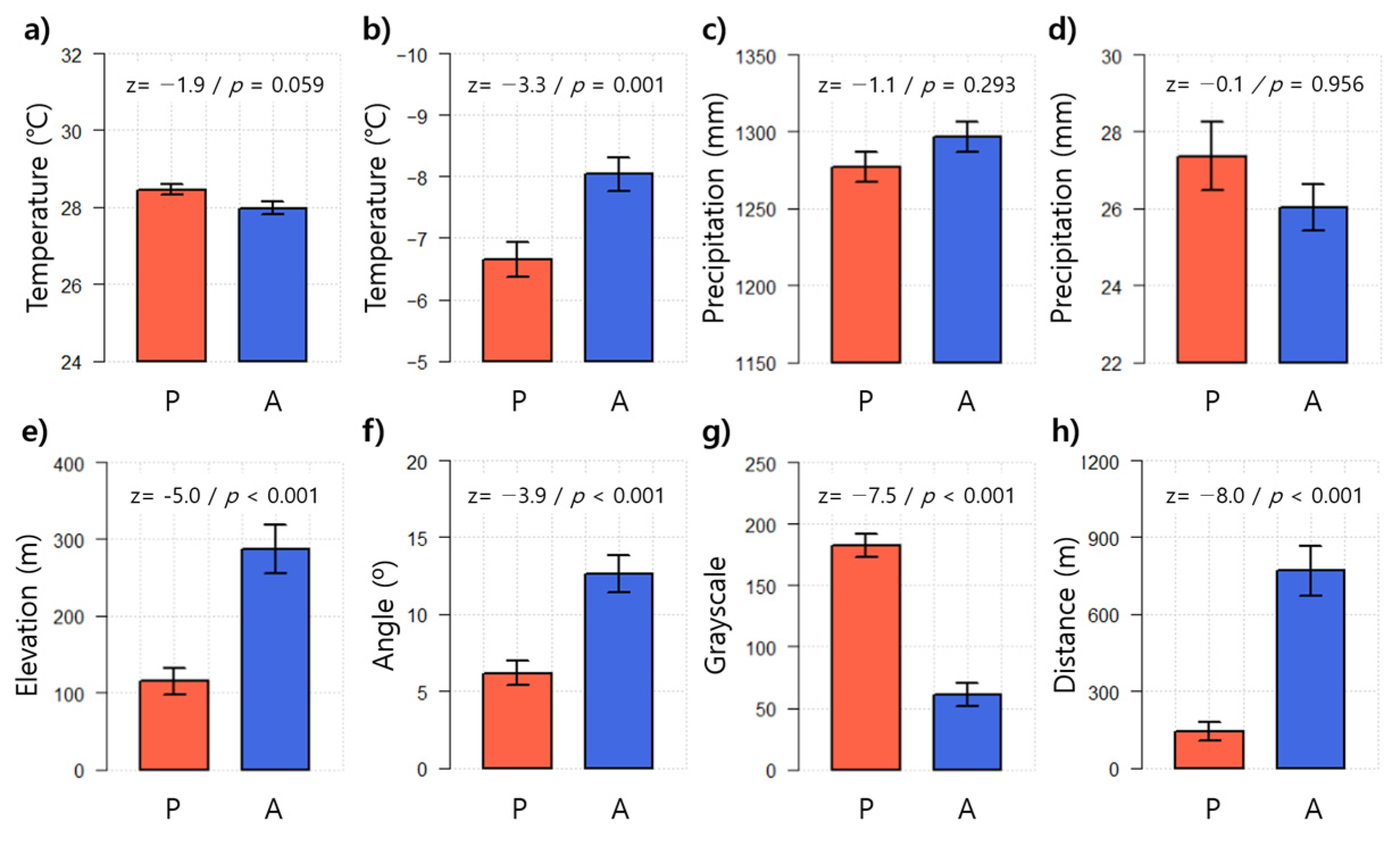
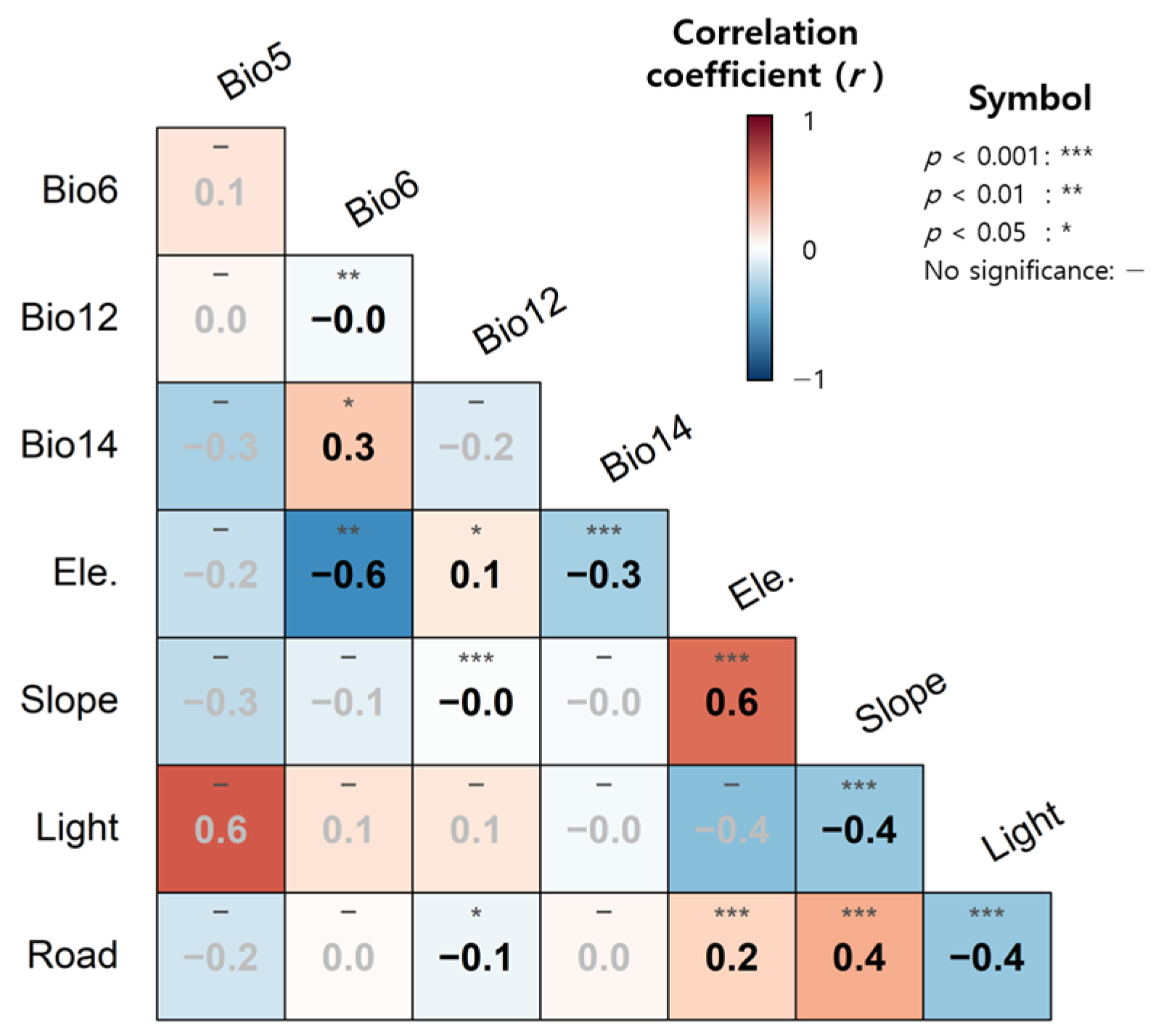
3.1. Environmental Characteristics of WCSB Occurrence
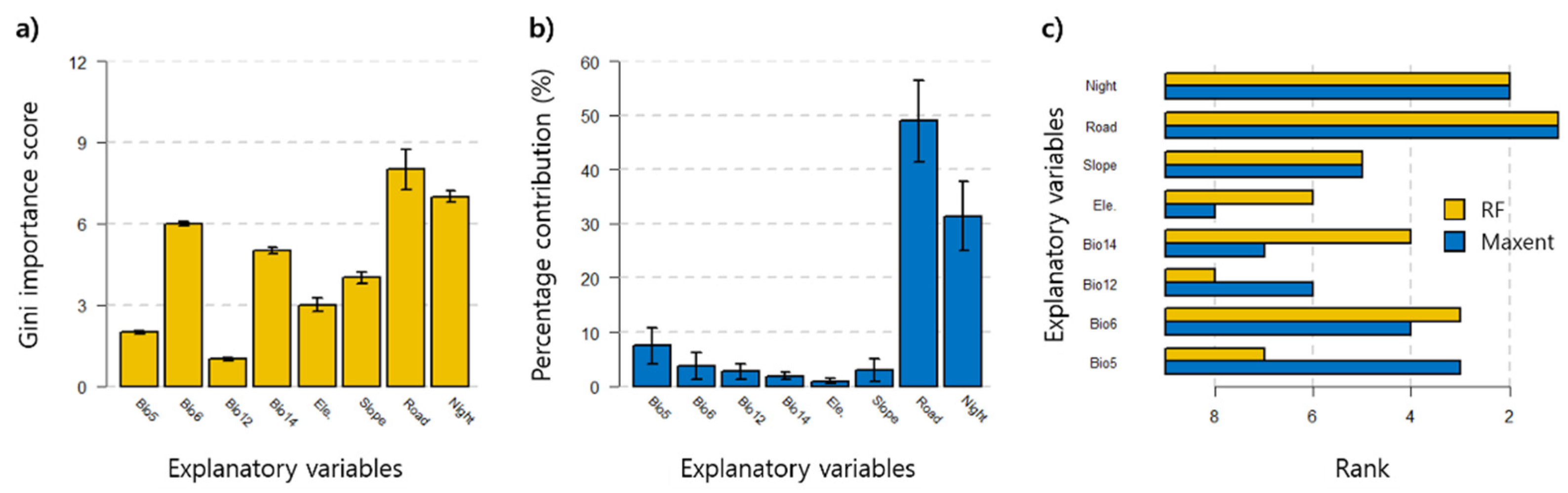
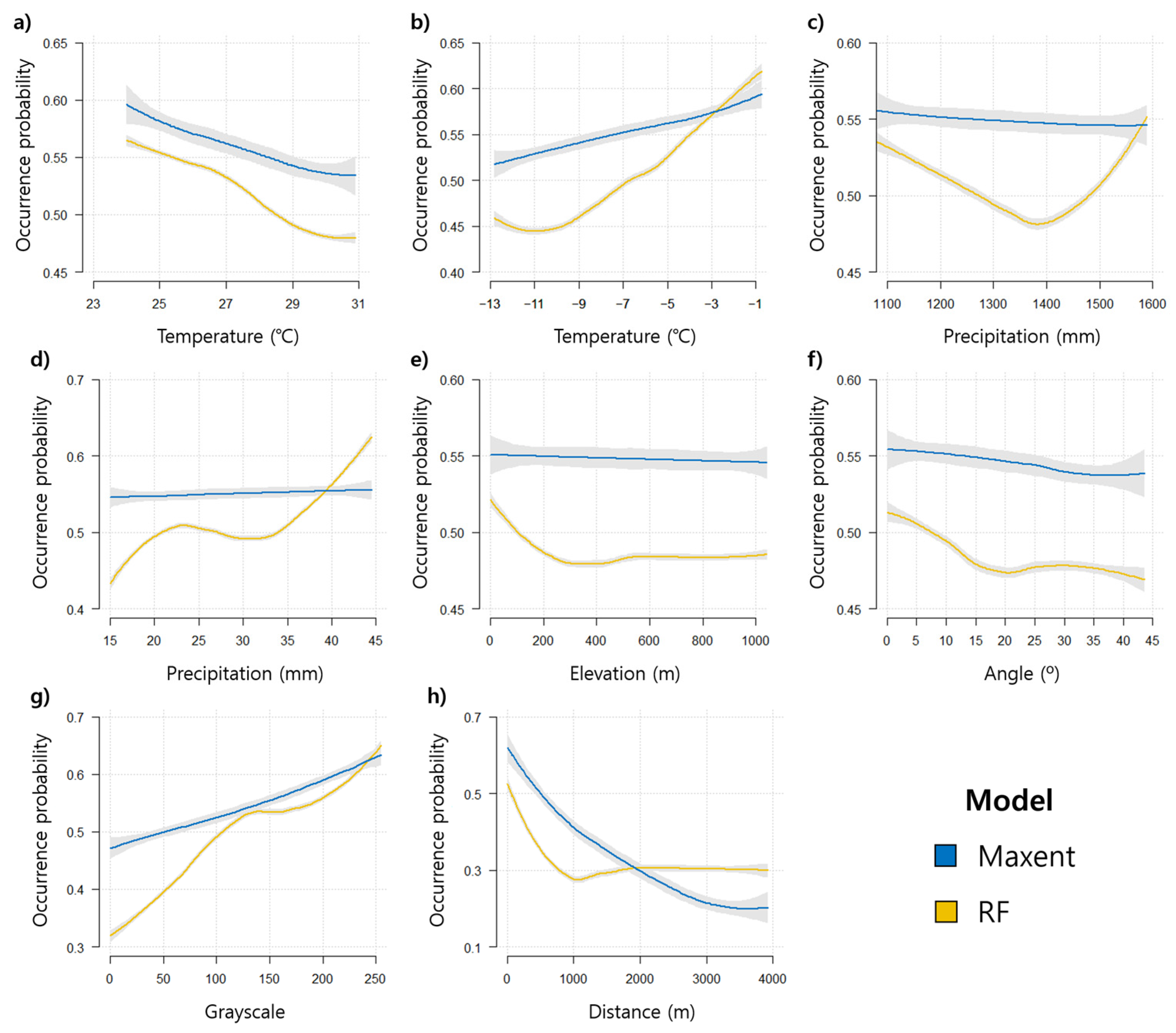
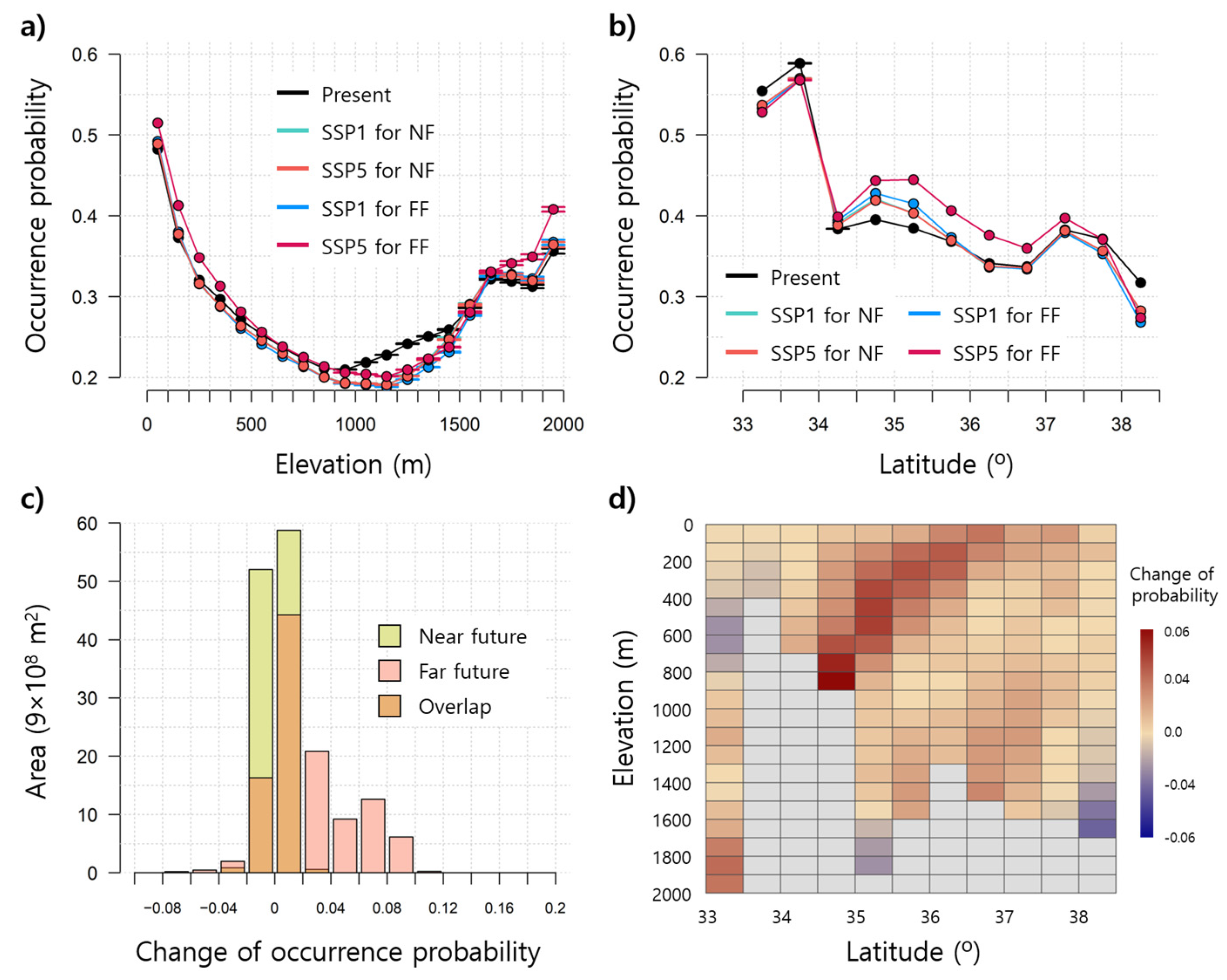
3.2. Prediction of WCSB Occurrence Using SDMs
4. Discussion
4.1. Occurrence and Dispersal Characteristics of WCSBs in South Korea
4.2. Interpretation of SDMs for WCSB Occurrence Prediction
4.3. Supporting the Management and Controls Strategies for Alien Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hughes, L. Biological consequences of global warming: Is the signal already. Trends Ecol. Evol. 2000, 15, 56–61. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group, Species Survival Commission, The International Union for Conservation of Nature: Auckland, New Zealand, 2000. [Google Scholar]
- Pyšek, P.; Richardson, D.M. Invasive species, environmental change and management, and health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture. Major Forest Insect and Disease Conditions in the United States 2015; U.S. Department of Agriculture: Washington, DC, USA, 2017.
- Lee, D.-S.; Bae, Y.-S.; Byun, B.-K.; Lee, S.; Park, J.K.; Park, Y.-S. Occurrence prediction of the citrus flatid planthopper (Metcalfa pruinosa (Say, 1830)) in South Korea using a random forest model. Forests 2019, 10, 583. [Google Scholar] [CrossRef] [Green Version]
- Liebhold, A.M.; Tobin, P.C. Population ecology of insect invasions and their management. Annu. Rev. Entomol. 2008, 53, 387–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherson, J.; Packauskas, R.; Taylor, S.; O’brien, M. Eastern range extension of Leptoglossus occidentalis with a key to Leptoglossus species of America north of Mexico (Heteroptera: Coreidae). Great Lakes Entomol. 1990, 23, 5. [Google Scholar]
- Kim, D.E.; Lee, H.; Kim, M.J.; Ban, Y.G.; Kim, D. Leptoglossus occidentalis (Hemiptera: Coreidae) occurrence, potential habitats, and COI diversity in South Korea. J. Asia-Pac. Biodivers. 2020, 13, 35–45. [Google Scholar] [CrossRef]
- Bates, S.L.; Borden, J.H. Life table for Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) and prediction of damage in lodgepole pine seed orchards. Agric. For. Entomol. 2005, 7, 145–151. [Google Scholar] [CrossRef]
- National Institute of Ecology. Investigating Ecological Risk of Alien Species (IV); National Institute of Ecology: Seocheon, Republic of Korea, 2017.
- Bates, S.L.; Borden, J.H.; Kermode, A.R.; Bennett, R.G. Impact of Leptoglossus occidentalis (Hemiptera: Coreidae) on Douglas-fir seed production. J. Econ. Entomol. 2000, 93, 1444–1451. [Google Scholar] [CrossRef]
- Koerber, T.W. Leptoglossus occidentalis (Hemiptera, Coreidae), a newly discovered pest of coniferous seed. Ann. Entomol. Soc. Am. 1963, 56, 229–234. [Google Scholar] [CrossRef]
- Park, K. Gyeonggi-do Forest Resources Research Institute. Decrease in Pine Nut Production in the Gapyeong Area. Reveal the Leptoglossus occidentalis. Media News, 8 October 2020. [Google Scholar]
- Beaury, E.M.; Fusco, E.J.; Jackson, M.R.; Laginhas, B.B.; Morelli, T.L.; Allen, J.M.; Pasquarella, V.J.; Bradley, B.A. Incorporating climate change into invasive species management: Insights from managers. Biol. Invasions 2020, 22, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, T. Climate change and its effects on terrestrial insects and herbivory patterns. Neotrop. Entomol. 2011, 40, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Lee, D.-S.; Kwon, T.-S.; Athar, M.; Park, Y.-S. Predicting the global distribution of Solenopsis geminata (Hymenoptera: Formicidae) under climate change using the MaxEnt model. Insects 2021, 12, 229. [Google Scholar] [CrossRef] [PubMed]
- Mainka, S.A.; Howard, G.W. Climate change and invasive species: Double jeopardy. Integr. Zool. 2010, 5, 102–111. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CAB Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- Drake, J.M.; Randin, C.; Guisan, A. Modelling ecological niches with support vector machines. J. Appl. Ecol. 2006, 43, 424–432. [Google Scholar] [CrossRef]
- Ahn, S.J.; Son, D.; Choo, H.Y.; Park, C.G. The first record on Leptoglossus occidentalis (Hemiptera: Coreidae) in Korea, a potential pest of the pinaceous tree species. J. Asia-Pac. Entomol. 2013, 16, 281–284. [Google Scholar] [CrossRef]
- National Institute of Environmental Research. A Photographic Guide to the Alien Insects and Their Host Plants in Korea; National Institute of Environmental Research: Incheon, Republic of Korea, 2013.
- Barta, M. Biology and temperature requirements of the invasive seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Europe. J. Pest Sci. 2016, 89, 31–44. [Google Scholar] [CrossRef]
- GBIF org. GBIF Occurrence Download. 2022. Available online: https://www.gbif.org/occurrence/download/0428875-210914110416597 (accessed on 5 September 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; p. 2391. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. QGIS Association. 2021. Available online: http://www.qgis.org (accessed on 1 May 2022).
- Zhu, G.-P.; Rédei, D.; Kment, P.; Bu, W.-J. Effect of geographic background and equilibrium state on niche model transferability: Predicting areas of invasion of Leptoglossus occidentalis. Biol. Invasions 2014, 16, 1069–1081. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Byeon, D.-H.; Jung, J.-M.; Park, Y.; Lee, H.-S.; Lee, J.-H.; Jung, S.; Lee, W.-H. Model-based assessment of changes in the potential distribution of Solenopsis geminata (Hymenoptera: Formicidae) according to climate change scenarios. J. Asia-Pac. Biodivers. 2020, 13, 331–338. [Google Scholar] [CrossRef]
- Kumar, S.; Yee, W.L.; Neven, L.G. Mapping global potential risk of establishment of Rhagoletis pomonella (Diptera: Tephritidae) using MaxEnt and CLIMEX niche models. J. Econ. Entomol. 2016, 109, 2043–2053. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 May 2022).
- Hijmans, R.J.; Raster: Geographic Data Analysis and Modeling. R package version 3.5-29. 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 1 May 2022).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling. R Package Version 1.3-5. 2017. Available online: https://CRAN.R-project.org/package=dismo (accessed on 1 May 2022).
- Sing, T.; Sander, O.; Beerenwinkel, N.; Lengauer, T. ROCR: Visualizing classifier performance in R. Bioinformatics 2005, 21, 3940–3941. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions, Version 3.4.1. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 1 May 2022).
- Choi, W.I.; Song, H.J.; Kim, D.S.; Lee, D.-S.; Lee, C.-Y.; Nam, Y.; Kim, J.-B.; Park, Y.-S. Dispersal patterns of pine wilt disease in the early stage of its invasion in South Korea. Forests 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Pantaleoni, R. The ways in which Metcalfa pruinosa (Say, 1830)(Auchenorrhyncha Flatidae) invades a new area. Boll. Dell’istituto DiEntomol. Guid. Grandi Della Univ. Degli Studi Di Bologna 1989, 43, 1–7. [Google Scholar]
- Lee, D.-S.; Choi, W.I.; Nam, Y.; Park, Y.-S. Predicting potential occurrence of pine wilt disease based on environmental factors in South Korea using machine learning algorithms. Ecol. Inform. 2021, 64, 101378. [Google Scholar] [CrossRef]
- Chen, S.; Ding, F.; Hao, M.; Jiang, D. Mapping the potential global distribution of red imported fire ant (Solenopsis invicta Buren) based on a machine learning method. Sustainability 2020, 12, 10182. [Google Scholar] [CrossRef]
- Husté, A.; Boulinier, T. Determinants of local extinction and turnover rates in urban bird communities. Ecol. Appl. 2007, 17, 168–180. [Google Scholar] [CrossRef]
- Molnar, C. Interpretable Machine Learning: A Guide for Making Black Box Models Explainable, 2nd ed.; Lulu: Morrisville, NC, USA, 2022. [Google Scholar]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]
- Régnière, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, D.; Wang, L.; Fang, J.; Sun, W.; Ren, B. Effects of altered precipitation on insect community composition and structure in a meadow steppe. Ecol. Entomol. 2014, 39, 453–461. [Google Scholar] [CrossRef]
- Olivera, L.; Minghetti, E.; Montemayor, S.I. Ecological niche modeling (ENM) of Leptoglossus clypealis a new potential global invader: Following in the footsteps of Leptoglossus occidentalis? Bull. Entomol. Res. 2021, 111, 289–300. [Google Scholar] [CrossRef] [PubMed]
- McGeoch, M.A.; Genovesi, P.; Bellingham, P.J.; Costello, M.J.; McGrannachan, C.; Sheppard, A. Prioritizing species, pathways, and sites to achieve conservation targets for biological invasion. Biol. Invasions 2016, 18, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, S.; Solís, D. Rapid Response Lowers Eradication Costs of Invasive Species. Choices 2018, 33, 1–9. [Google Scholar]
- Tamburini, M.; Maresi, G.; Salvadori, C.; Battisti, A.; Zottele, F.; Pedrazzoli, F. Adaptation of the invasive western conifer seed bug Leptoglossus occidentalis to Trentino, an alpine region (Italy). Bull. Insectology 2012, 65, 161–170. [Google Scholar]
- Byeon, D.-h.; Jung, J.-M.; Jung, S.; Lee, W.-H. Distribution analysis of Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) in South Korea using climate and host plant ensemble maps. J. Asia-Pac. Entomol. 2021, 24, 1077–1086. [Google Scholar] [CrossRef]
- Rehage, J.S.; Sih, A. Dispersal behavior, boldness, and the link to invasiveness: A comparison of four Gambusia species. Biol. Invasions 2004, 6, 379–391. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-S.; Lee, T.-G.; Bae, Y.-S.; Park, Y.-S. Occurrence Prediction of Western Conifer Seed Bug (Leptoglossus occidentalis: Coreidae) and Evaluation of the Effects of Climate Change on Its Distribution in South Korea Using Machine Learning Methods. Forests 2023, 14, 117. https://doi.org/10.3390/f14010117
Lee D-S, Lee T-G, Bae Y-S, Park Y-S. Occurrence Prediction of Western Conifer Seed Bug (Leptoglossus occidentalis: Coreidae) and Evaluation of the Effects of Climate Change on Its Distribution in South Korea Using Machine Learning Methods. Forests. 2023; 14(1):117. https://doi.org/10.3390/f14010117
Chicago/Turabian StyleLee, Dae-Seong, Tak-Gi Lee, Yang-Seop Bae, and Young-Seuk Park. 2023. "Occurrence Prediction of Western Conifer Seed Bug (Leptoglossus occidentalis: Coreidae) and Evaluation of the Effects of Climate Change on Its Distribution in South Korea Using Machine Learning Methods" Forests 14, no. 1: 117. https://doi.org/10.3390/f14010117