
Litter Decomposition of Qinghai Spruce (Picea crassifolia) Is Dependent on Mn Concentration in the Qilian Mountains, Northwest China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection and Preparation
2.3. Processing Litterbags
2.4. Chemical Analysis
2.5. Data Analysis
2.6. Statistical Analysis
3. Results
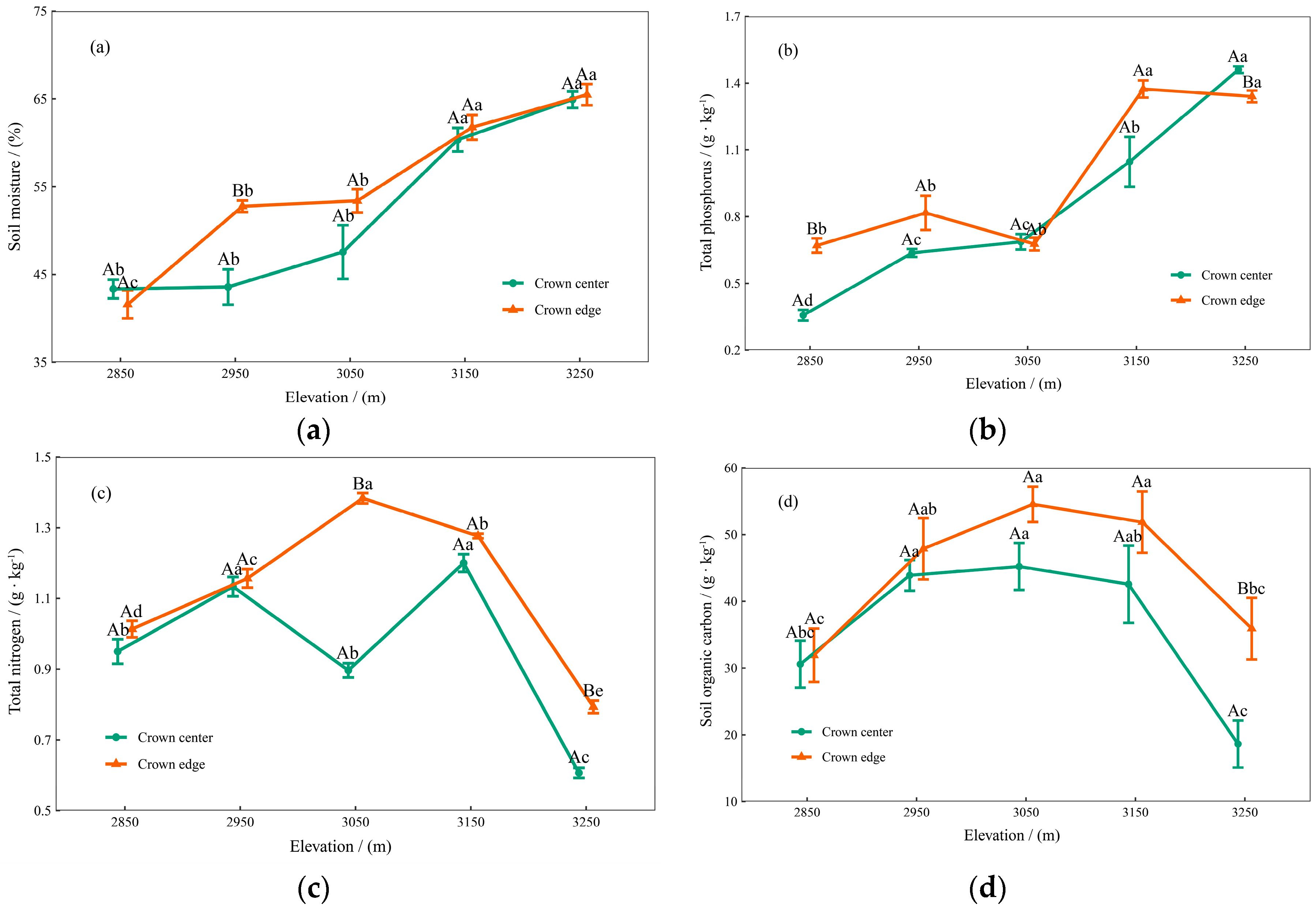
3.1. Soil Physical and Chemical Properties
3.2. Initial Chemical Composition of Qinghai Spruce Leaf Litter
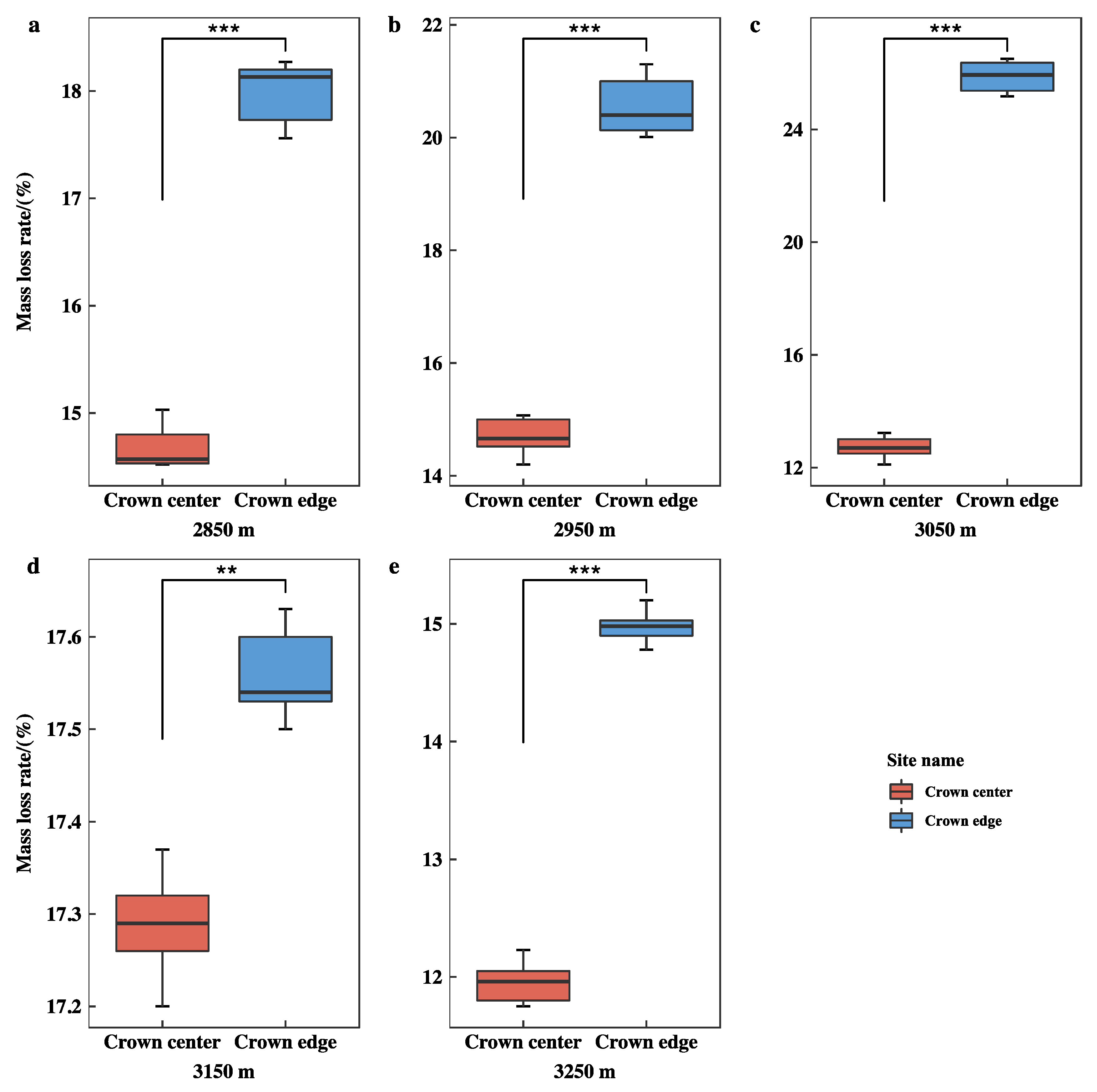
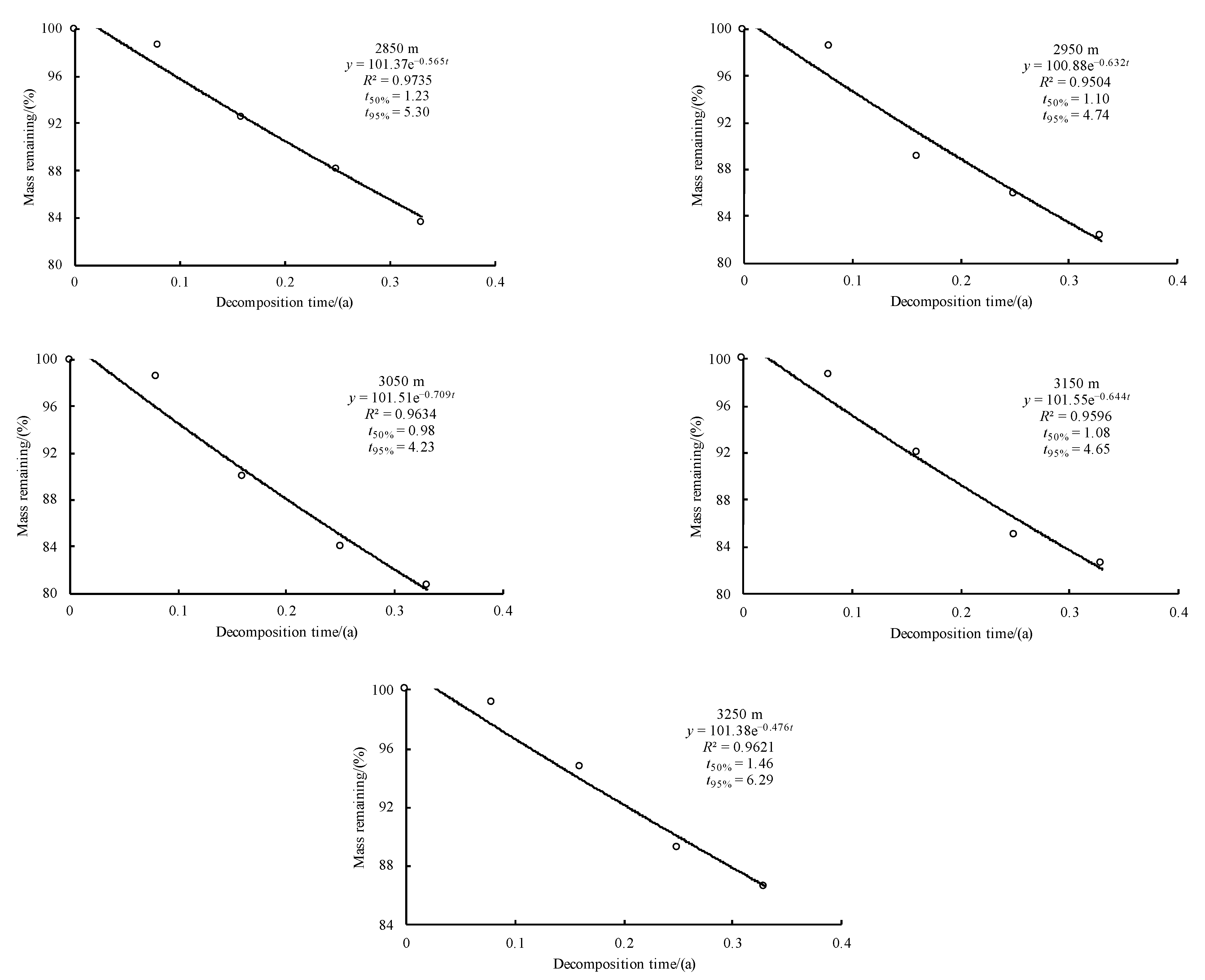
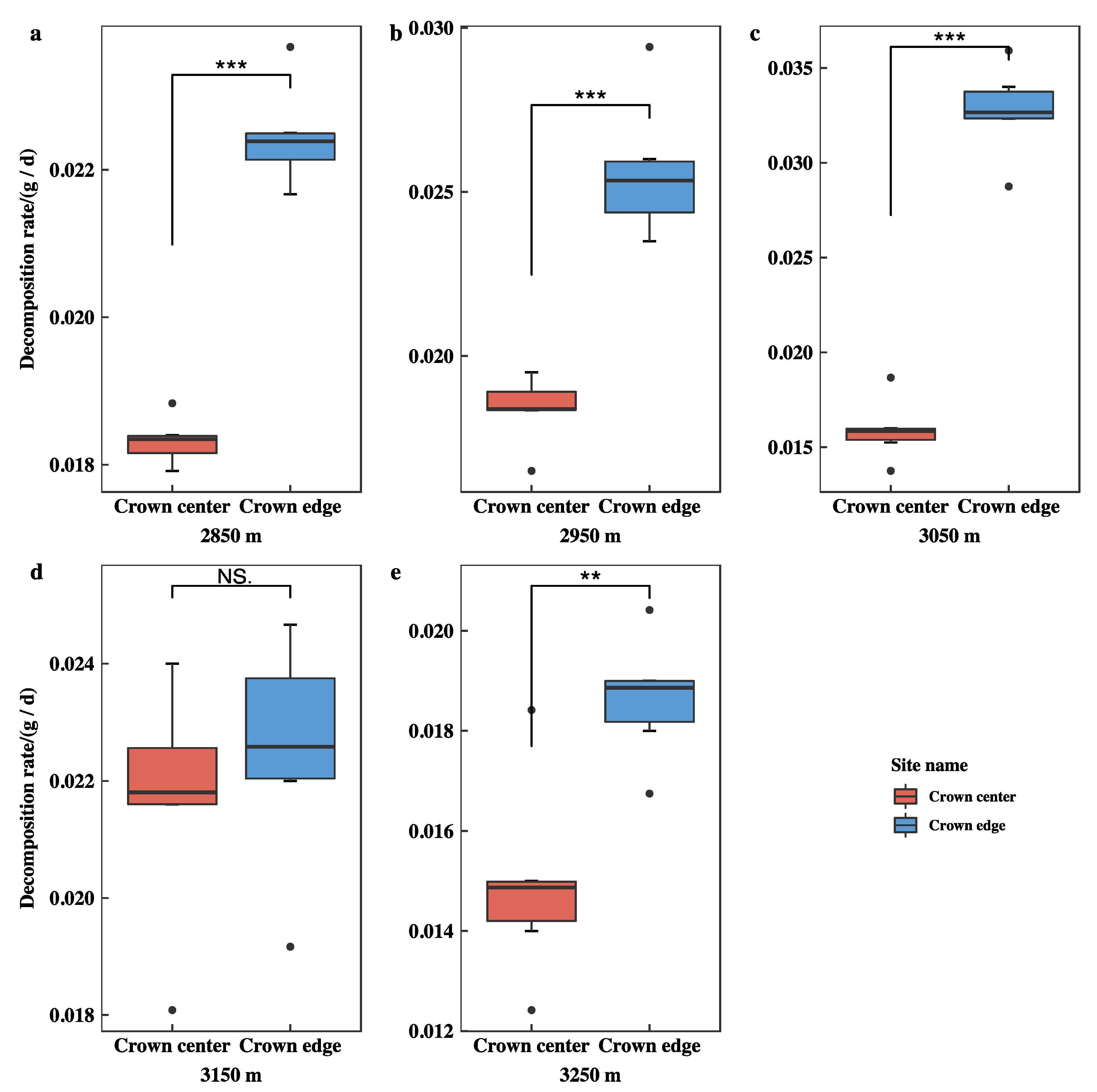
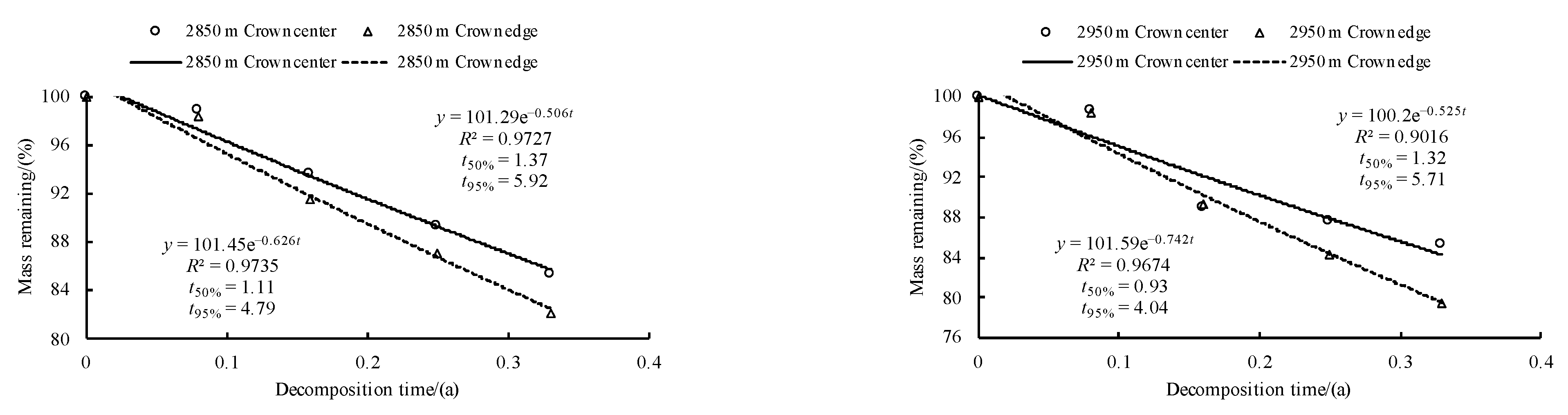
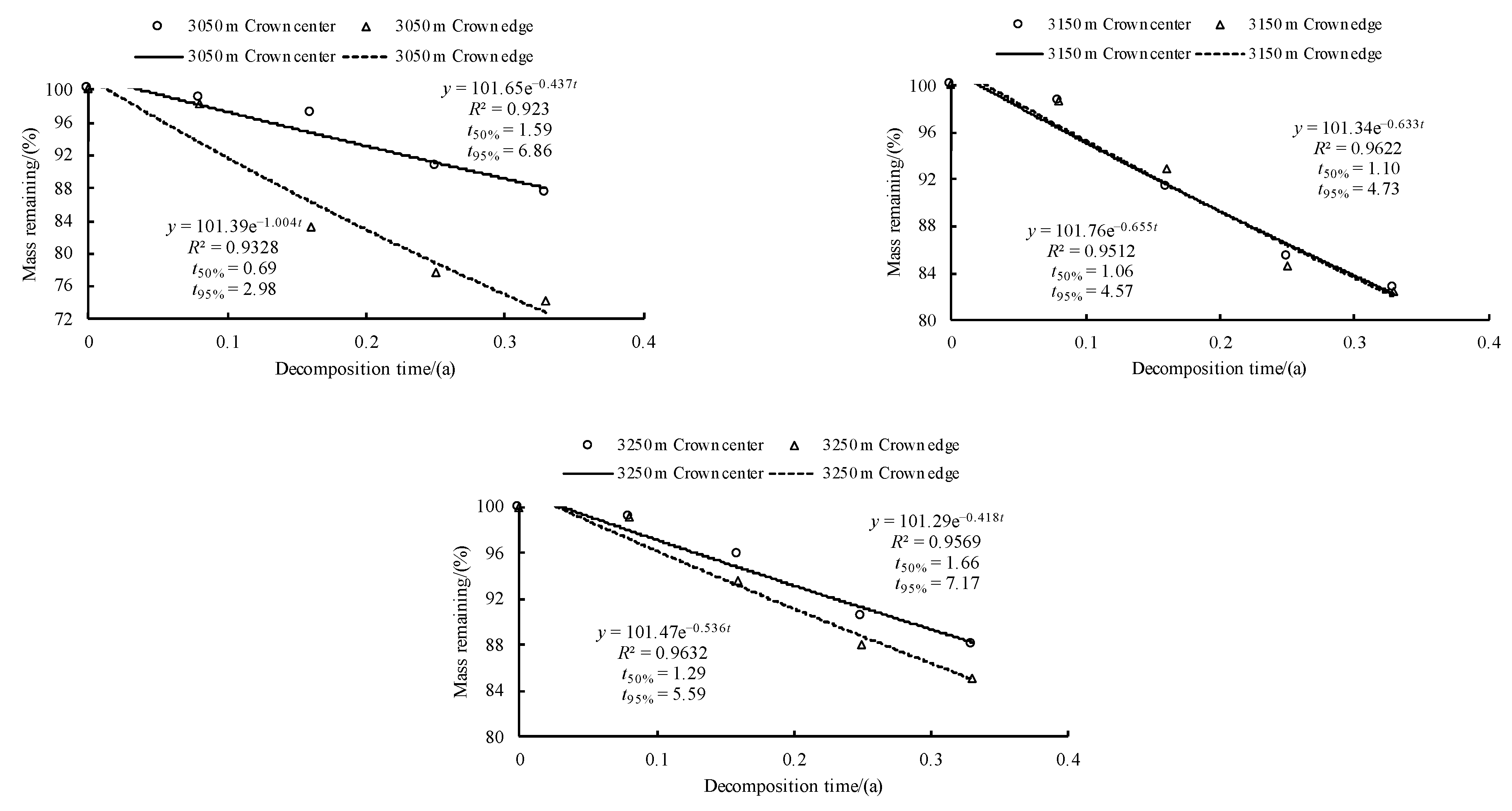
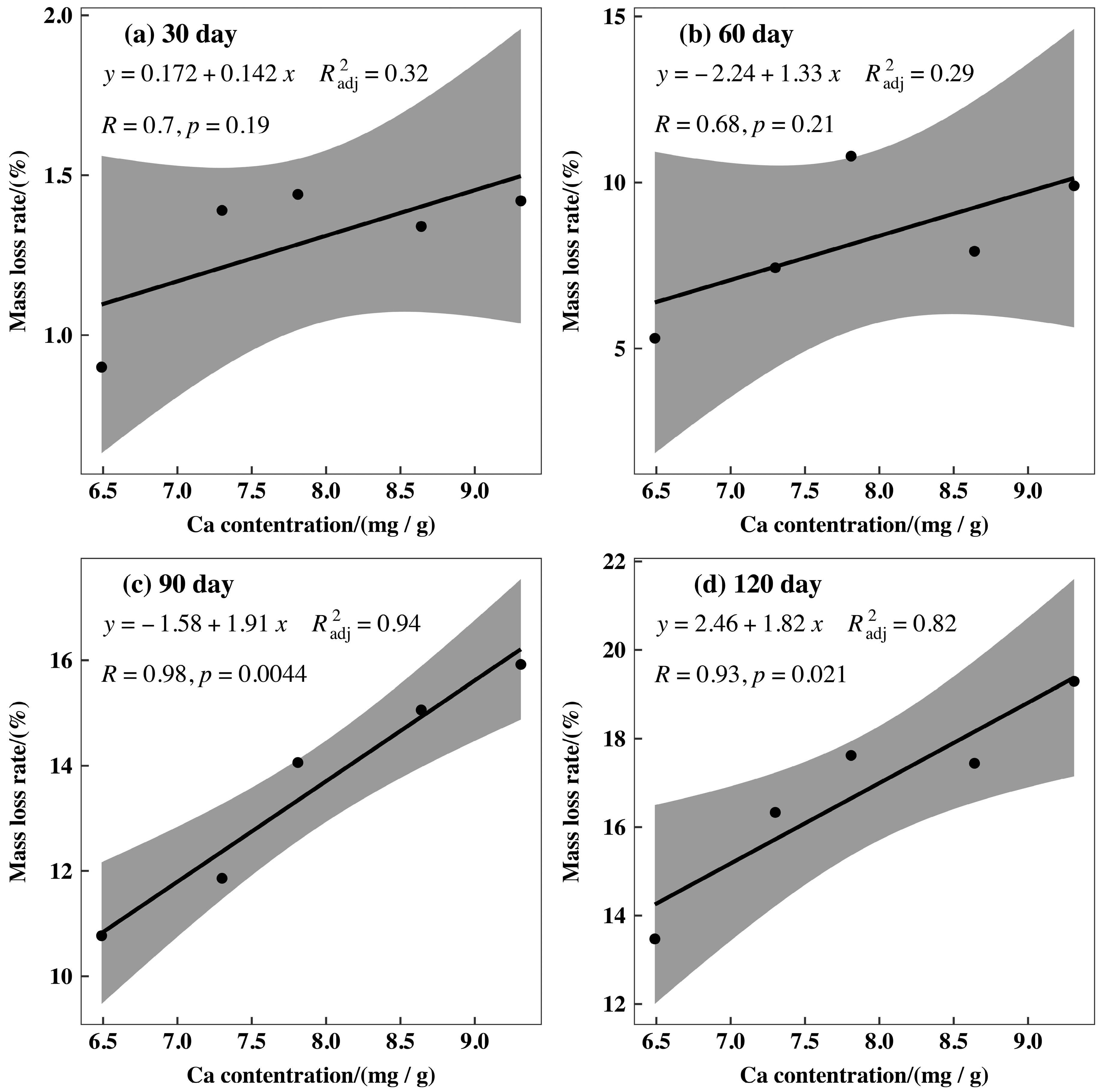
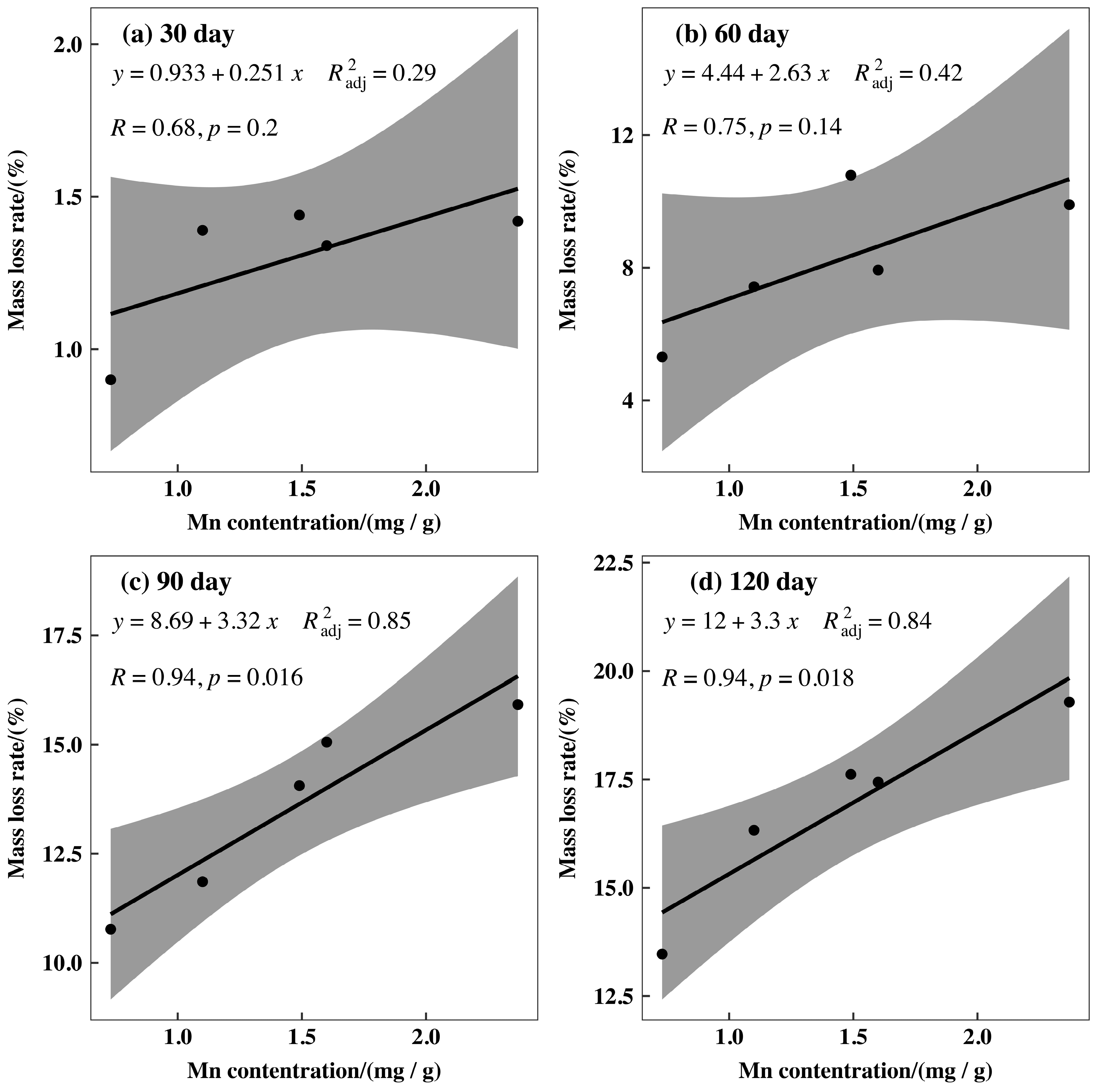
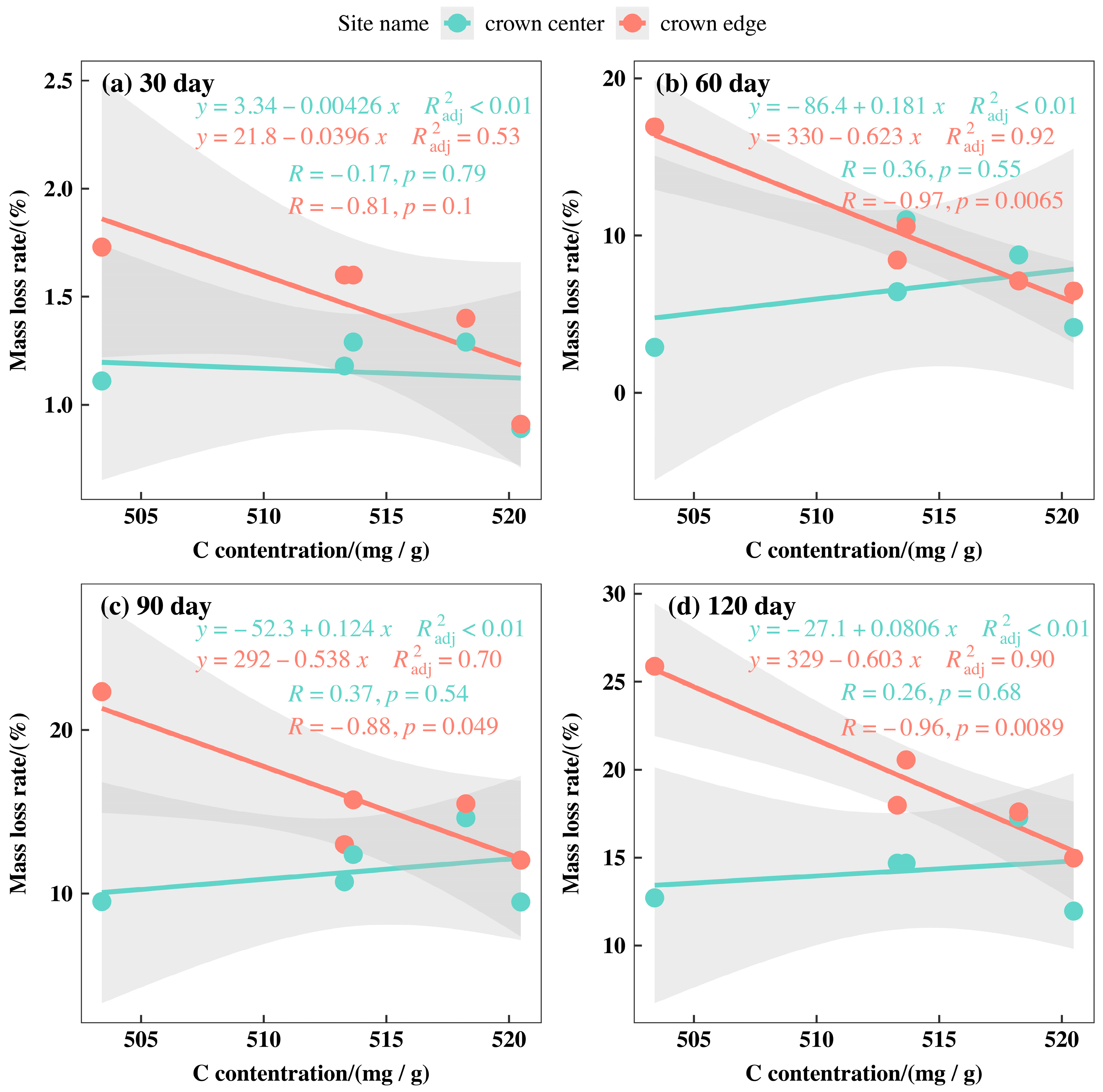
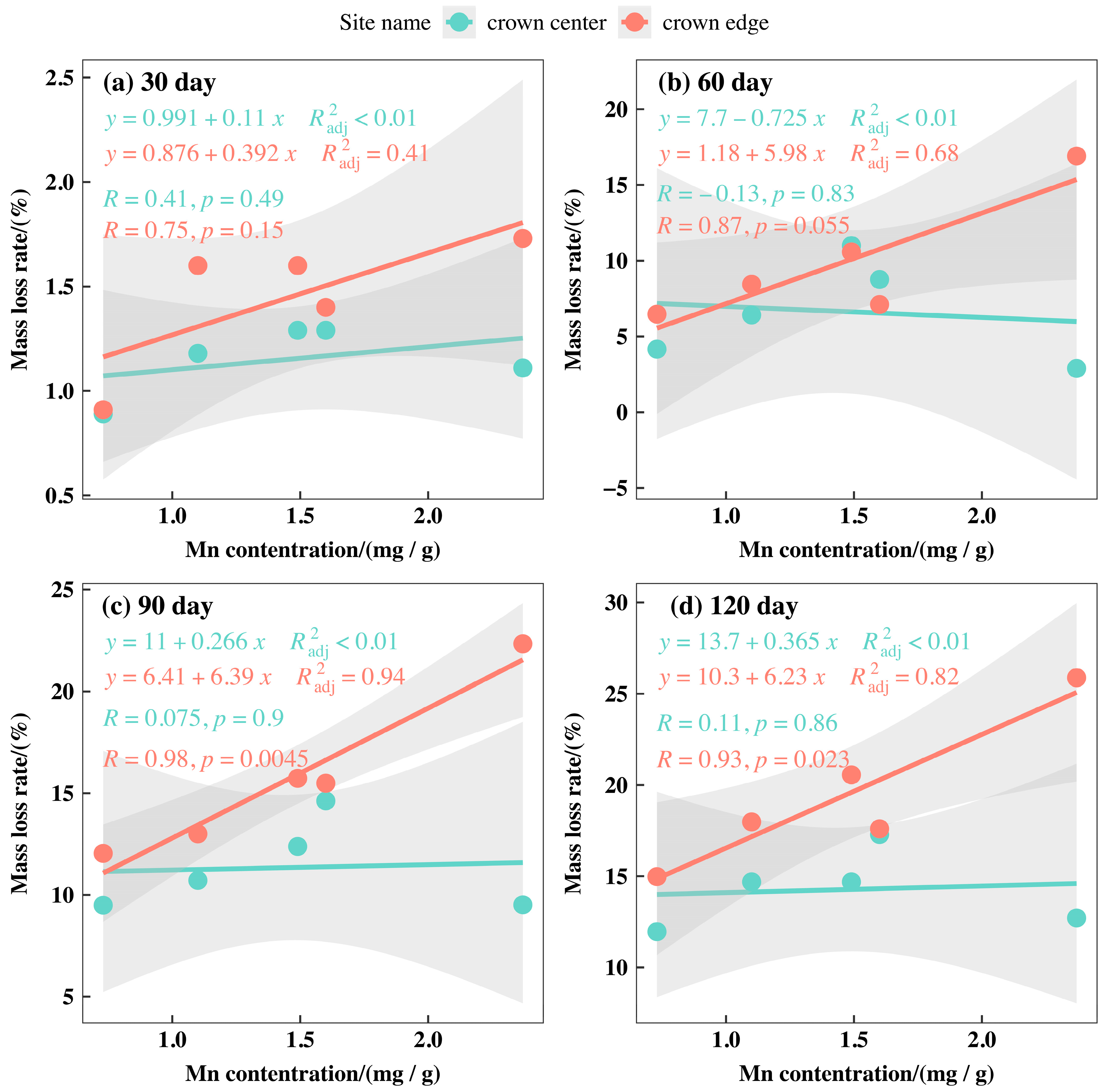
3.3. Initial Mass Loss Rates (%) of Qinghai Spruce Leaf Litter
3.4. Initial Decomposition Rates (g·day−1) of Qinghai Spruce Leaf Litter
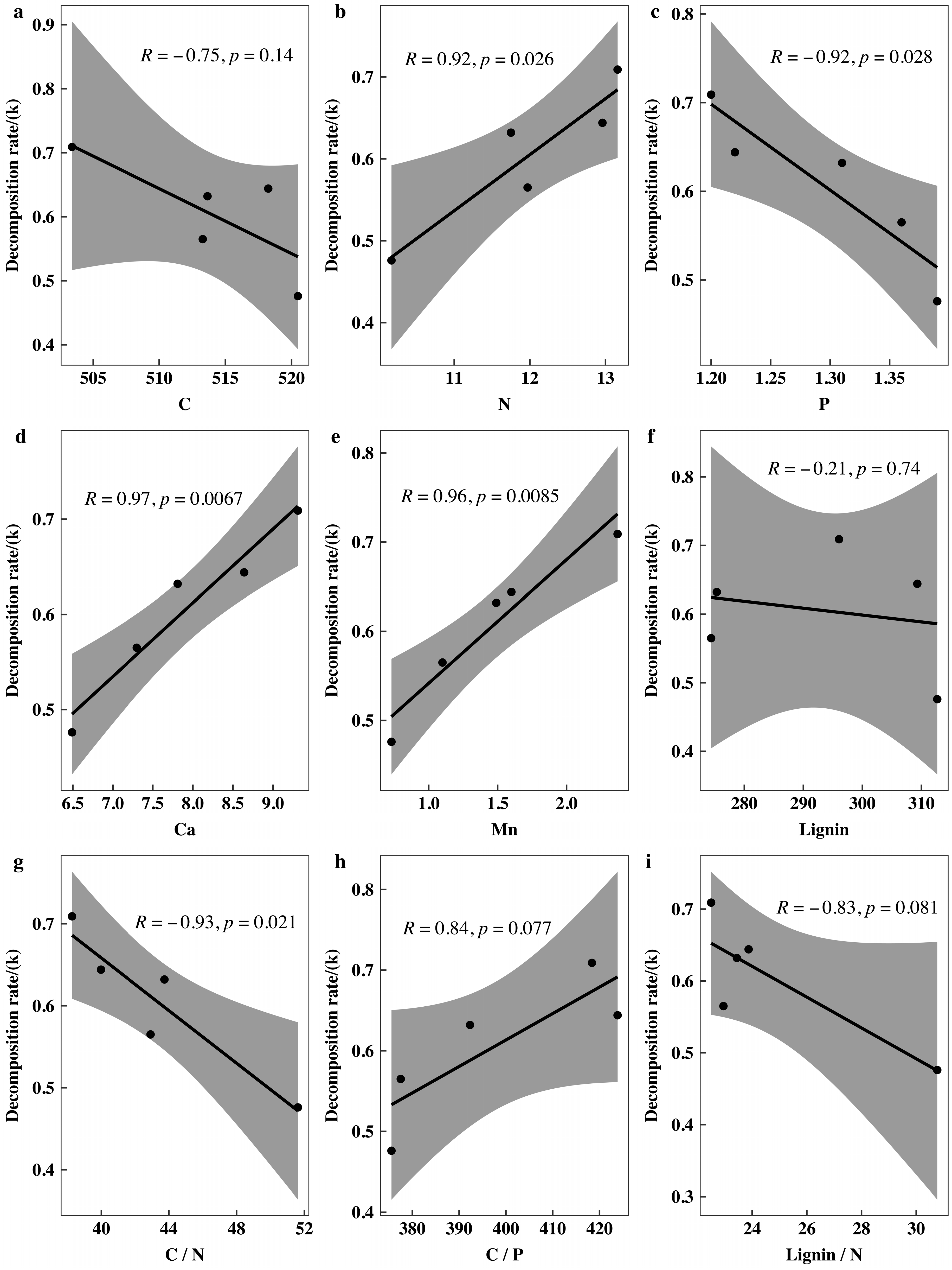
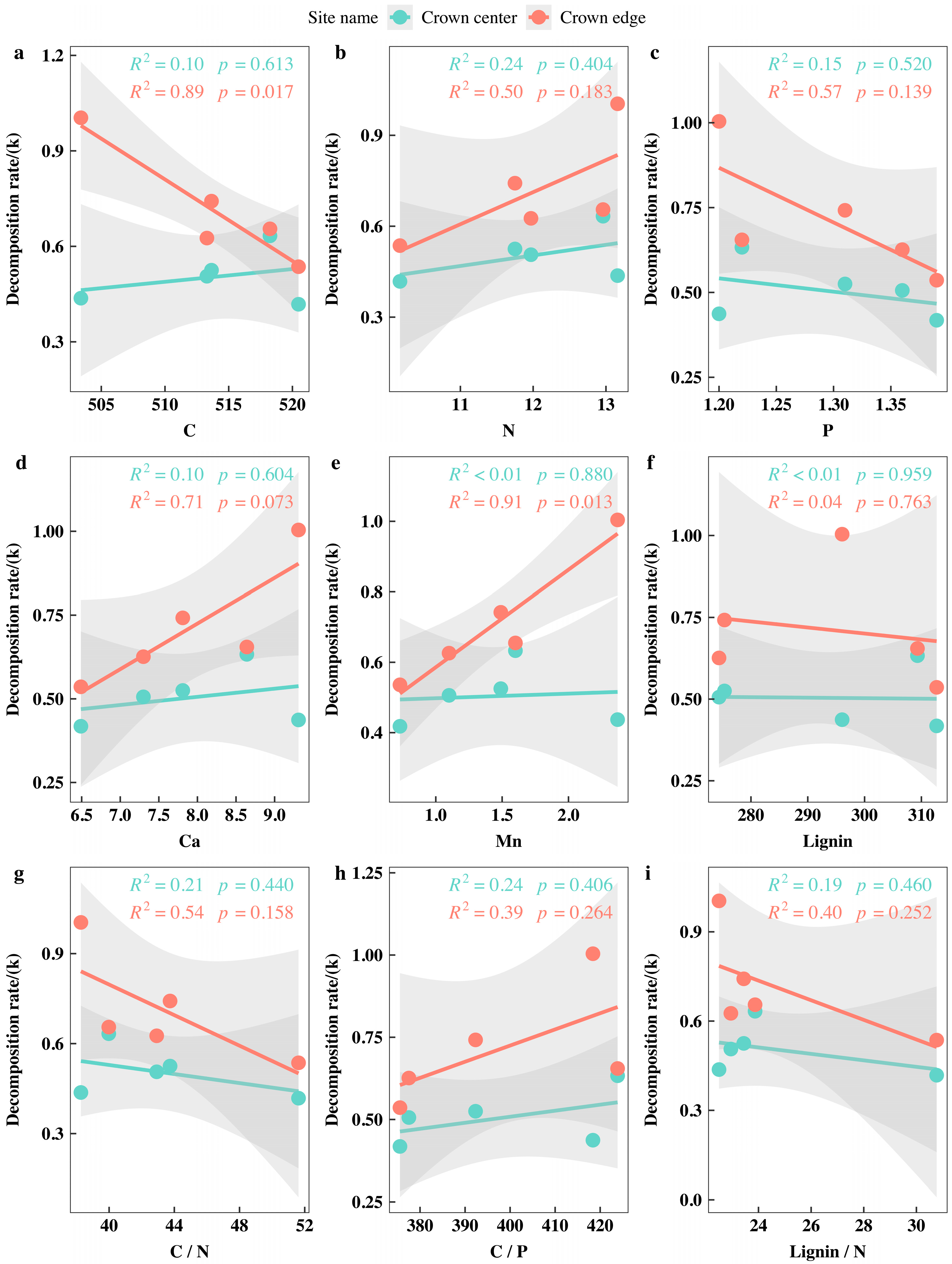
3.5. Correlations between the Initial Nutrient and Lignin Concentrations and Eco-Stoichiometric Ratios and the Initial Decomposition Rate (k)
4. Discussion
4.1. Initial Nutrient Concentration
4.2. Initial Mass Loss Rates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, S.; Liu, L.L.; Sayer, E.J. Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall-manipulation experiments. Biogeosciences 2013, 10, 5245–5272. [Google Scholar] [CrossRef]
- Liu, X.; Lin, T.C.; Vadeboncoeur, M.A.; Yang, Z.; Chen, S.; Xiong, D.; Xu, C.; Li, Y.; Yang, Y. Root litter inputs exert greater influence over soil C than does aboveground litter in a subtropical natural forest. Plant Soil 2019, 444, 489–499. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Dong, L.; Mao, Z. Simulated atmospheric nitrogen deposition alters decomposition of ephemeral roots. Ecosystems 2015, 18, 1240–1252. [Google Scholar] [CrossRef]
- Maroušek, J.; Trakal, L. Techno-economic analysis reveals the untapped potential of wood biochar. Chemosphere 2022, 291, 133000. [Google Scholar] [CrossRef]
- Mcclaugherty, C.A.; Pastor, J.; Aber, J.D.; Melillo, J.M. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 1985, 66, 266–275. [Google Scholar] [CrossRef]
- Upadhyay, V.P.; Singh, J.S. Nitrogen dynamics of decomposing hardwood leaf litter in a central himalayan forest. Soil Biol. Biochem. 1985, 17, 827–830. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in terrestrial ecosystems. Stud. Ecol. 1979, 5, 2772–2774. [Google Scholar] [CrossRef]
- Davey, M.P.; Berg, B.; Emmett, B.A.; Rowland, P. Decomposition of oak leaf litter is related to initial litter Mn concentrations. Can. J. Bot. 2007, 85, 16–24. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 1–14. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, F.; Wu, Q.; Kang, Z.; Yue, K.; Yang, Y.; Ni, X. Seasonal variations of non-structural carbohydrates in fresh litters of three dominant tree species in subtropical forests. Acta Ecol. Sin. 2022, 42, 1–10. [Google Scholar] [CrossRef]
- Berg, B.; Mcclaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Heidelberg/Berlin, Germany, 2013. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; He, J.; Feng, X. Iron-mediated soil carbon response to water-table decline in an alpine wetland. Nat. Commun. 2017, 8, 15972. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Tsuchida, H.; Matsumoto, T.; Makita, Y.; Kawashima, M.; Kikuchi, J.; Matsui, M. Screening of fungi for decomposition of lignin-derived products from Japanese cedar. J. Biosci. Bioeng. 2018, 126, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yin, X.; Kou, X.; Wang, Z.; Li, X.; Jiang, Y.; Wang, H.; Bernard, E.C. Effects of soil fauna on cellulose and lignin decomposition of plant litter in the Changbai Mountain, China. Environ. Entomol. 2019, 48, 592–602. [Google Scholar] [CrossRef]
- Eriksson, K.E.; Blanchette, R.A.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer: Heidelberg/Berlin, Germany, 1990. [Google Scholar] [CrossRef]
- Steffen, K.T.; Cajthaml, T.; Snajdr, J.; Baldrian, P. Differential degradation of oak (Quercus petraea) leaf litter by litter-decomposing basidiomycetes. Res. Microbiol. 2007, 158, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.R. Litter decomposition and its underlying mechanisms. Chinese J. Plant Ecol. 2019, 43, 648–657. [Google Scholar] [CrossRef]
- Li, N.; Zhao, C.; Hao, H.; Zang, F.; Chang, Y.; Wang, H.; Yang, J. Decomposition and its nutrients dynamic of Qinghai spruce leaf litter with elevation gradient in Qilian Mountains. Acta Ecol. Sin. 2021, 41, 4493–4502. [Google Scholar] [CrossRef]
- Zang, F.; Wang, H.; Zhao, C.; Nan, Z.; Wang, S.; Yang, J.; Li, N. Atmospheric wet deposition of trace elements to forest ecosystem of the Qilian Mountains, northwest China. Catena 2021, 197, 104966. [Google Scholar] [CrossRef]
- Li, N.; Zhao, C.; Zang, F.; Yang, J.; Hao, H.; An, J.; Zhang, M. Degradation dynamics of lignocellulose in the litterfall of Picea crassifolia in the Qilian Mountains. J. Lanzhou Univ. (Nat. Sci.) 2021, 57, 483–490. [Google Scholar] [CrossRef]
- Xie, H.; Ma, W.; Zhao, C.; Gao, Y.; Wang, Q.; Ge, H.; Lin, M.; Huang, H. Effects of litter and moss on soil respiration in a spruce forest in the Qilian Mountains, Qinghai. Acta Ecol. Sin. 2017, 37, 1379–1390. [Google Scholar] [CrossRef]
- Peng, S.; Zhao, C.; Xu, Z.; Ashiq, M.W. Restoration and conservation potential of destroyed Qinghai spruce (Picea crassifolia) forests in the Qilian Mountains of northwest China. Mitig. Adapt. Strat. Glob. Change 2016, 21, 153–165. [Google Scholar] [CrossRef]
- Gu, J.; Zhou, B.; Han, H.; Zhao, C. Temporal and spatial changes of microbial biomass in litter decomposition early process in Qinghai spruce (Picea crassifolia) forests. Acta Ecol. Sin. 2022, 43. [Google Scholar] [CrossRef]
- Briones, M.J.I. Soil fauna and soil functions: A jigsaw puzzle. Front. Environ. Sci. 2014, 2, 1–22. [Google Scholar] [CrossRef]
- Allen, S.E.; Grimshaw, H.M.; Parkinson, J.A.; Quarmby, C. Chemical analysis of ecological materials. J. Appl. Ecol. 1974, 13, 565–604. [Google Scholar] [CrossRef]
- Rowland, A.P.; Roberts, J.D. Lignin and cellulose fractions in decomposition studies using acid detergent fibre methods. Commun. Soil Sci. Plant Anal. 1994, 25, 269–277. [Google Scholar] [CrossRef]
- Wen, H.; Fu, H.; Guo, D. Influence of nitrogen addition on Stipa bungeana and Heteropappus altaicus litter decomposition and nutrient release in a steppe located on the Loess Plateau. Acta Ecol. Sin. 2017, 37, 2014–2022. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Barantal, S.; Schimann, H.; Fromin, N.; Hattenschwiler, S. C, N and P fertilization in an Amazonian rainforest supports stoichiometric dissimilarity as a driver of litter diversity effects on decomposition. Proc. R. Soc. B 2014, 281, 20141682. [Google Scholar] [CrossRef]
- Liu, J.; Fang, X.; Deng, Q.; Han, T.; Huang, W.; Li, Y. CO2 enrichment and N addition increase nutrient loss from decomposing leaf litter in subtropical model forest ecosystems. Sci. Rep. 2015, 5, 7952. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, J.; Zhang, Z.; Ren Fei Chen, L.; He, J. Changes in litter quality induced by nutrient addition alter litter decomposition in an alpine meadow on the Qinghai-Tibet Plateau. Sci. Rep. 2016, 6, 34290. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G. Litter mass-loss rates and decomposition patterns in some needle and leaf litter types. Long-term decomposition in a Scots pine forest. VII. Can. J. Bot. 1991, 69, 1449–1456. [Google Scholar] [CrossRef]
- Hatakka, A. Biodegradation of lignin. In Biopolymers, Vol. 1. Lignin, Humic Substances and Coal; Hofman, M., Stein, A., Eds.; Wiley: Weinheim, Germany, 2001; pp. 129–180. [Google Scholar] [CrossRef]
- Steffen, K.T.; Hatakka, A.; Hofrichter, M. Degradation of humic acids by the litter-decomposing basidiomycete Collybia dryophila. Appl. Environ. Microbiol. 2002, 68, 3442–3448. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.H.C. An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. J. Ecol. 1996, 84, 573–582. [Google Scholar] [CrossRef]
- Prescott, C.E. Do rates of litter decomposition tell us anything we really need to know? For. Ecol. Manag. 2005, 220, 66–74. [Google Scholar] [CrossRef]
- Berg, B.; Matzner, E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environ. Rev. 1997, 5, 1–25. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C.; Virzo De Santo, A.; Johnson, D. Humus buildup in boreal forests: Effects of litter fall and its N concentration. Can. J. For. Res. 2011, 31, 988–998. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
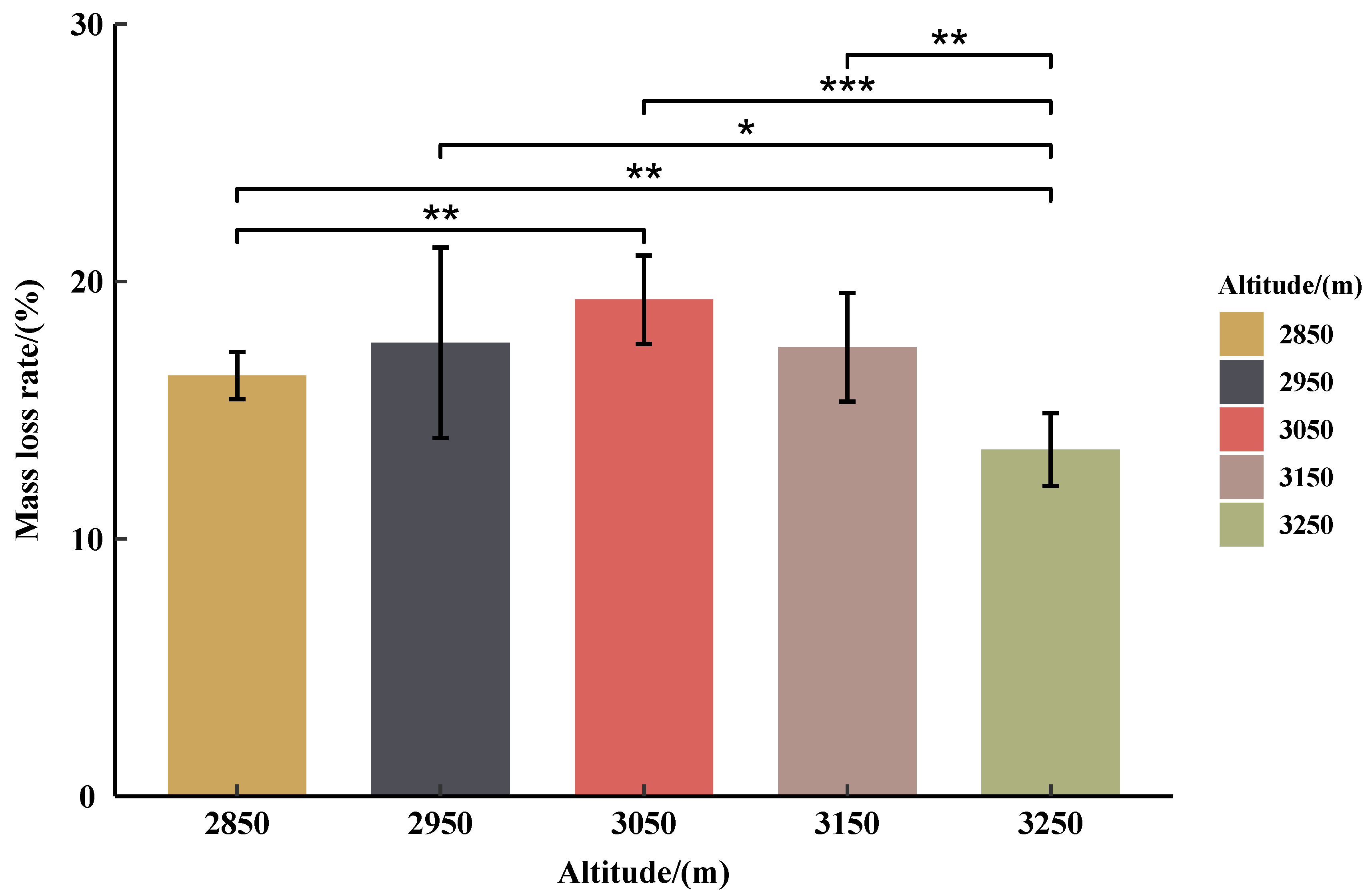{kind=link}
{kind=link}
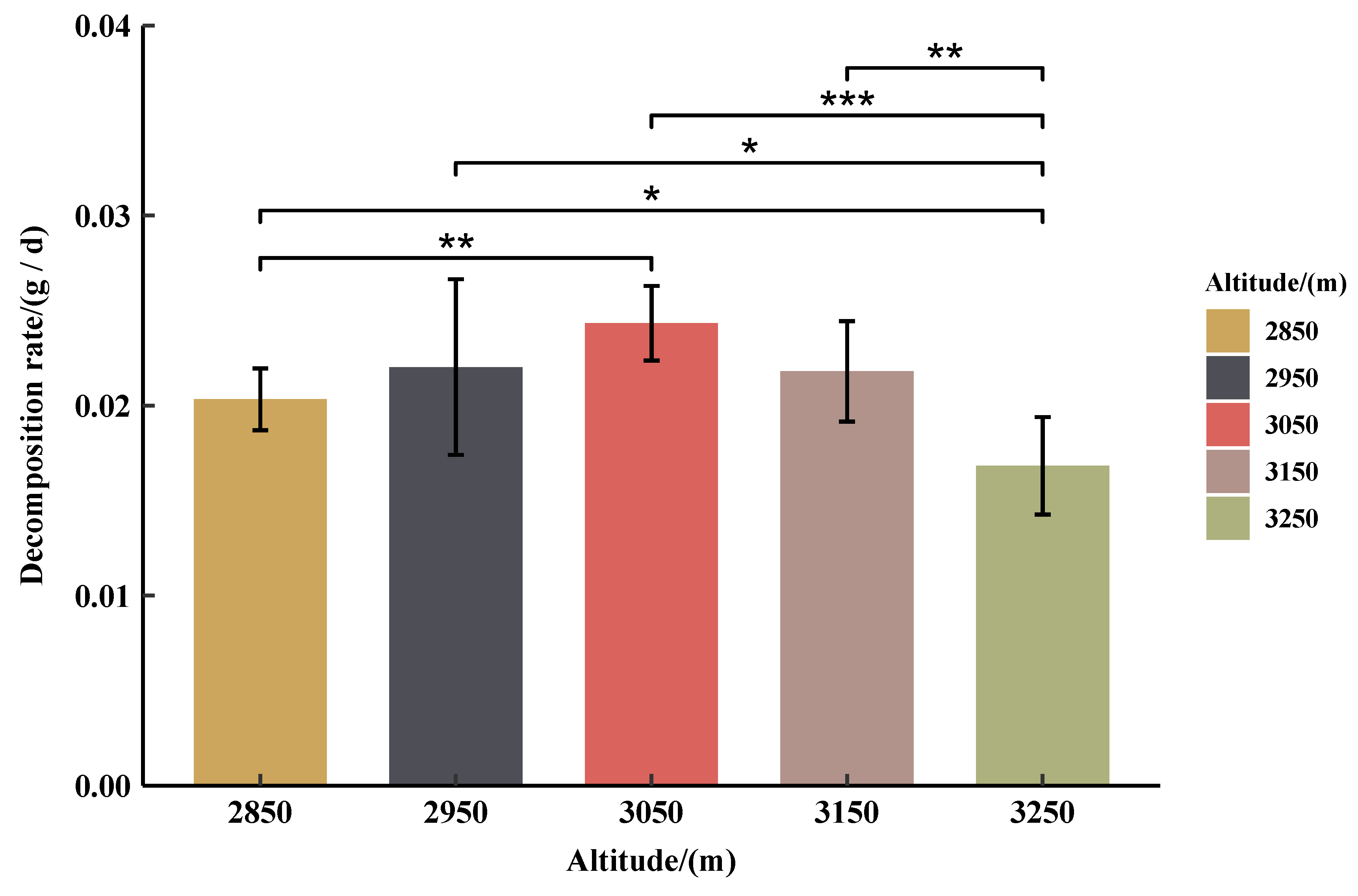{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (Altitude (m)) | Concentration (mg·g–1 DM) | Eco-Stoichiometric Ratio | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | N | P | Ca | Mn | Lignin | C/N | C/P | Lignin/N | |
| 2850 | 513.29 ± 3.66 ab | 11.97 ± 0.27 b | 1.36 ± 0.02 ab | 7.30 ± 0.12 d | 1.10 ± 0.08 c | 274.38 ± 1.75 c | 42.92 ± 0.68 b | 377.52 ± 3.34 c | 22.95 ± 0.40 b |
| 2950 | 513.65 ± 3.07 ab | 11.75 ± 0.16 b | 1.31 ± 0.03 b | 7.81 ± 0.11 c | 1.49 ± 0.03 b | 275.34 ± 1.53 c | 43.74 ± 0.32 b | 392.33 ± 5.85 bc | 23.44 ± 0.18 b |
| 3050 | 503.40 ± 2.44 b | 13.16 ± 0.29 a | 1.20 ± 0.01 c | 9.31 ± 0.15 a | 2.37 ± 0.10 a | 296.05 ± 3.37 b | 38.27 ± 0.66 c | 418.39 ± 3.03 ab | 22.50 ± 0.25 b |
| 3150 | 518.24 ± 6.49 a | 12.96 ± 0.23 a | 1.22 ± 0.03 c | 8.64 ± 0.21 b | 1.60 ± 0.08 b | 309.31 ± 2.87 a | 39.98 ± 0.21 c | 423.87 ± 4.94 a | 23.87 ± 0.29 b |
| 3250 | 520.48 ± 2.89 a | 10.17 ± 0.34 c | 1.39 ± 0.03 a | 6.49 ± 0.14 e | 0.73 ± 0.06 d | 312.69 ± 7.43 a | 51.26 ± 1.46 a | 375.54 ± 5.10 c | 30.77 ± 0.79 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Zhou, B.; Zhao, C.; Tang, Y.; Tian, J.; Zhao, X. Litter Decomposition of Qinghai Spruce (Picea crassifolia) Is Dependent on Mn Concentration in the Qilian Mountains, Northwest China. Forests 2022, 13, 1418. https://doi.org/10.3390/f13091418
Gu J, Zhou B, Zhao C, Tang Y, Tian J, Zhao X. Litter Decomposition of Qinghai Spruce (Picea crassifolia) Is Dependent on Mn Concentration in the Qilian Mountains, Northwest China. Forests. 2022; 13(9):1418. https://doi.org/10.3390/f13091418
Chicago/Turabian StyleGu, Jixiong, Bilian Zhou, Chuanyan Zhao, Yuan Tang, Junkai Tian, and Xinning Zhao. 2022. "Litter Decomposition of Qinghai Spruce (Picea crassifolia) Is Dependent on Mn Concentration in the Qilian Mountains, Northwest China" Forests 13, no. 9: 1418. https://doi.org/10.3390/f13091418