
Morphophysiological Acclimation of Developed and Senescing Beech Leaves to Different Light Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Biochemical Analyses
2.2.1. Leaf Pigments
2.2.2. Oxidative Stress Parameters
Hydrogen Peroxide
Thiobarbituric Acid-Reactive Substances
2.2.3. The Activity of the Antioxidant System
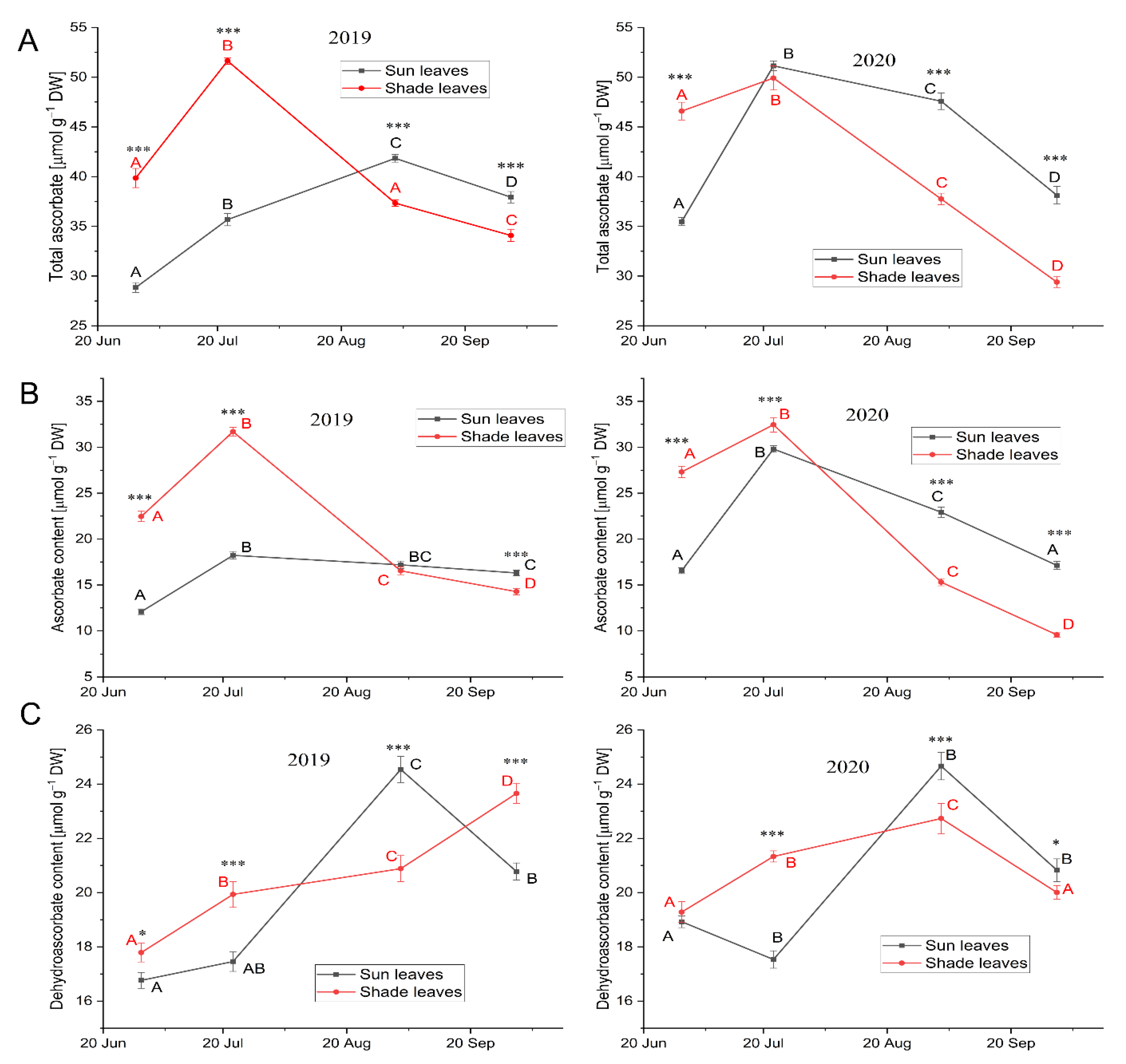
Ascorbate and Dehydroascorbate
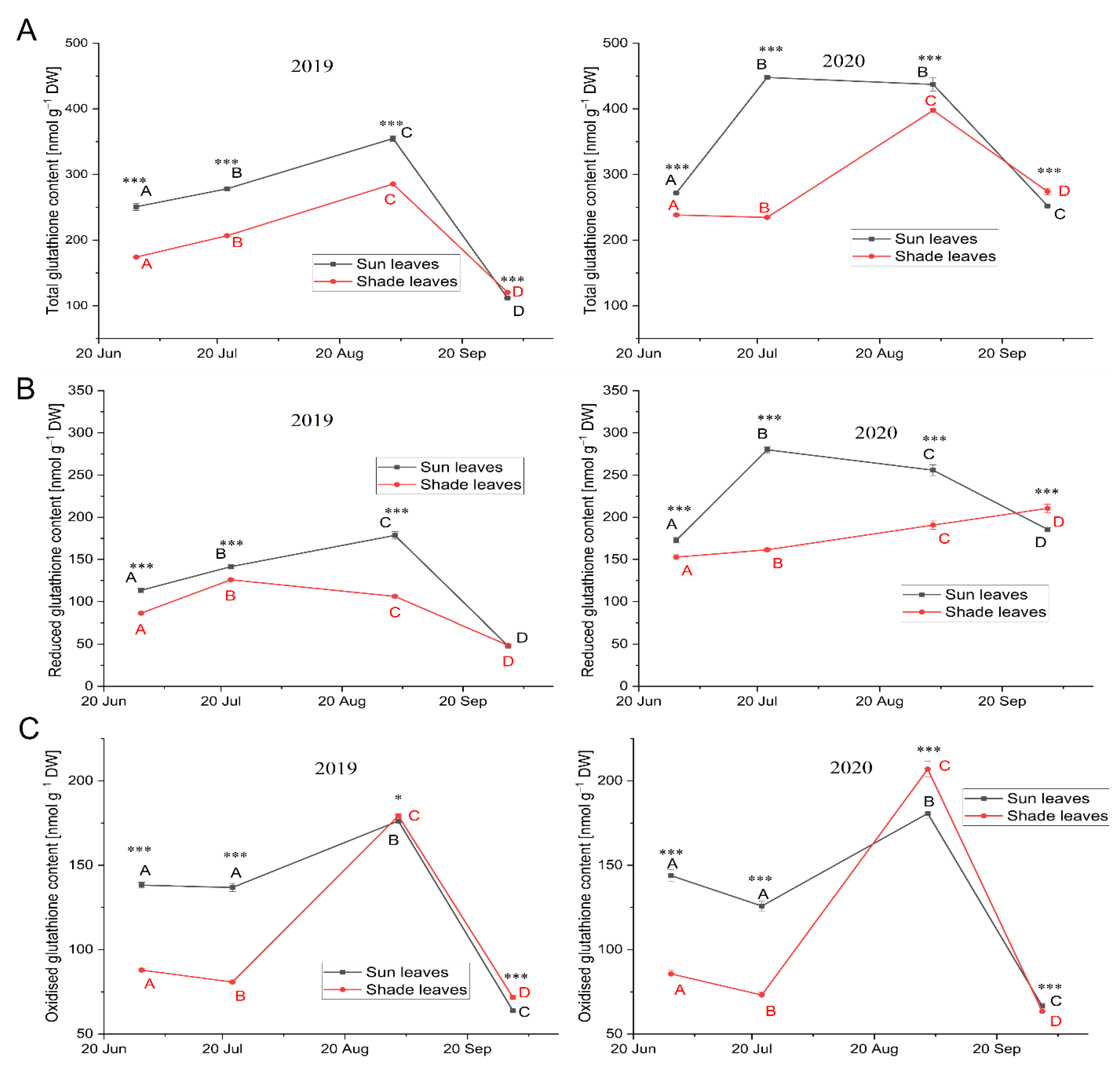
Glutathione and Glutathione Disulfide
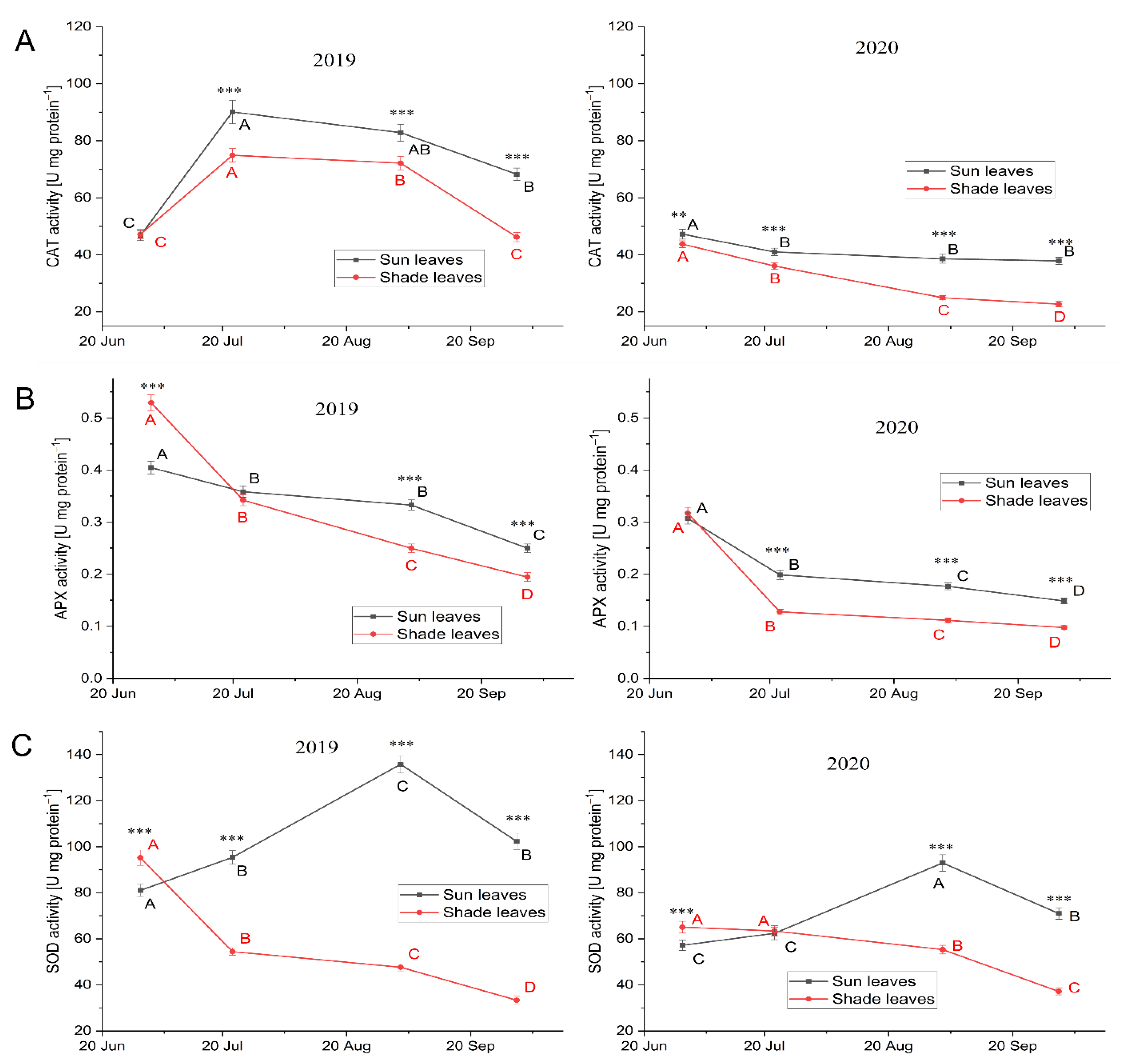
Extraction and Activity of Antioxidative Enzymes
Catalase
Ascorbate Peroxidase
Superoxide Dismutase
2.2.4. Data Analysis
3. Results
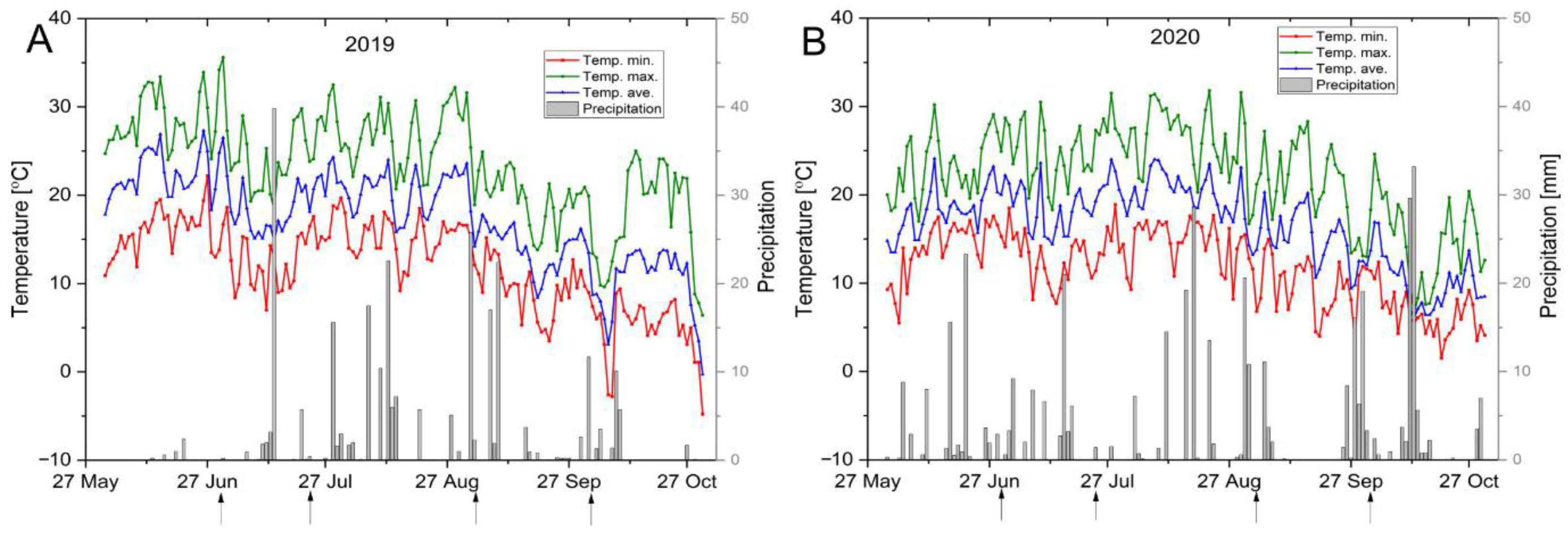
3.1. Climatic Conditions
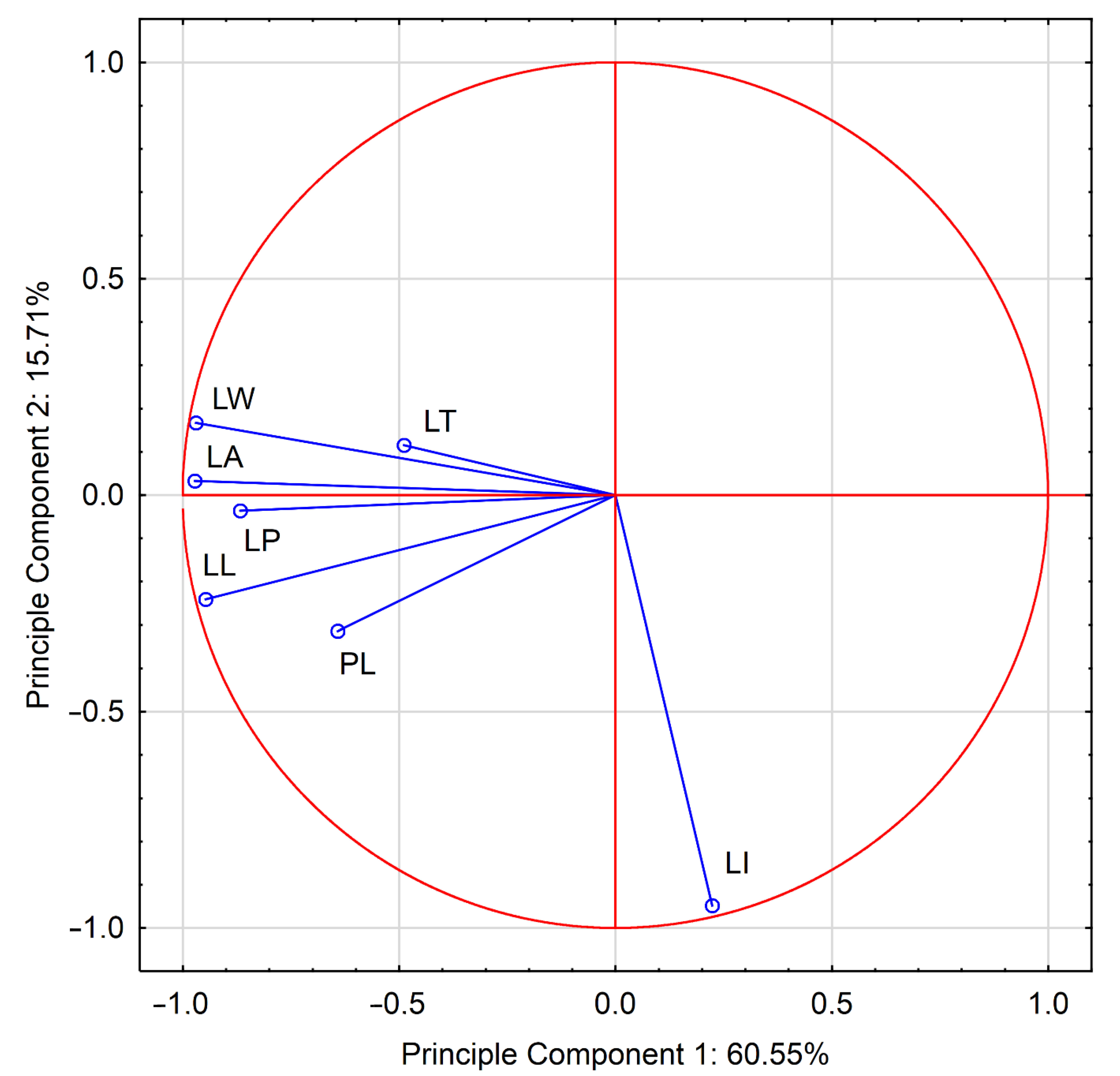
3.2. Morphological Features of Leaves
3.3. Biochemical Analyses
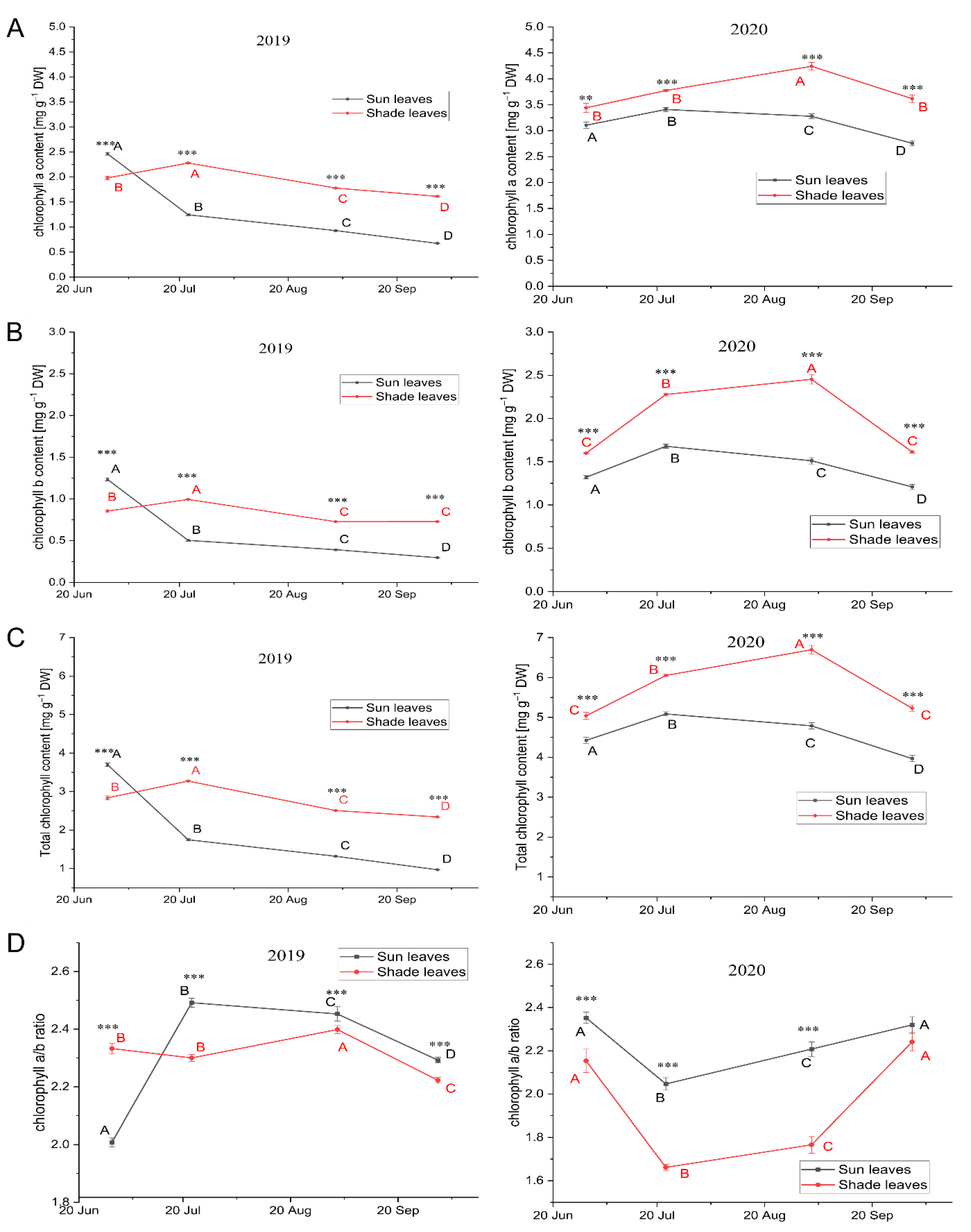
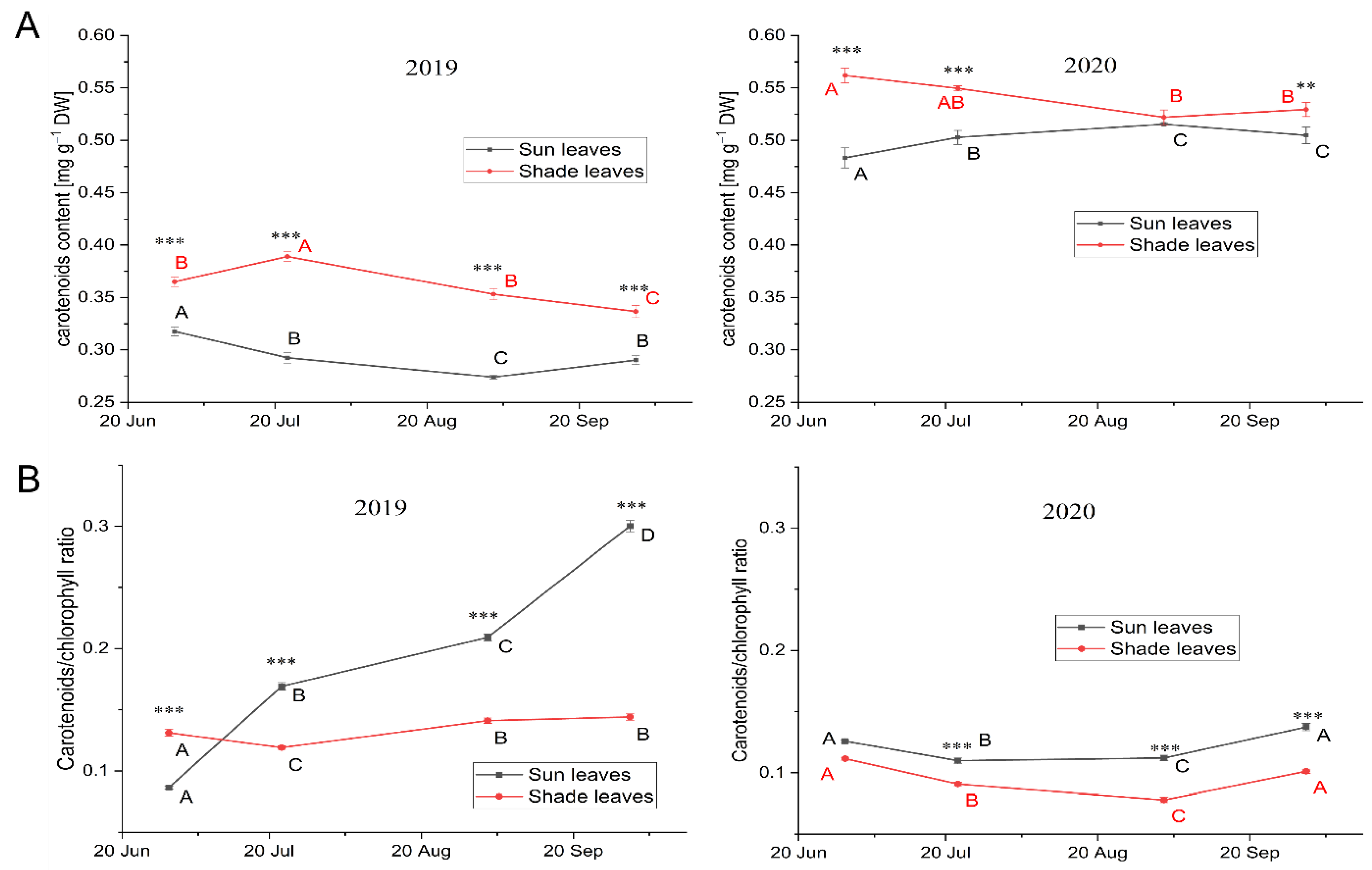
3.3.1. Leaf Pigment Content
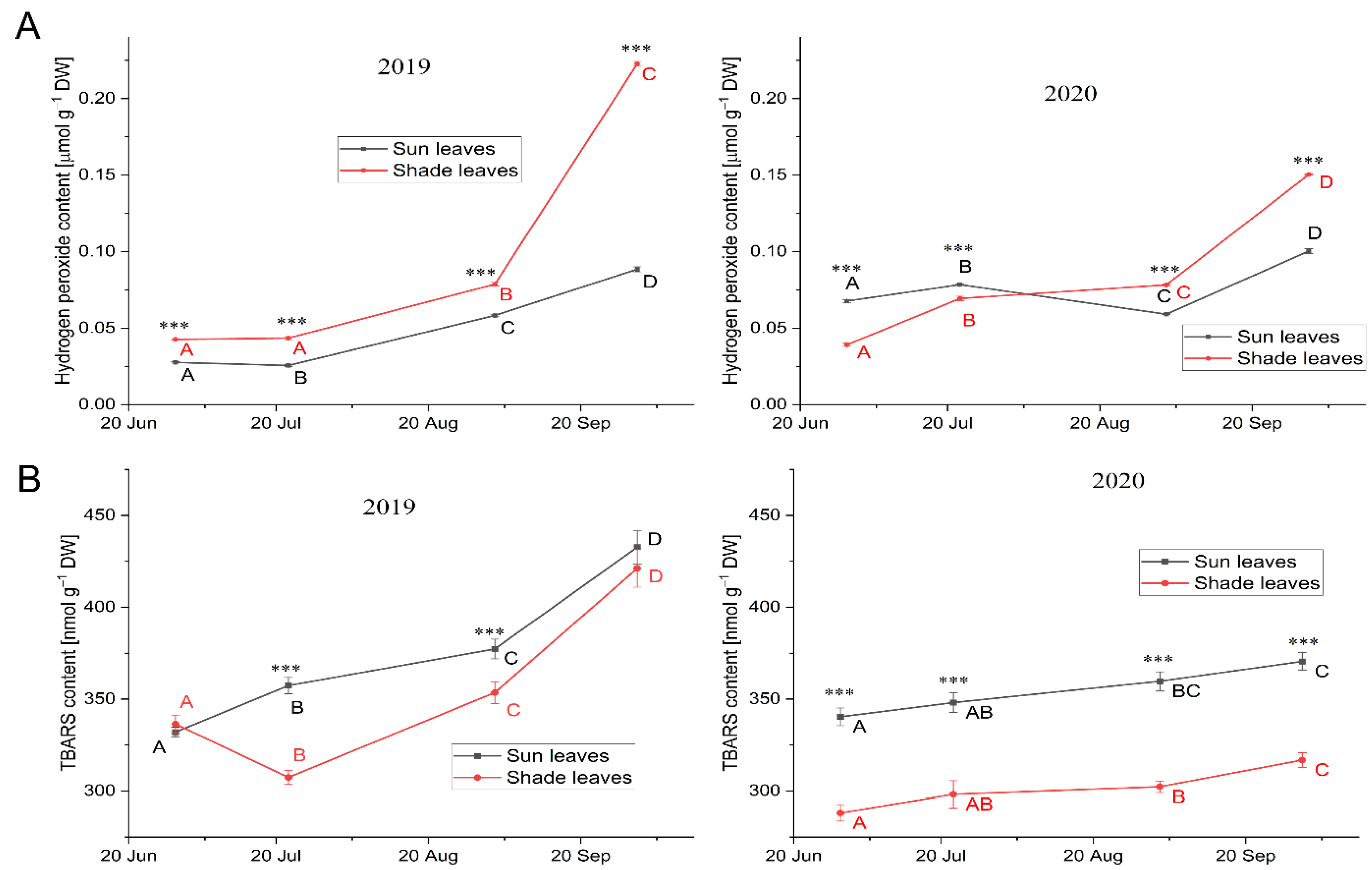
3.3.2. Oxidative Stress Markers
3.3.3. Low-Molecular-Weight Antioxidants
3.3.4. Antioxidative Enzymes
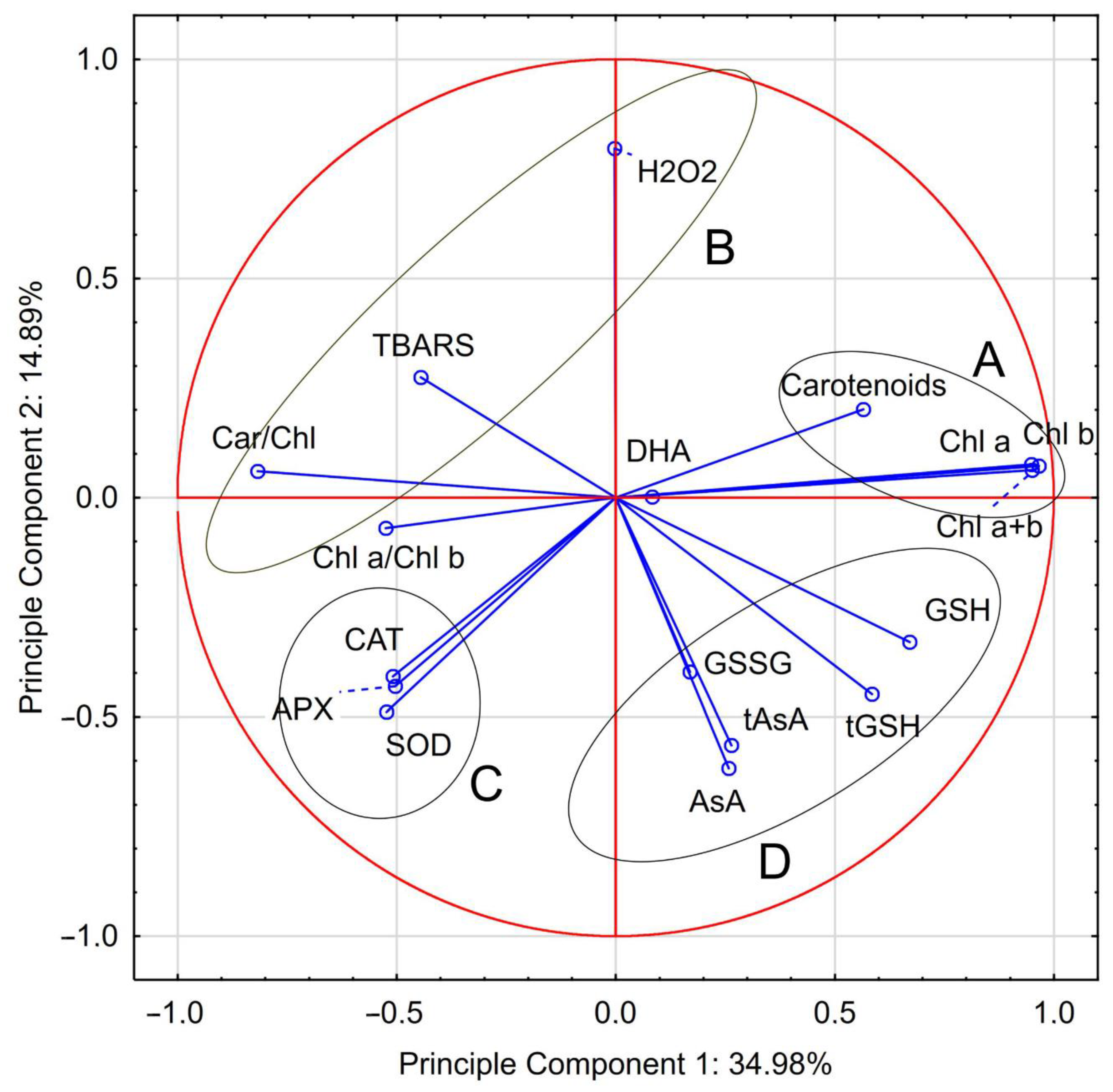
3.3.5. Correlation and Multivariate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lambers, H.; Chapin, F.S.; Pons, T.L. (Eds.) Growth and allocation in plant physiological ecology. In Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; pp. 321–374. [Google Scholar]
- Pons, T.L. Regulation of leaf traits in canopy gradients. In Canopy Photosynthesis: From Basics to Applications; Hikosaka, K., Niinemets, Ü., Anten, N.P.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 143–168. [Google Scholar]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Zentgraf, U. The correlation between oxidative stress and leaf senescence during plant development. Cell. Mol. Biol. Lett. 2005, 10, 515–534. [Google Scholar] [PubMed]
- Curien, G.; Flori, S.; Villanova, V.; Magneschi, L.; Giustini, C.; Forti, G.; Matringe, M.; Petroutsos, D.; Kuntz, M.; Finazzi, G. The Water to Water Cycles in Microalgae. Plant Cell Physiol. 2016, 57, 1354–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polle, A. Mehler reaction: Friend or foe in photosynthesis? Bot. Acta 1996, 109, 84–89. [Google Scholar] [CrossRef]
- Koppenol, W.H. The Haber-Weiss cycle—70 years later. Redox Rep. 2001, 6, 229–234. [Google Scholar] [CrossRef]
- Terashima, I.; Miyazawa, S.I.; Hanba, Y.T. Why are sun leaves thicker than shade leaves? Consideration based on analyses of CO2 diffusion in the leaf. J. Plant Res. 2001, 114, 93–105. [Google Scholar] [CrossRef]
- Kraj, W.; Sztorc, A. Genetic structure and variability of phenological forms in the European beech (Fagus sylvatica L.). Ann. For. Sci. 2009, 66, 203. [Google Scholar] [CrossRef] [Green Version]
- Kraj, W.; Zarek, M. Biochemical basis of altitude adaptation and antioxidant system activity during autumn leaf senescence in beech populations. Forests 2021, 12, 529. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Kramer, P.J.; Pallardy, S.G. (Eds.) Physiological and environmental requirements for tree growth. In The Physiological Ecology of Woody Plants; Academic Press: San Diego, CA, USA, 1991; pp. 31–68. [Google Scholar]
- Desotgiu, R.; Cascio, C.; Pollastrini, M.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Short and long term photosynthetic adjustments in sun and shade leaves of Fagus sylvatica L., investigated by fluorescence transient (FT) analysis. Plant Biosyst. 2012, 146, 206–216. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Gil Nam, H. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Buchanan-Wollaston, V.; Earl, S.; Harrison, E.; Mathas, E.; Navabpour, S.; Page, T.; Pink, D. The molecular analysis of leaf senescence—A genomics approach. Plant Biotechnol. J. 2003, 1, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Fracheboud, Y.; Luquez, V.; Björkén, L.; Sjödin, A.; Tuominen, H.; Jansson, S. The control of autumn senescence in European aspen. Plant Physiol. 2009, 149, 1982–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraj, W. Antioxidative enzyme activity as the factor causing differential autumn senescence in phenological forms of beech (Fagus sylvatica L.). Acta Physiol. Plant. 2017, 39, 16. [Google Scholar] [CrossRef]
- Kraj, W. Reactive oxygen species and antioxidant levels as the factors of autumn senescence in phenological forms of beech (Fagus sylvatica L.). Acta Physiol. Plant. 2016, 38, 32. [Google Scholar] [CrossRef]
- Kraj, W. Chlorophyll degradation and the activity of chlorophyllase and Mg-dechelatase during leaf senescence in Fagus sylvatica. Dendrobiology 2015, 74, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Hatziskakis, S.; Tsiripidis, I.; Papageorgiou, A.C. Leaf morphological variation in beech (Fagus sylvatica L.) populations in Greece and its relation to their post-glacial origin. Bot. J. Linn. Soc. 2011, 165, 422–436. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Gillespie, K.M.; Ainsworth, E.A. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc. 2007, 2, 871. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Isolation, purification, characterization, and assay of antioxygenic enzymes: Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate–Glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. A microtiter plate assay for superoxide dismutase using a water-soluble tetrazolium salt (WST-1). Clin. Chim. Acta 2000, 293, 157–166. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; A Series of books in biology; W. H. Freeman and Company: New York, NY, USA, 1995; p. 887. [Google Scholar]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Pallardy, S.G. (Ed.) Vegetative Growth. In Physiology of Woody Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2008; pp. 39–86. [Google Scholar]
- Coble, A.P.; Fogel, M.L.; Parker, G.G. Canopy gradients in leaf functional traits for species that differ in growth strategies and shade tolerance. Tree Physiol. 2017, 37, 1415–1425. [Google Scholar] [CrossRef]
- Naidu, S.L.; DeLucia, E.H. Physiological and morphological acclimation of shade-grown tree seedlings to late-season canopy gap formation. Plant Ecol. 1998, 138, 27–40. [Google Scholar] [CrossRef]
- Eschrich, W.; Burchardt, R.; Essiamah, S. The induction of sun and shade leaves of the European beech (Fagus sylvatica L.): Anatomical studies. Trees 1989, 3, 1–10. [Google Scholar] [CrossRef]
- Uemura, A.; Ishida, A.; Nakano, T.; Terashima, I.; Tanabe, H.; Matsumoto, Y. Acclimation of leaf characteristics of Fagus species to previous-year and current-year solar irradiances. Tree Physiol. 2000, 20, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Dolnicki, A.; Kraj, W. Leaf morphology and the dynamics of frost-hardiness of shoots in two phenological forms of European beech (Fagus sylvatica L.) from Southern Poland. EJPAU 2001, 4. Available online: http://www.ejpau.media.pl/volume4/issue2/forestry/art-01.html (accessed on 16 July 2022).
- Stojnić, S.; Orlović, S.; Miljković, D.; Von Wuehlisch, G. Intra- and interprovenance variations in leaf morphometric traits in European beech (Fagus sylvatica L.). Arch. Biol. Sci. 2016, 68, 781–788. [Google Scholar] [CrossRef]
- Kempf, M.; Banach, J.; Skrzyszewska, K. Morphological variability of beech leaves from early and late flushing provenances. Balt. For. 2018, 24, 210–217. [Google Scholar]
- Ishida, A.; Toma, T. Marjenah Limitation of leaf carbon gain by stomatal and photochemical processes in the top canopy of Macaranga conifera, a tropical pioneer tree. Tree Physiol. 1999, 19, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Oguchi, R.; Hikosaka, K.; Hirose, T. Leaf anatomy as a constraint for photosynthetic acclimation: Differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees. Plant Cell Environ. 2005, 28, 916–927. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. (Eds.) Photosynthesis, Respiration and Long-Distance Transport. In Plant Physiological Ecology; Springer: New York, NY, USA, 2008; pp. 11–99. [Google Scholar]
- Petkova, K.; Molle, E.; Huber, G.; Konnert, M.; Gaviria, J. Spring and autumn phenology of Bulgarian and German provenances of Common beech (Fagus sylvatica L.) under similar climatic conditions. Silvae Genet. 2017, 66, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Chmura, D.J.; Rożkowski, R. Variability of beech provenances in spring and autumn phenology. Silvae Genet. 2002, 51, 123–127. [Google Scholar]
- Schueler, S.; Liesebach, M. Latitudinal population transfer reduces temperature sum requirements for bud burst of European beech. Plant Ecol. 2014, 216, 111–122. [Google Scholar] [CrossRef]
- Kraj, W. Proteolytic activity and nitrogen remobilisation in senescing leaves of phenological forms of Fagus sylvatica. Dendrobiology 2014, 72, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Denk, T. The taxonomy of Fagus in western Eurasia. 2: Fagus sylvatica subsp. sylvatica. Feddes Repert. 1999, 110, 381–412. [Google Scholar] [CrossRef]
- Hollinger, D.Y. Optimality and nitrogen allocation in a tree canopy. Tree Physiol. 1996, 16, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Babani, F. Light adaptation and senescence of the photosynthetic apparatus. Changes in pigment composition, chlorophyll fluorescence parameters and photosynthetic activity. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, G., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 713–736. [Google Scholar]
- Vitasse, Y.; Baumgarten, F.; Zohner, C.M.; Kaewthongrach, R.; Fu, Y.H.; Walde, M.G.; Moser, B. Impact of microclimatic conditions and resource availability on spring and autumn phenology of temperate tree seedlings. New Phytol. 2021, 232, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.A. Reactive Oxygen Species and Photosynthesis. In Antioxidants and Reactive Oxygen Species in Plants; Smirnoff, N., Ed.; Oxford Blackwell Publishing: Oxford, UK, 2005; pp. 250–267. [Google Scholar]
- Racchi, M.L. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef] [PubMed]
- Tukaj, Z.; Pokora, W. Individual and combined effect of anthracene, cadmium, and chloridazone on growth and activity of SOD izoformes in three Scenedesmus species. Ecotox. Environ. Safe. 2006, 65, 323–331. [Google Scholar] [CrossRef]
- Mach, J.M.; Greenberg, J.T. Free Radicals and Oxidative Stress. In Plant Cell Death Processes; Noodén, L.D., Ed.; Academic Press: San Diego, CA, USA, 2004; pp. 203–214. [Google Scholar]
- Wingler, A.; Marès, M.; Pourtau, N. Spatial patterns and metabolic regulation of photosynthetic parameters during leaf senescence. New Phytol. 2004, 161, 781–789. [Google Scholar] [CrossRef]
- Hörtensteiner, S.; Kräutler, B. Chlorophyll breakdown in higher plants. BBA-Bioenerg. 2011, 1807, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Kraj, W. Stem girdling affects the carbon/nitrogen imbalance and oxidative stress, and induces leaf senescence in phenological forms of beech (Fagus sylvatica). Acta Biol. Cracov. Bot. 2017, 59, 67–79. [Google Scholar] [CrossRef]
- Delpierre, N.; Dufrêne, E.; Soudani, K.; Ulrich, E.; Cecchini, S.; Boé, J.; François, C. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France. Agric. For. Meteorol. 2009, 149, 938–948. [Google Scholar] [CrossRef]
- Liu, Q.; Piao, S.; Campioli, M.; Gao, M.; Fu, Y.H.; Wang, K.; He, Y.; Li, X.; Janssens, I.A. Modeling leaf senescence of deciduous tree species in Europe. Glob. Chang. Biol. 2020, 26, 4104–4118. [Google Scholar] [CrossRef]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Jubany-Marí, T.; Alegre, L. Drought-induced senescence is characterized by a loss of antioxidant defences in chloroplasts. Plant Cell Environ. 2001, 24, 1319–1327. [Google Scholar] [CrossRef]
- García-Plazaola, J.I.; Becerril, J.M. Seasonal changes in photosynthetic pigments and antioxidants in beech (Fagus sylvatica) in a Mediterranean climate: Implications for tree decline diagnosis. Aust. J. Plant Physiol. 2001, 28, 225–232. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Year of Leaf Collection | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | |||||||||
| Temperature [°C] | Total Prec. | Temperature [°C] | Total Prec. | |||||||
| Min. | Ave. | Max | Amp. | [mm] | Min. | Ave. | Max | Amp. | [mm] | |
| June (last 2 weeks) | 16.6 | 22.2 | 28.06 | 11.49 | 4.0 | 15.7 | 19.2 | 23.8 | 8.15 | 52.1 |
| July | 13.5 | 19.2 | 25.4 | 11.87 | 74.7 | 13.1 | 19.0 | 25.3 | 12.19 | 65.5 |
| August | 15.0 | 20.3 | 26.7 | 11.64 | 79.3 | 14.7 | 20.3 | 26.7 | 11.98 | 108.6 |
| September | 9.9 | 14.5 | 20.2 | 10.27 | 78.3 | 9.7 | 15.1 | 21.2 | 11.47 | 79.2 |
| Whole sampling period | ||||||||||
| Average | 13.3 | 18.6 | 24.6 | 11.37 | 236.3 | 13.0 | 18.3 | 24.4 | 11.41 | 305.4 |
| SD | 3.8 | 3.9 | 4.7 | 3.45 | 3.23 | 4.16 | ||||
| V% | 28.3 | 20.7 | 18.9 | 26.61 | 17.62 | 17.05 | ||||
| Leaf Feature | Leaf Type | Mann–Whitney U Test Z Value | p | |||||
|---|---|---|---|---|---|---|---|---|
| Light | Shade | |||||||
| Ave. | SE | V% | Ave. | SE | V% | |||
| Leaf length [cm] | 5.74 | 0.13 | 17.61 | 4.70 | 0.1 | 16.37 | 5.708 | <0.001 |
| Leaf width [cm] | 3.21 | 0.08 | 20.28 | 2.54 | 0.05 | 15.29 | 6.186 | <0.001 |
| Leaf area [cm2] | 12.1 | 0.55 | 35.63 | 7.94 | 0.31 | 30.35 | 5.934 | <0.001 |
| Leaf thickness [μm] | 110.83 | 2.07 | 14.44 | 82.3 | 1.36 | 12.80 | 8.212 | <0.001 |
| Leaf perimeter [cm] | 16.17 | 0.36 | 19.70 | 11.91 | 0.40 | 26.07 | 4.876 | <0.001 |
| Petiole length [cm] | 0.69 | 0.03 | 28.07 | 0.55 | 0.02 | 26.18 | 4.905 | <0.001 |
| Leaf index | 180.4 | 2.07 | 8.88 | 185.8 | 1.94 | 8.07 | −1.918 | 0.055 |
| Petiole index | 12.16 | 0.34 | 21.58 | 16.68 | 0.75 | 49.27 | −3.825 | <0.001 |
| A | ||||||||||||
| Factor | H2O2 | TBARS | Car/chl | |||||||||
| Light | Shade | Light | Shade | Light | Shade | |||||||
| r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | |
| Chl a | −0.694 *** | −0.03 | −0.669 *** | −0.17 | −0.340 *** | −32.03 | −0.406 *** | −80.31 | −0.887 *** | −0.10 | −0.638 *** | −0.04 |
| Chl b | −0.623 *** | −0.04 | −0.532 *** | −0.30 | −0.287 *** | −49.98 | −0.328 *** | −144.14 | −0.847 *** | −0.18 | −0.649 *** | −0.10 |
| Chl a + b | −0.671 *** | −0.02 | −0.636 *** | −0.11 | −0.322 *** | −19.76 | −0.387 *** | −53.58 | −0.875 *** | −0.07 | −0.651 *** | −0.03 |
| B | ||||||||||||
| Factor | tAsA | AsA | DHA | |||||||||
| Light | Shade | Light | Shade | Light | Shade | |||||||
| r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | |
| H2O2 | 0.220 *** | 50.73 | −0.372 *** | −37.29 | 0.147 ** | 20.60 | −0.626 *** | 63.99 | 0.189 ** | 30.10 | 0.491 *** | 25.59 |
| TBARS | 0.064 | 0.010 | −0.150 *** | −0.019 | −0.011 | 0.000 | −0.376 *** | −0.049 | 0.125 | 0.010 | 0.442 *** | 0.029 |
| Car/chl | 0.328 *** | 25.05 | −0.342 * | −126.63 | 0.347 *** | 15.75 | −0.394 *** | −148.81 | 0.168 ** | 8.85 | 0.147 * | 28.21 |
| C | ||||||||||||
| Factor | tGSH | GSH | GSSG | |||||||||
| Light | Shade | Light | Shade | Light | Shade | |||||||
| r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | |
| H2O2 | −0.526 *** | −1818.44 | −0.587 *** | −475.06 | −0.553 *** | −880.31 | −0.252 *** | −149.10 | −0.475 *** | −940.19 | −0.801 *** | −325.95 |
| TBARS | −0.443 *** | −0.590 | −0.238 *** | −0.245 | −0.452 *** | −0.290 | −0.048 | −0.036 | −0.408 *** | −0.330 | −0.404 *** | −0.209 |
| Car/chl | −0.475 *** | −543.15 | −0.049 | −149.17 | −0.530 *** | −279.37 | 0.177 ** | 386.12 | −0.410 *** | −268.80 | −0.356 *** | −535.29 |
| D | ||||||||||||
| Factor | CAT | APX | SOD | |||||||||
| Light | Shade | Light | Shade | Light | Shade | |||||||
| r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | r | Slope | |
| H2O2 | 0.165 ** | 123.32 | −0.441 *** | −255.40 | −0.508 *** | −1.880 | −0.585 *** | −1.196 | 0.273 *** | 330.45 | −0.560 *** | −214.94 |
| TBARS | 0.000 | 0.000 | −0.190** | −0.140 | −0.237 *** | 0.000 | −0.501 *** | −0.001 | −0.096 | 0.050 | −0.426 *** | −0.208 |
| Car/chl | 0.319 *** | 78.84 | −0.317 *** | −678.47 | −0.553 *** | −0.680 | −0.223 *** | −1.689 | 0.278 *** | 111.44 | −0.171 ** | −242.109 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraj, W.; Ślepaczuk, A. Morphophysiological Acclimation of Developed and Senescing Beech Leaves to Different Light Conditions. Forests 2022, 13, 1333. https://doi.org/10.3390/f13081333
Kraj W, Ślepaczuk A. Morphophysiological Acclimation of Developed and Senescing Beech Leaves to Different Light Conditions. Forests. 2022; 13(8):1333. https://doi.org/10.3390/f13081333
Chicago/Turabian StyleKraj, Wojciech, and Artur Ślepaczuk. 2022. "Morphophysiological Acclimation of Developed and Senescing Beech Leaves to Different Light Conditions" Forests 13, no. 8: 1333. https://doi.org/10.3390/f13081333