
Changes in Forest Conditions in a Siberian Larch Forest Induced by an Extreme Wet Event

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
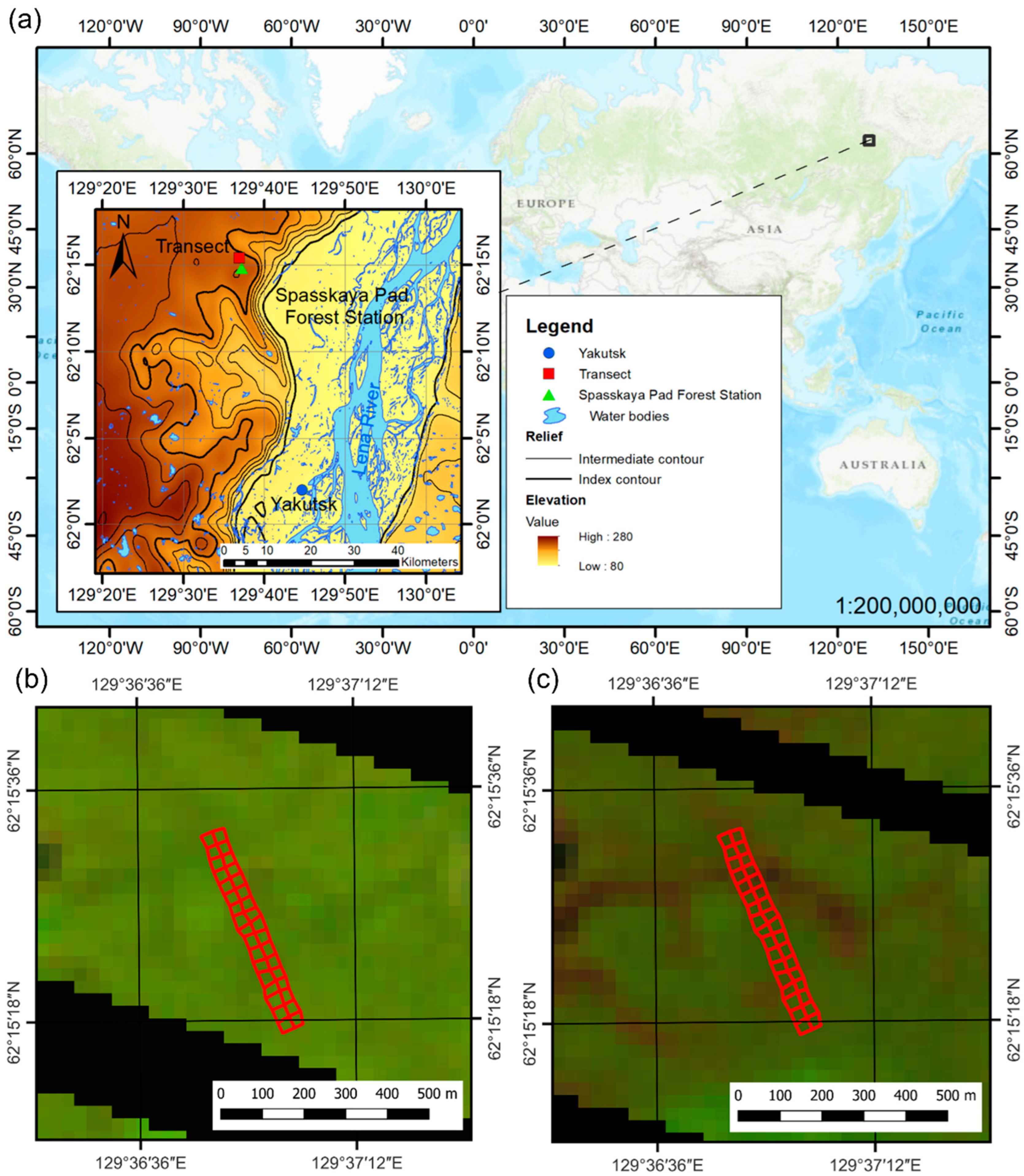
2.1. Study Site and Setting of the Observation Plots
2.2. NDVI
2.3. Larch Needle Sampling and Carbon and Nitrogen Analyses
2.4. Statistical Analysis
3. Results
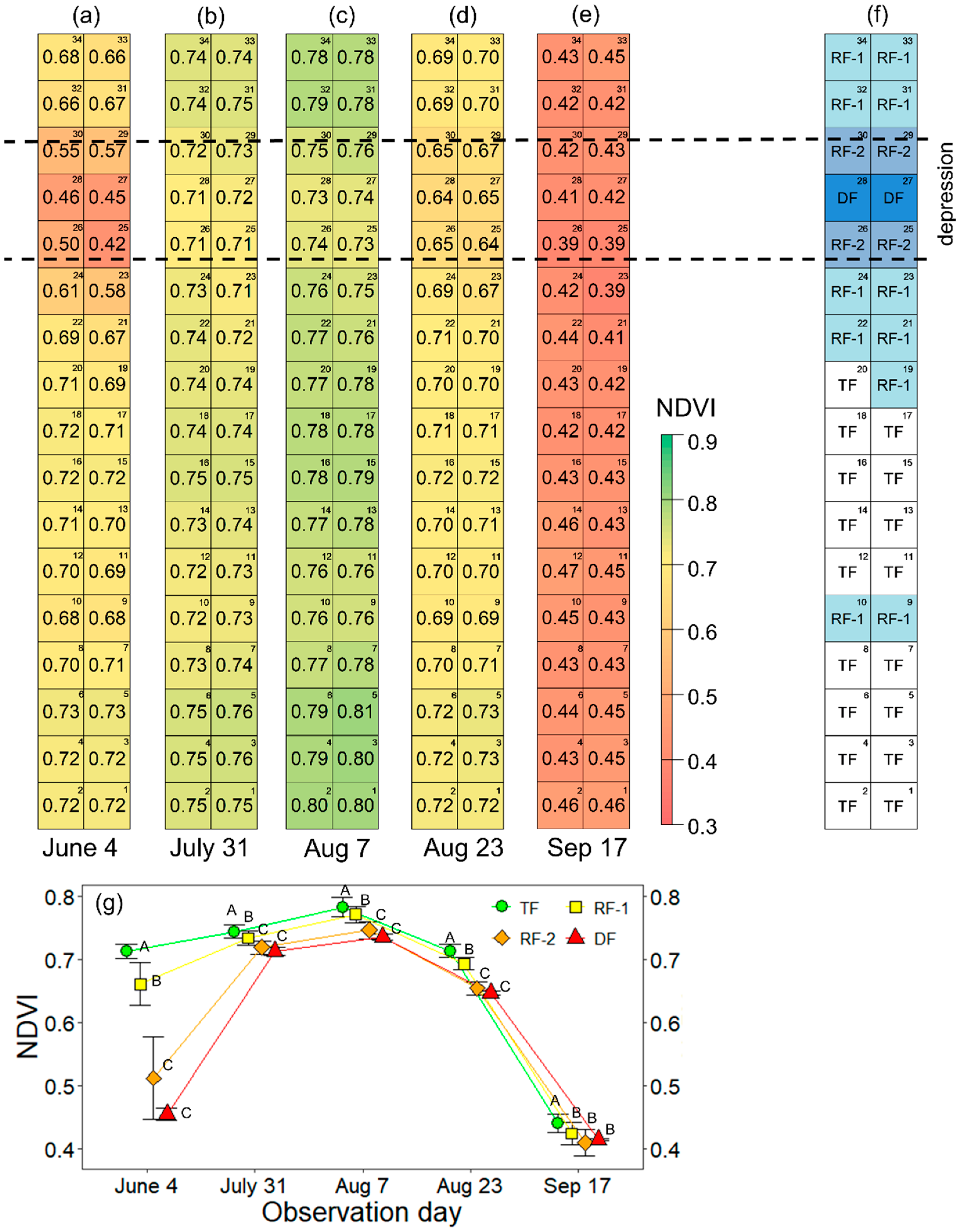
3.1. Seasonal Variation of the NDVI and Forest Condition
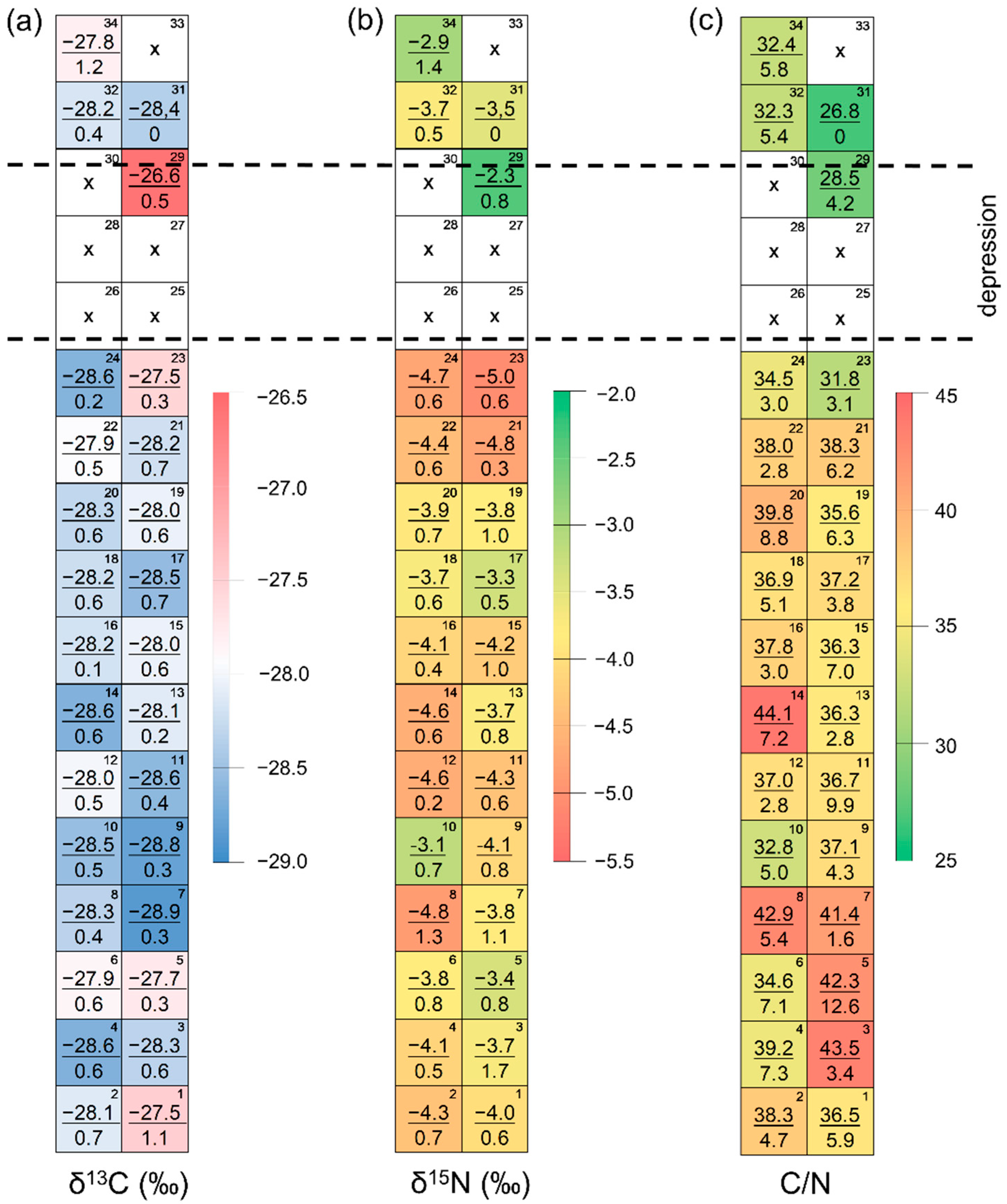
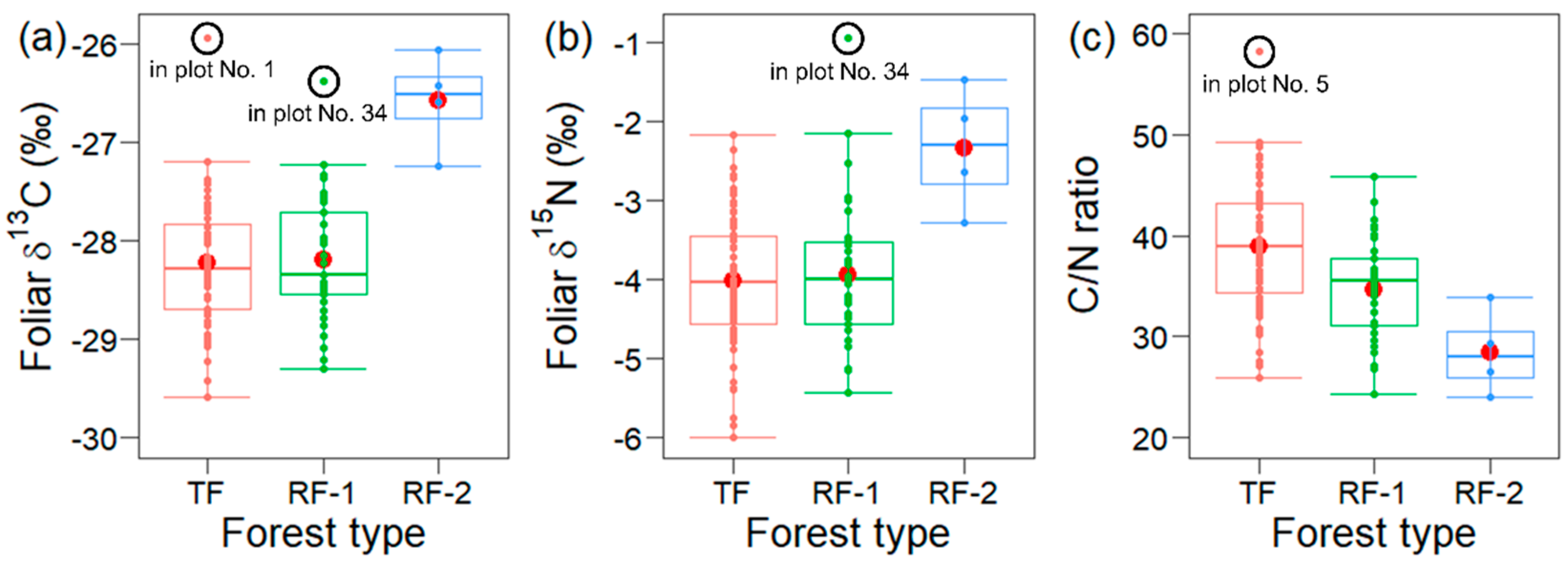
3.2. Spatial Variations in the Larch Foliar Traits
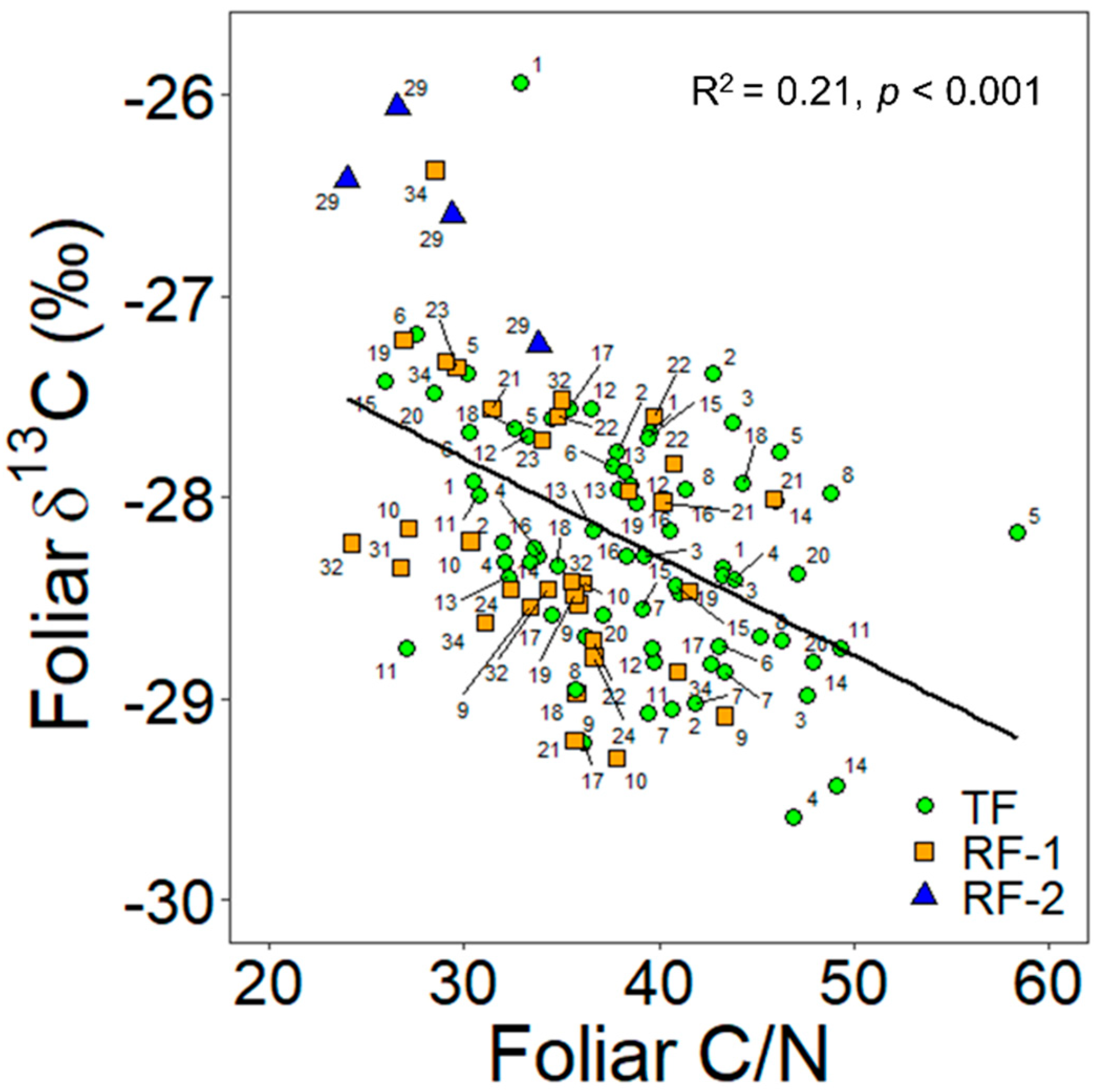
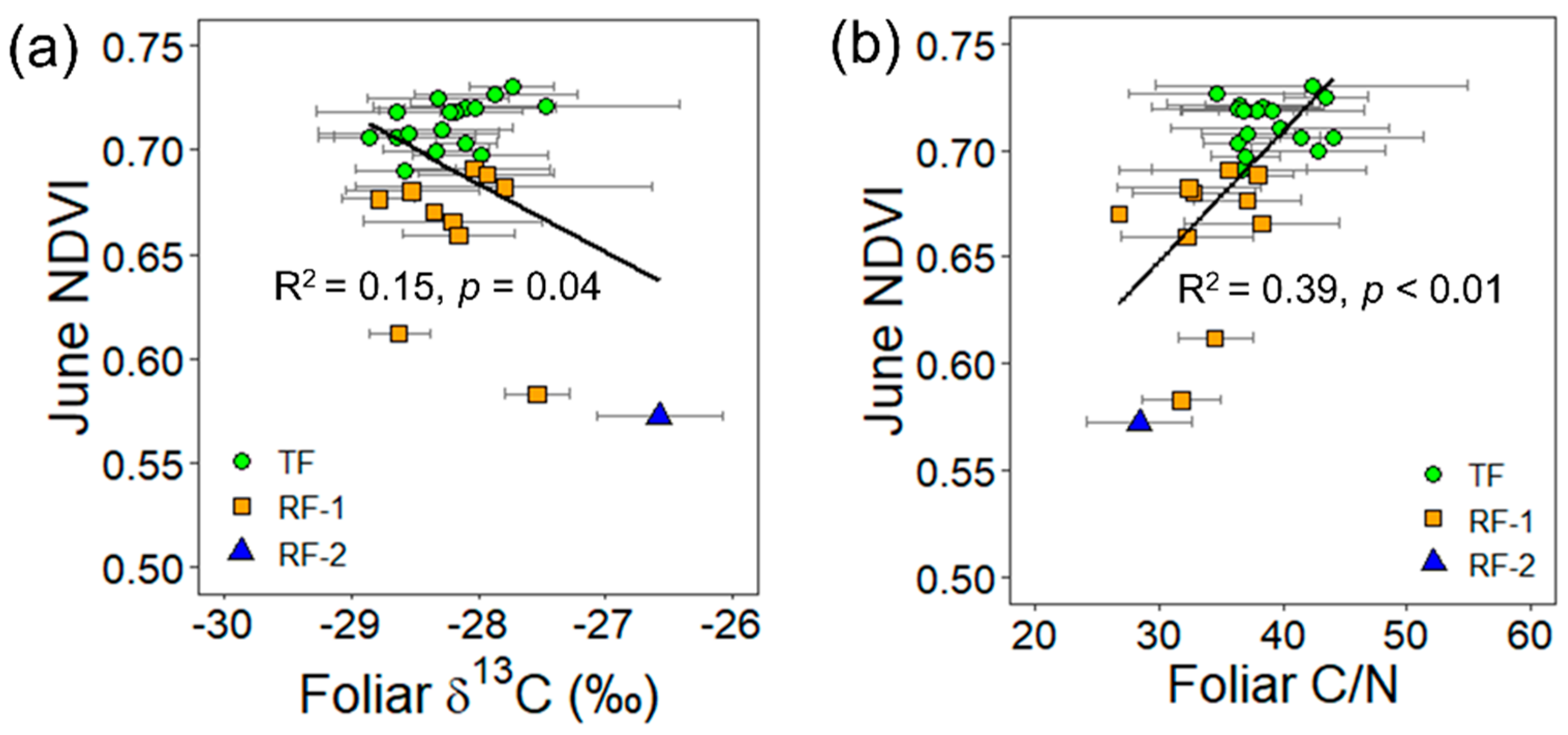
3.3. Relationships between the NDVI and the Foliar Traits
4. Discussion
4.1. Plant Phenology and Seasonality of the Ecosystem of Each Forest Type
4.2. Forest Condition after the Damage by the Wet Event
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Douville, H.; Raghavan, K.; Renwick, J.; Allan, R.P.; Arias, P.A.; Barlow, M.; Cerezo-Mota, R.; Cherchi, A.; Gan, T.Y.; Gergis, J.; et al. Water Cycle Changes. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 1055–1210. [Google Scholar]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of More Extreme Precipitation Regimes for Terrestrial Ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Jiang, P.; Liu, H.Y.; Piao, S.L.; Ciais, P.; Wu, X.C.; Yin, Y.; Wang, H.Y. Enhanced growth after extreme wetness compensates for post-drought carbon loss in dry forests. Nat. Commun. 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Heisler-White, J.L.; Blair, J.M.; Kelly, E.F.; Harmoney, K.; Knapp, A.K. Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Glob. Chang. Biol. 2009, 15, 2894–2904. [Google Scholar] [CrossRef]
- Rozas, V.; Garcia-Gonzalez, I. Too wet for oaks? Inter-tree competition and recent persistent wetness predispose oaks to rainfall-induced dieback in Atlantic rainy forest. Glob. Planet. Chang. 2012, 94–95, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, H.; Saito, H.; Kuwao, K.; Maximov, T.C.; Hasegawa, S. Forest decline caused by high soil water conditions in a permafrost region. Hydrol. Earth Syst. Sci. 2010, 14, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Abaimov, A.P.; Lesinski, J.A.; Martinsson, O.; Milyutin, L.I. Variability and Ecology of Siberian Larch Species; Reports; Swedish University of Agricultural Sciences, Departament of Silviculture: Umeå, Sweden, 1998; Volume 43, p. 123. [Google Scholar]
- Archibold, O.W. The coniferous forests. In Ecology of World Vegetation; Springer: Dordrecht, The Netherlands, 1995; pp. 238–279. [Google Scholar] [CrossRef]
- Sugimoto, A.; Yanagisawa, N.; Naito, D.; Fujita, N.; Maximov, T.C. Importance of permafrost as a source of water for plants in east Siberian taiga. Ecol. Res. 2002, 17, 493–503. [Google Scholar] [CrossRef]
- Sugimoto, A.; Naito, D.; Yanagisawa, N.; Ichiyanagi, K.; Kurita, N.; Kubota, J.; Kotake, T.; Ohata, T.; Maximov, T.C.; Fedorov, A.N. Characteristics of soil moisture in permafrost observed in East Siberian taiga with stable isotopes of water. Hydrol. Process. 2003, 17, 1073–1092. [Google Scholar] [CrossRef]
- Popova, A.S.; Tokuchi, N.; Ohte, N.; Ueda, M.U.; Osaka, K.; Maximov, T.C.; Sugimoto, A. Nitrogen availability in the taiga forest ecosystem of northeastern Siberia. Soil Sci. Plant Nutr. 2013, 59, 427–441. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, Y.; Hirobe, M. Soil carbon and nitrogen, and characteristics of soil active layer in Siberian permafrost region. In Permafrost Ecosystems. Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 149–163. [Google Scholar] [CrossRef]
- Sugimoto, A. Stable Isotopes of Water in Permafrost Ecosystem. In Water-Carbon Dynamics in Eastern Siberia. Ecological Studies (Analysis and Synthesis); Ohta, T., Hiyama, T., Iijima, Y., Kotani, A., Maximov, T., Eds.; Springer: Singapore, 2019; Volume 236. [Google Scholar]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Yamazaki, T.; Maximov, T.C. Reconstruction of soil moisture for the past 100 years in eastern Siberia by using delta C-13 of larch tree rings. J. Geophys. Res.-Biogeosci. 2013, 118, 1256–1265. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using landsat TM images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Wang, Q.; Adiku, S.; Tenhunen, J.; Granier, A. On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens. Environ. 2005, 94, 244–255. [Google Scholar] [CrossRef]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Penuelas, J.; Valentini, R. Relationships between NDVI, canopy structure, and photosynthesis in 3 Californian vegetation types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Juszak, I.; Erb, A.M.; Maximov, T.C.; Schaepman-Strub, G. Arctic shrub effects on NDVI, summer albedo and soil shading. Remote Sens. Environ. 2014, 153, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Blok, D.; Schaepman-Strub, G.; Bartholomeus, H.; Heijmans, M.; Maximov, T.C.; Berendse, F. The response of Arctic vegetation to the summer climate: Relation between shrub cover, NDVI, surface albedo and temperature. Environ. Res. Lett. 2011, 6, 035502. [Google Scholar] [CrossRef] [Green Version]
- Myneni, R.B.; Williams, D.L. On the relationship between FAPAR and NDVI. Remote Sens. Environ. 1994, 49, 200–211. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Clevers, J.; Gitelson, A.A. Remote estimation of crop and grass chlorophyll and nitrogen content using red-edge bands on Sentinel-2 and-3. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 344–351. [Google Scholar] [CrossRef]
- Wang, Z.H.; Wang, T.J.; Darvishzadeh, R.; Skidmore, A.K.; Jones, S.; Suarez, L.; Woodgate, W.; Heiden, U.; Heurich, M.; Hearne, J. Vegetation Indices for Mapping Canopy Foliar Nitrogen in a Mixed Temperate Forest. Remote Sens. 2016, 8, 491. [Google Scholar] [CrossRef] [Green Version]
- Dyer, M.I.; Turner, C.L.; Seastedt, T.R. Mowing and fertilization effects on productivity and spectral reflectance in Bromus-inermis plots. Ecol. Appl. 1991, 1, 443–452. [Google Scholar] [CrossRef]
- Santos, G.O.; Rosalen, D.L.; De Faria, R.T. Use of active optical sensor in the characteristics analysis of the fertigated Brachiaria with treated sewage. Eng. Agric. 2017, 37, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.L.; Seastedt, T.R.; Dyer, M.I.; Kittel, T.G.F.; Schimel, D.S. Effects of management and topography on the radiometric response of a tallgrass prairie. J. Geophys. Res.-Atmos. 1992, 97, 18855–18866. [Google Scholar] [CrossRef]
- Zhu, Y.; Tian, Y.C.; Yao, X.; Liu, X.J.; Cao, W.X. Analysis of common canopy reflectance spectra for indicating leaf nitrogen concentrations in wheat and rice. Plant Prod. Sci. 2007, 10, 400–411. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yang, C.M.; Chang, K.W.; Shen, Y. A simple spectral index using reflectance of 735 nm to assess nitrogen status of rice canopy. Agron. J. 2008, 100, 205–212. [Google Scholar] [CrossRef]
- Liang, M.C.; Sugimoto, A.; Tei, S.; Bragin, I.V.; Takano, S.; Morozumi, T.; Shingubara, R.; Maximov, T.C.; Kiyashko, S.I.; Velivetskaya, T.A.; et al. Importance of soil moisture and N availability to larch growth and distribution in the Arctic taiga-tundra boundary ecosystem, northeastern Siberia. Polar Sci. 2014, 8, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, M.; Choi, W.J.; Chang, S.X. White spruce foliar delta C-13 and delta N-15 indicate changed soil N availability by understory removal and N fertilization in a 13-year-old boreal plantation. Plant Soil 2012, 361, 375–384. [Google Scholar] [CrossRef]
- Liu, J.X.; Price, D.T.; Chen, J.A. Nitrogen controls on ecosystem carbon sequestration: A model implementation and application to Saskatchewan, Canada. Ecol. Model. 2005, 186, 178–195. [Google Scholar] [CrossRef]
- Li, S.G.; Tsujimura, M.; Sugimoto, A.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Temporal variation of delta C-13 of larch leaves from a montane boreal forest in Mongolia. Trees-Struct. Funct. 2007, 21, 479–490. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Yousfi, S.; Kellas, N.; Saidi, L.; Benlakehal, Z.; Chaou, L.; Siad, D.; Herda, F.; Karrou, M.; Vergara, O.; Gracia, A.; et al. Comparative performance of remote sensing methods in assessing wheat performance under Mediterranean conditions. Agric. Water Manag. 2016, 164, 137–147. [Google Scholar] [CrossRef]
- Stamatiadis, S.; Taskos, D.; Tsadila, E.; Christofides, C.; Tsadilas, C.; Schepers, J.S. Comparison of passive and active canopy sensors for the estimation of vine biomass production. Precis. Agric. 2010, 11, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.M.; Xie, G.D. The relationship between plant stable carbon isotope composition, precipitation and satellite data, Tibet Plateau, China. Quat. Int. 2006, 144, 68–71. [Google Scholar] [CrossRef]
- del Castillo, J.; Voltas, J.; Ferrio, J.P. Carbon isotope discrimination, radial growth, and NDVI share spatiotemporal responses to precipitation in Aleppo pine. Trees-Struct. Funct. 2015, 29, 223–233. [Google Scholar] [CrossRef]
- Ale, R.; Zhang, L.; Li, X.; Raskoti, B.B.; Pugnaire, F.I.; Luo, T.X. Water Shortage Drives Interactions Between Cushion and Beneficiary Species Along Elevation Gradients in Dry Himalayas. J. Geophys. Res.-Biogeosci. 2018, 123, 226–238. [Google Scholar] [CrossRef]
- Duursma, R.A.; Marshall, J.D. Vertical canopy gradients in delta C-13 correspond with leaf nitrogen content in a mixed-species conifer forest. Trees-Struct. Funct. 2006, 20, 496–506. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Field, C.B.; Lin, Z.F.; Kuo, C.Y. Leaf carbon isotope and mineral-composition in subtropical plants along an irradiance cline. Oecologia 1986, 70, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Garten, C.T.; Taylor, G.E. Foliar delta C-13 within a temperature deciduous forest-spatial, temporal, and species sources of variation. Oecologia 1992, 90, 1–7. [Google Scholar] [CrossRef]
- Michelsen, A.; Schmidt, I.K.; Jonasson, S.; Quarmby, C.; Sleep, D. Leaf N-15 abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia 1996, 105, 53–63. [Google Scholar] [CrossRef]
- Handley, L.L.; Austin, A.T.; Robinson, D.; Scrimgeour, C.M.; Raven, J.A.; Heaton, T.H.E.; Schmidt, S.; Stewart, G.R. The N-15 natural abundance (delta N-15) of ecosystem samples reflects measures of water availability. Aust. J. Plant Physiol. 1999, 26, 185–199. [Google Scholar] [CrossRef]
- Fujiyoshi, L.; Sugimoto, A.; Tsukuura, A.; Kitayama, A.; Caceres, M.L.L.; Mijidsuren, B.; Saraadanbazar, A.; Tsujimura, M. Spatial variations in larch needle and soil N-15 at a forest-grassland boundary in northern Mongolia. Isot. Environ. Health Stud. 2017, 53, 54–69. [Google Scholar] [CrossRef]
- Makarov, M.I.; Malysheva, T.I.; Cornelissen, J.H.C.; van Logtestijn, R.S.P.; Glasser, B. Consistent patterns of N-15 distribution through soil profiles in diverse alpine and tundra ecosystems. Soil Biol. Biochem. 2008, 40, 1082–1089. [Google Scholar] [CrossRef]
- Ohta, T.; Kotani, A.; Iijima, Y.; Maximov, T.C.; Ito, S.; Hanamura, M.; Kononov, A.V.; Maximov, A.P. Effects of waterlogging on water and carbon dioxide fluxes and environmental variables in a Siberian larch forest, 1998–2011. Agric. For. Meteorol. 2014, 188, 64–75. [Google Scholar] [CrossRef]
- Iijima, Y.; Ohta, T.; Kotani, A.; Fedorov, A.N.; Kodama, Y.; Maximov, T.C. Sap flow changes in relation to permafrost degradation under increasing precipitation in an eastern Siberian larch forest. Ecohydrology 2014, 7, 177–187. [Google Scholar] [CrossRef]
- Kotani, A.; Saito, A.; Kononov, A.V.; Petrov, R.E.; Maximov, T.C.; Iijima, Y.; Ohta, T. Impact of unusually wet permafrost soil on understory vegetation and CO2 exchange in a larch forest in eastern Siberia. Agric. For. Meteorol. 2019, 265, 295–309. [Google Scholar] [CrossRef]
- Shin, N.; Kotani, A.; Sato, T.; Sugimoto, A.; Maximov, T.C.; Nogovitcyn, A.; Miyamoto, Y.; Kobayashi, H.; Tei, S. Direct measurement of leaf area index in a deciduous needle-leaf forest, eastern Siberia. Polar Sci. 2020, 25, 100550. [Google Scholar] [CrossRef]
- Suzuki, R.; Nomaki, T.; Yasunari, T. Spatial distribution and its seasonality of satellite-derived vegetation index (NDVI) and climate in Siberia. Int. J. Climatol. 2001, 21, 1321–1335. [Google Scholar] [CrossRef]
- Bahru, T.; Ding, Y.L. Effect of stand density, canopy leaf area index and growth variables on Dendrocalamus brandisii (Munro) Kurz litter production at Simao District of Yunnan Province, southwestern China. Glob. Ecol. Conserv. 2020, 23, e01051. [Google Scholar] [CrossRef]
- Will, R.E.; Narahari, N.V.; Shiver, B.D.; Teskey, R.O. Effects of planting density on canopy dynamics and stem growth for intensively managed loblolly pine stands. For. Ecol. Manag. 2005, 205, 29–41. [Google Scholar] [CrossRef]
- Loranty, M.M.; Davydov, S.P.; Kropp, H.; Alexander, H.D.; Mack, M.C.; Natali, S.M.; Zimov, N.S. Vegetation Indices Do Not Capture Forest Cover Variation in Upland Siberian Larch Forests. Remote Sens. 2018, 10, 1686. [Google Scholar] [CrossRef] [Green Version]
- Dearborn, K.D.; Baltzer, J.L. Unexpected greening in a boreal permafrost peatland undergoing forest loss is partially attributable to tree species turnover. Glob. Chang. Biol. 2021, 27, 2867–2882. [Google Scholar] [CrossRef]
- Hall, F.G.; Shimabukuro, Y.E.; Huemmrich, K.F. Remote-sensing of forest biophysical structure using mixture decomposition and geometric reflectance models. Ecol. Appl. 1995, 5, 993–1013. [Google Scholar] [CrossRef]
- Li, F.; Sugimoto, A. Effect of waterlogging on carbon isotope discrimination during photosynthesis in Larix gmelinii. Isot. Environ. Health Stud. 2018, 54, 63–77. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Forest Types | NDVI (Mean ± SD) | RF-1 | RF-2 | DF | RF-2 & DF + |
|---|---|---|---|---|---|---|
| 4 June | TF | 0.71 ± 0.01 | <0.001 *** | 0.008 ** | 0.012 * | <0.001 *** |
| RF-1 | 0.66 ± 0.03 | 0.002 ** | 0.026 * | <0.001 *** | ||
| RF-2 | 0.51 ± 0.06 | 0.533 ns | ||||
| DF | 0.46 ± 0.01 | |||||
| RF-2 & DF | 0.49 ± 0.06 | |||||
| 31 July | TF | 0.74 ± 0.01 | 0.02 * | <0.001 *** | 0.012 * | <0.001 *** |
| RF-1 | 0.73 ± 0.01 | 0.03 * | 0.051 ′ | 0.006 ** | ||
| RF-2 | 0.72 ± 0.01 | 0.533 ns | ||||
| DF | 0.71 ± 0.01 | |||||
| RF-2 & DF | 0.72 ± 0.01 | |||||
| 7 August | TF | 0.78 ± 0.01 | 0.04 * | <0.001 *** | 0.012 * | <0.001 *** |
| RF-1 | 0.77 ± 0.01 | 0.006 ** | 0.026 * | <0.001 *** | ||
| RF-2 | 0.75 ± 0.01 | 0.533 ns | ||||
| DF | 0.74 ± 0.00 | |||||
| RF-2 & DF | 0.74 ± 0.01 | |||||
| 23 August | TF | 0.71 ± 0.01 | <0.001 *** | <0.001 *** | 0.012 * | <0.001 *** |
| RF-1 | 0.69 ± 0.01 | 0.002 ** | 0.026 * | <0.001 *** | ||
| RF-2 | 0.65 ± 0.01 | 0.533 ns | ||||
| DF | 0.65 ± 0.00 | |||||
| RF-2 & DF | 0.65 ± 0.01 | |||||
| 17 September | TF | 0.44 ± 0.01 | 0.02 * | 0.003 ** | 0.012 * | <0.001 *** |
| RF-1 | 0.42 ± 0.02 | 0.21 ns | 0.231 ns | 0.17 ns | ||
| RF-2 | 0.41 ± 0.02 | 1 ns | ||||
| DF | 0.42 ± 0.00 | |||||
| RF-2 & DF | 0.41 ± 0.02 |
| Day | Foliar δ13C (‰) | Foliar δ13N (‰) | Foliar C/N | ||||||
|---|---|---|---|---|---|---|---|---|---|
| NDVI = b1x + b0 | R2 | p | NDVI = b1x + b0 | R2 | p | NDVI = b1x + b0 | R2 | p | |
| 4 June | −0.240x − 0.033 | 0.15 | 0.04 * | <0.01 | 0.93 ns | 0.006x + 0.465 | 0.39 | <0.01 ** | |
| 31 July | 0.01 | 0.64 ns | 0.007x + 0.769 | 0.16 | 0.04 * | 0.04 | 0.30 ns | ||
| 7 August | 0.01 | 0.64 ns | 0.10 | 0.11 ns | 0.06 | 0.20 ns | |||
| 23 August | 0.06 | 0.20 ns | 0.00 | 1.00 ns | 0.002x + 0.618 | 0.39 | <0.01 ** | ||
| 17 September | 0.00 | 0.90 ns | 0.01 | 0.63 ns | 0.002x + 0.376 | 0.15 | 0.04 * | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogovitcyn, A.; Shakhmatov, R.; Morozumi, T.; Tei, S.; Miyamoto, Y.; Shin, N.; Maximov, T.C.; Sugimoto, A. Changes in Forest Conditions in a Siberian Larch Forest Induced by an Extreme Wet Event. Forests 2022, 13, 1331. https://doi.org/10.3390/f13081331
Nogovitcyn A, Shakhmatov R, Morozumi T, Tei S, Miyamoto Y, Shin N, Maximov TC, Sugimoto A. Changes in Forest Conditions in a Siberian Larch Forest Induced by an Extreme Wet Event. Forests. 2022; 13(8):1331. https://doi.org/10.3390/f13081331
Chicago/Turabian StyleNogovitcyn, Aleksandr, Ruslan Shakhmatov, Tomoki Morozumi, Shunsuke Tei, Yumiko Miyamoto, Nagai Shin, Trofim C. Maximov, and Atsuko Sugimoto. 2022. "Changes in Forest Conditions in a Siberian Larch Forest Induced by an Extreme Wet Event" Forests 13, no. 8: 1331. https://doi.org/10.3390/f13081331