
Strongly Active Responses of Pinus tabuliformis Carr. and Sophora viciifolia Hance to CO2 Enrichment and Drought Revealed by Tree-Ring Isotopes on the Central China Loess Plateau


Abstract
:1. Introduction
2. Materials and Methods
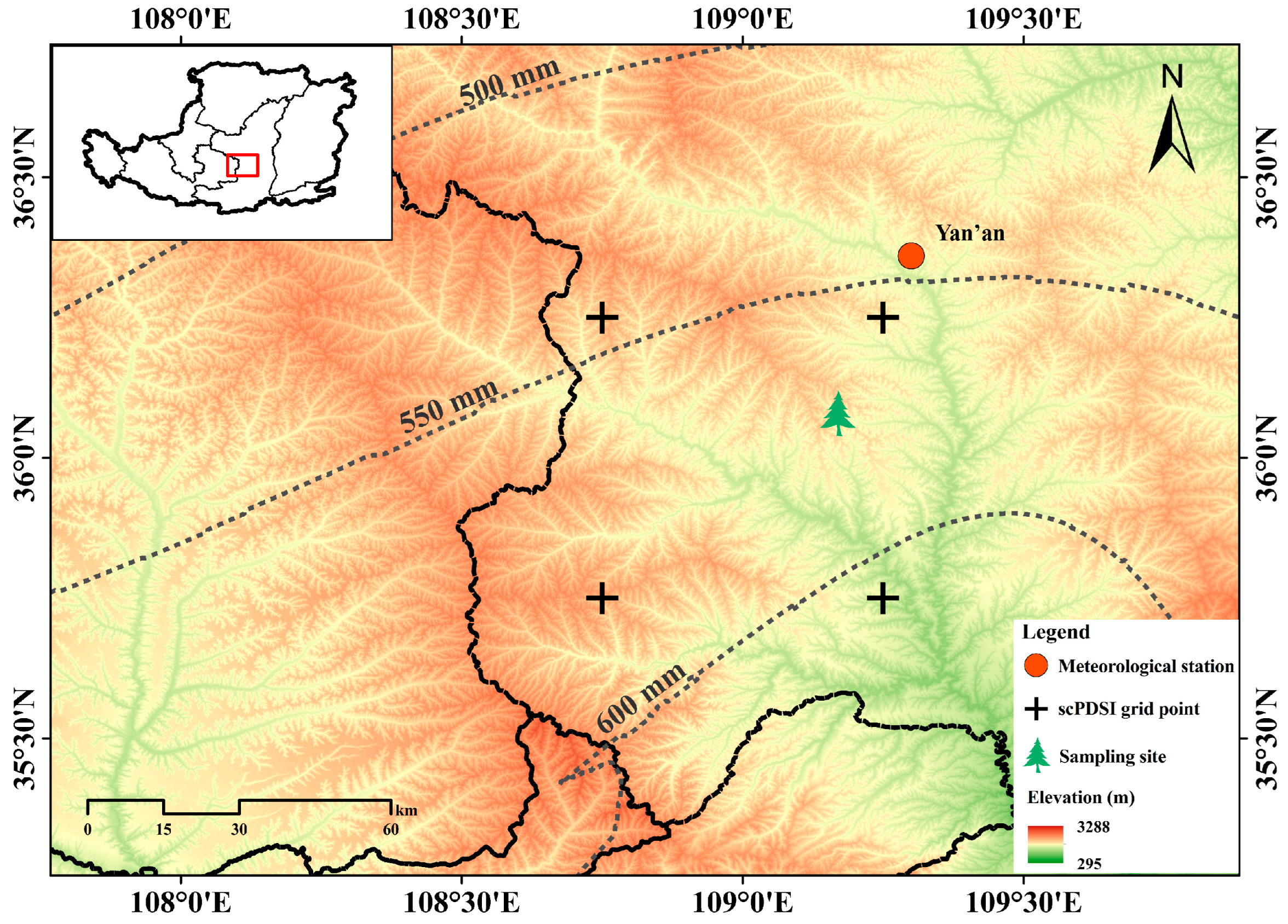
2.1. Study Area and Climate
2.2. Sample Preparation and Isotopic Measurements
2.3. Definitions and Basic Equations
2.4. Data Analyses
3. Results
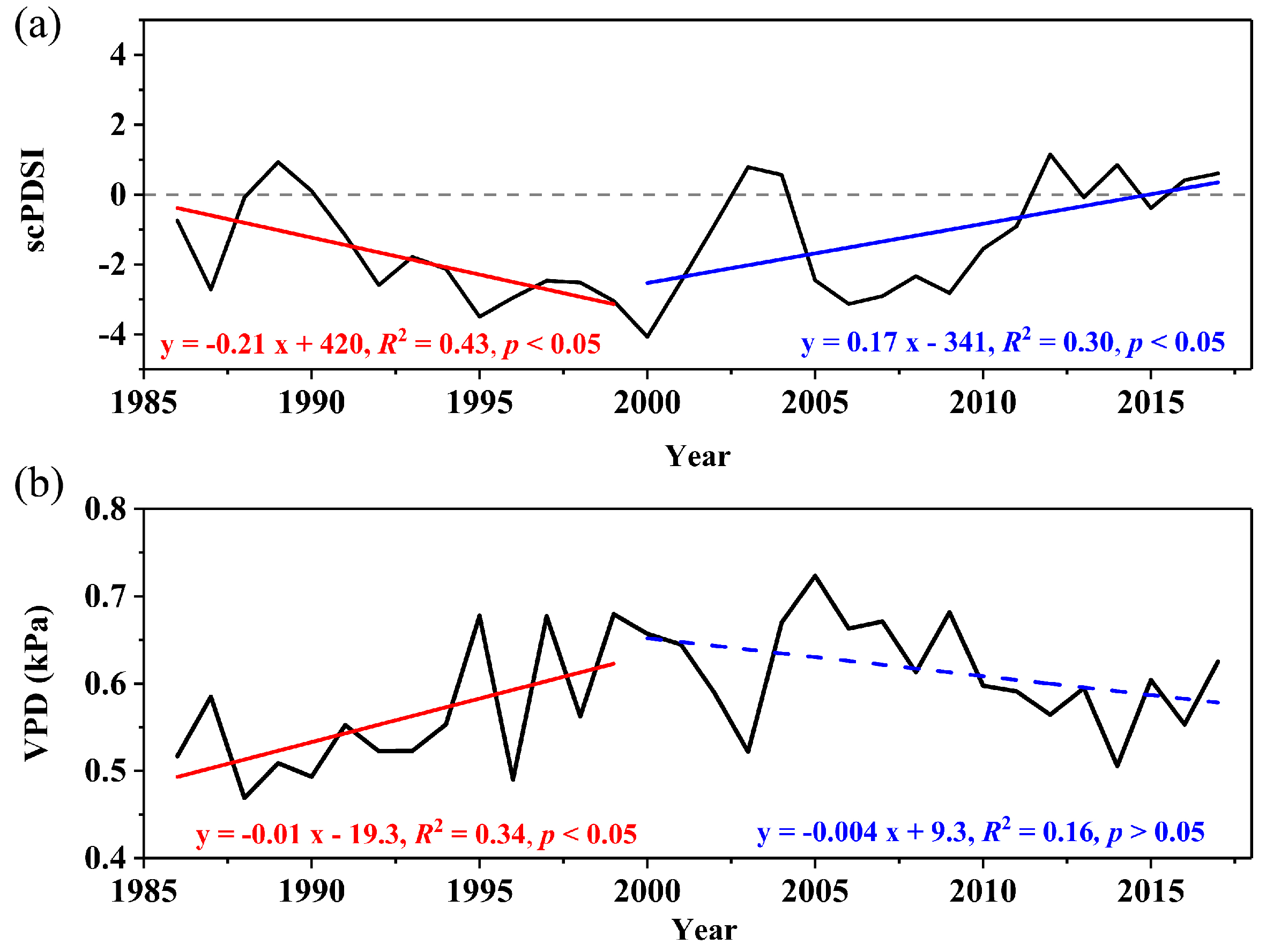
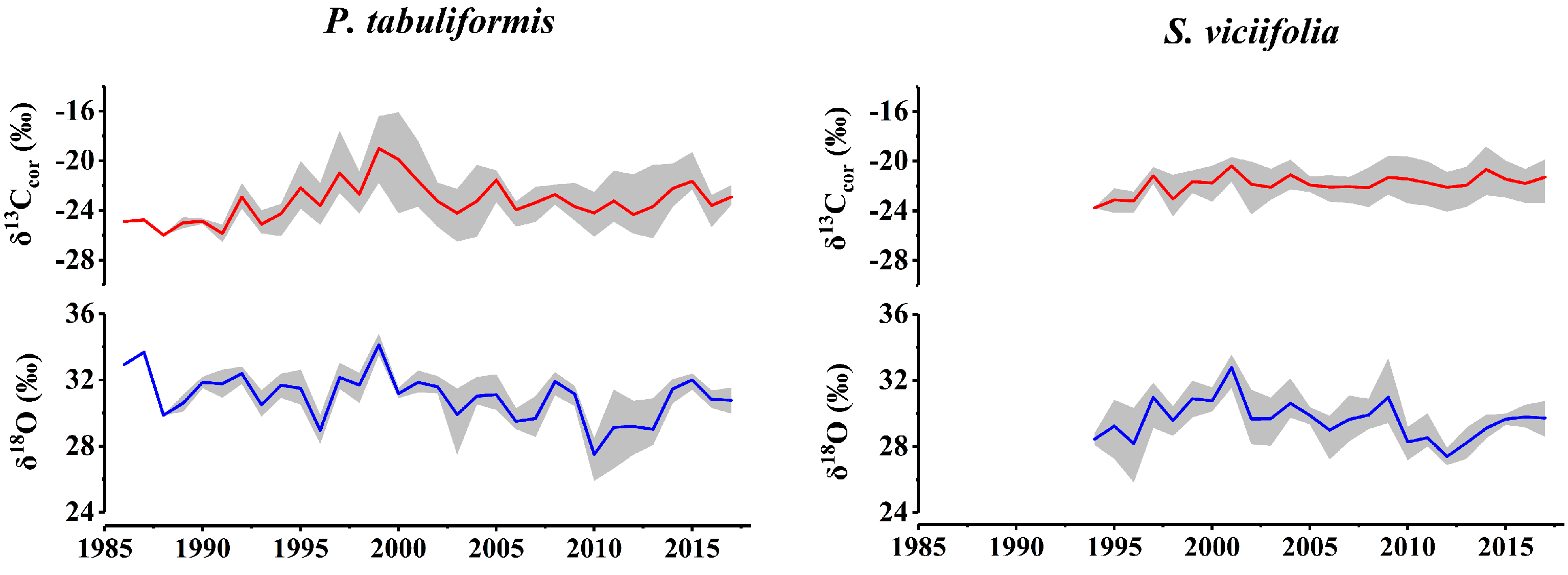
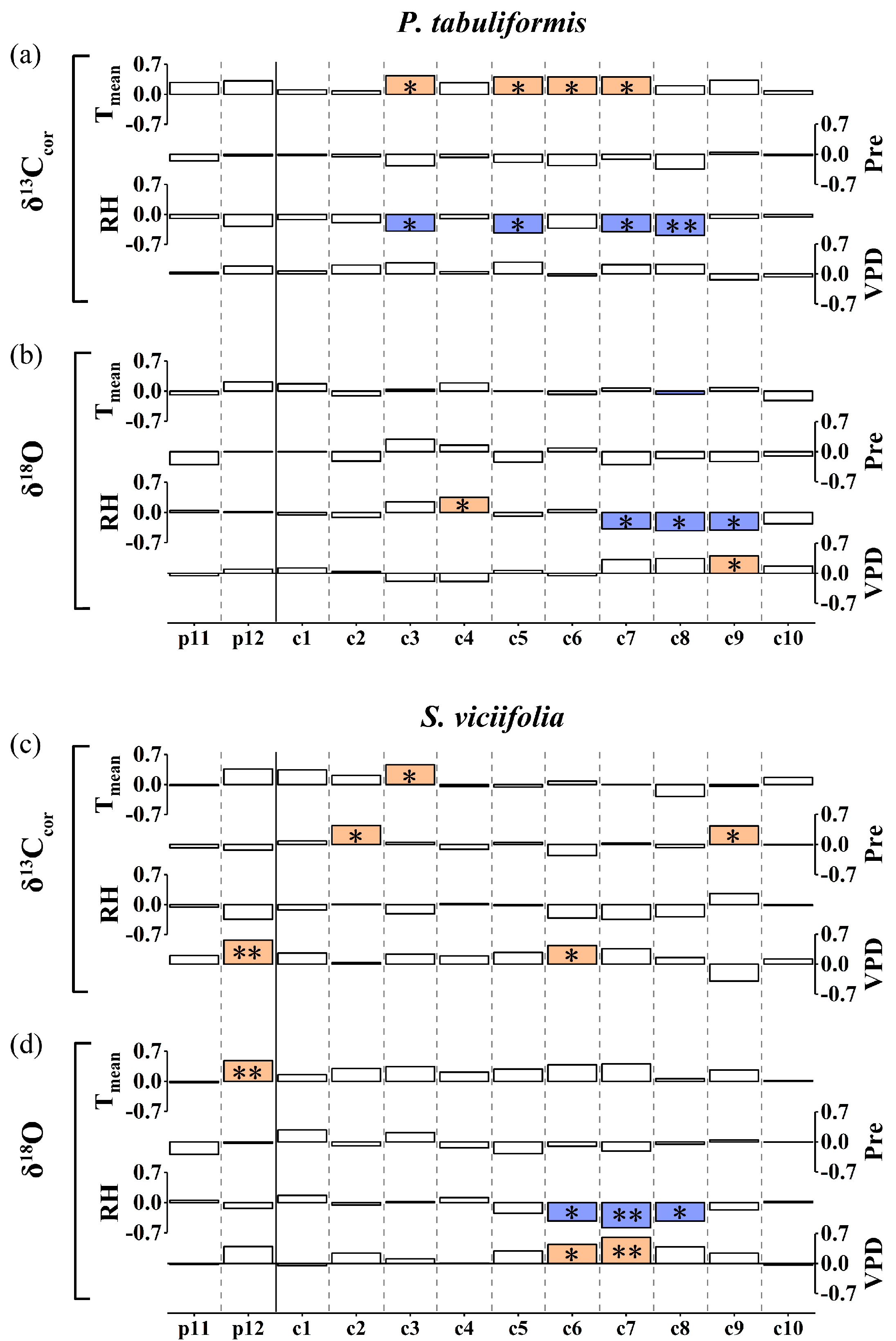
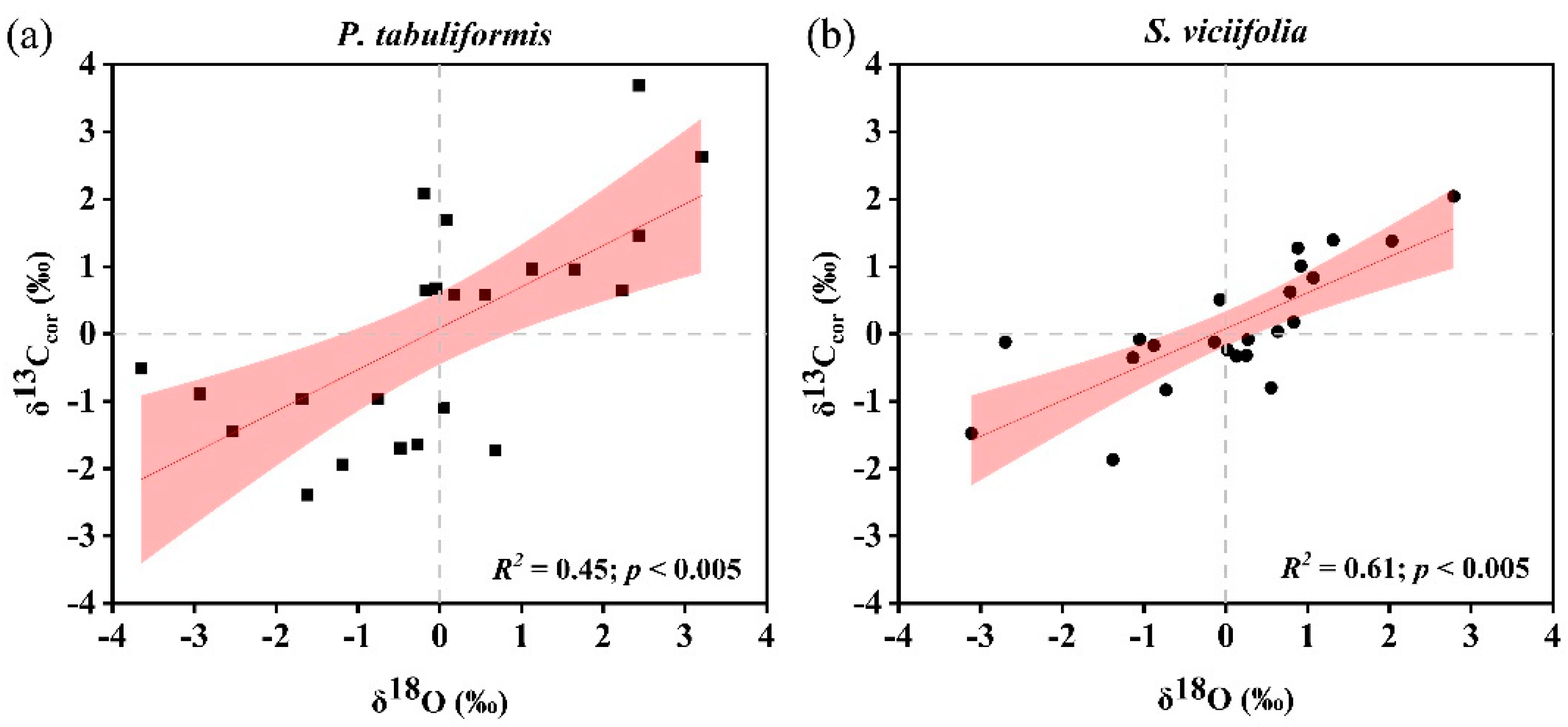
3.1. Characteristics and Climate Responses of δ13Ccor and δ18O
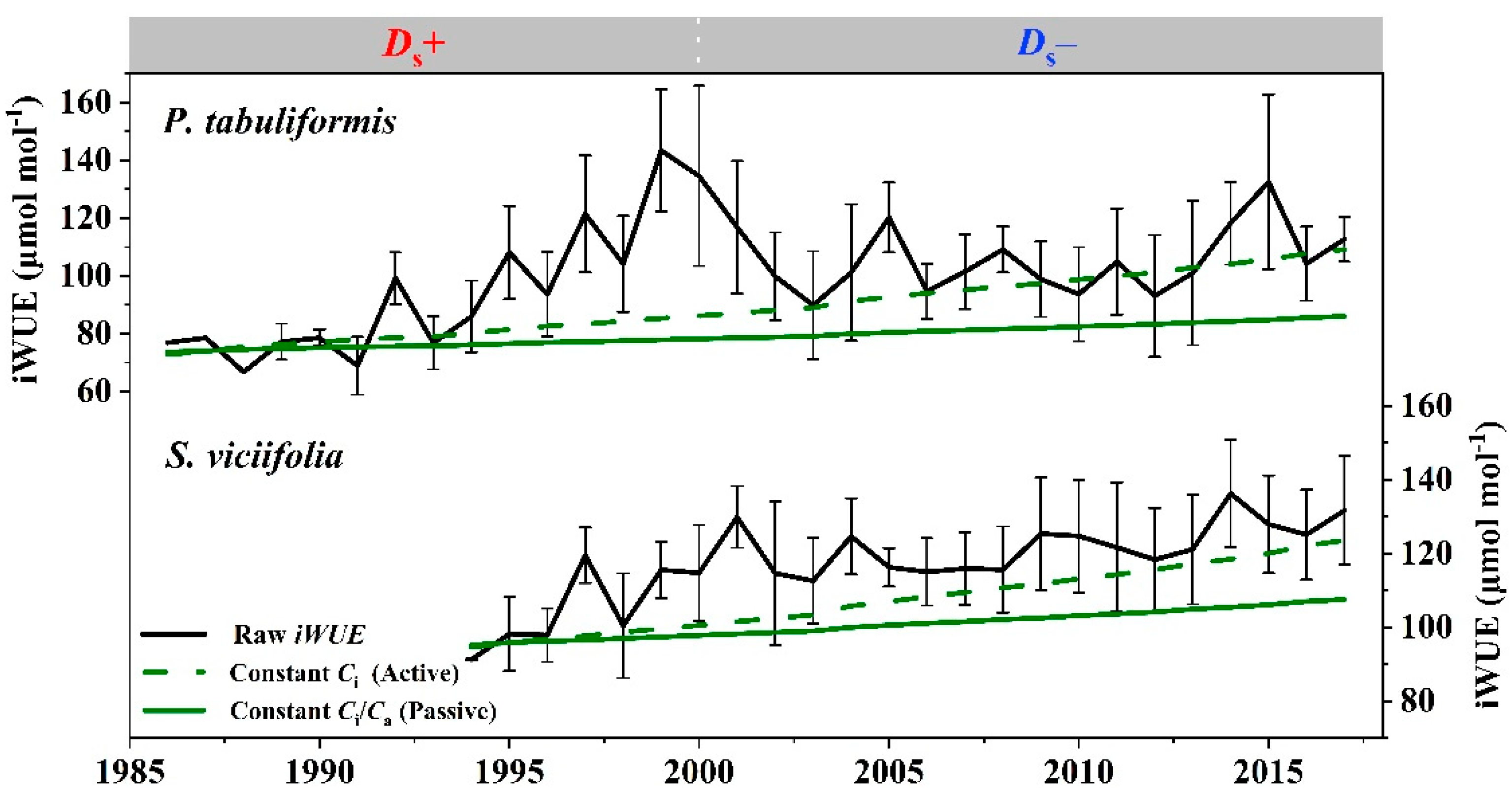
3.2. Trends of iWUE and Dual-Isotopes Relationships
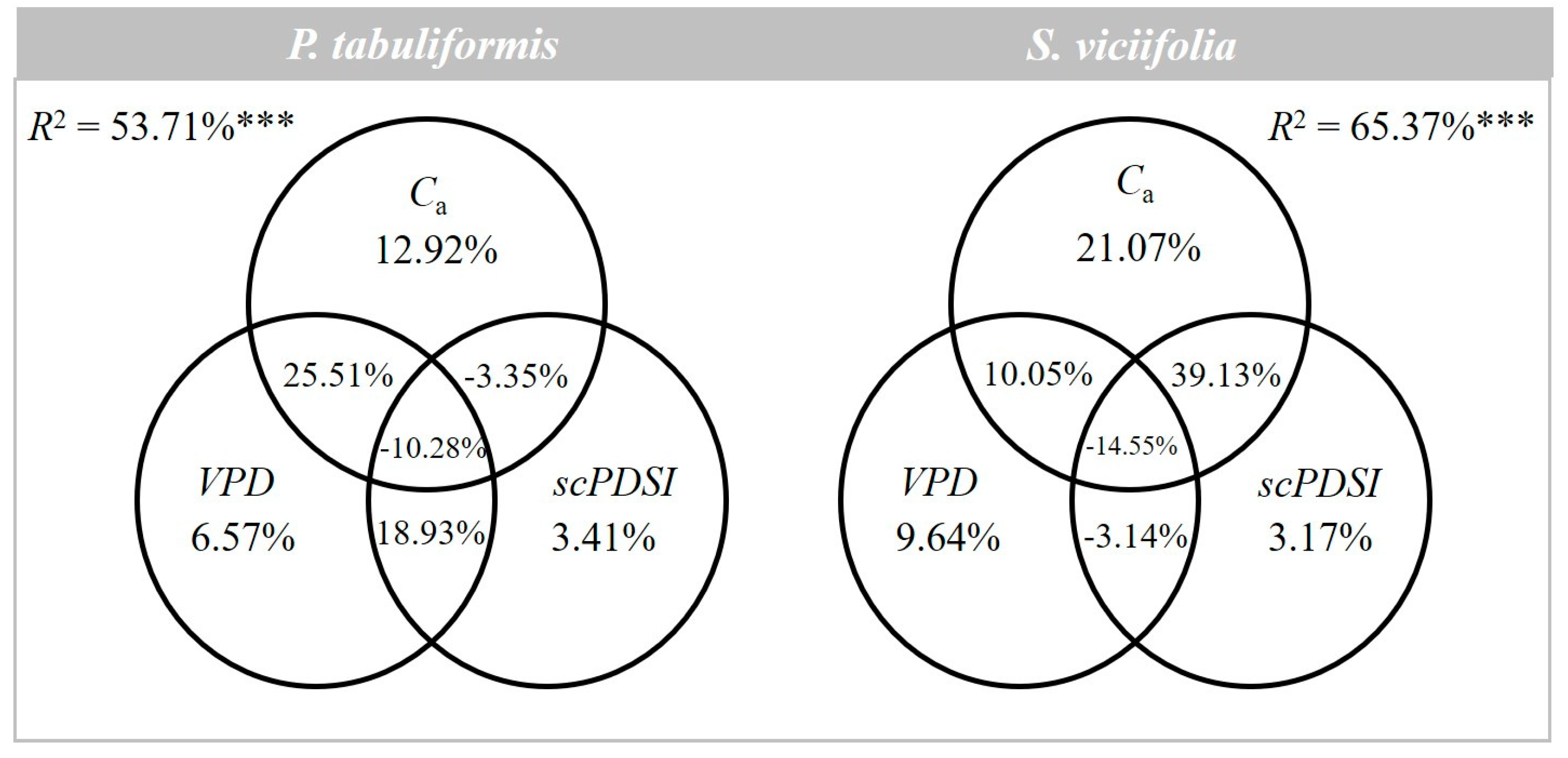
3.3. Contribution of CO2 and Drought to iWUE
4. Discussion
4.1. Climate Drivers of δ13Ccor and δ18O
4.2. Stomatal Conductance Dominates the Regulation of iWUE
4.3. The Contribution of CO2 and Drought to iWUE
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WMO. WMO Greenhouse Gas Bulletin (GHG Bulletin)—No.16: The State of Greenhouse Gases in the Atmosphere Based on Global Observations through 2019; World Meteorological Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Dai, A.G. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2012, 3, 52–58. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Mathias, J.M.; Thomas, R.B. Global tree intrinsic water use efficiency is enhanced by increased atmospheric CO2 and modulated by climate and plant functional types. Proc. Natl. Acad. Sci. USA 2021, 118, e2014286118. [Google Scholar] [CrossRef] [PubMed]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; McCarthy, H.R.; Norby, R.J.; Cotrufo, M.F. Elevated CO2 increases tree-level intrinsic water use efficiency: Insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restaino, C.M.; Peterson, D.L.; Littell, J. Increased water deficit decreases Douglas fir growth throughout western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 9557–9562. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Williams, A.P.; Berg, A.M.; Cook, B.I.; Zhang, Y.; Hagemann, S.; Lorenz, R.; Seneviratne, S.I.; Gentine, P. Land-atmosphere feedbacks exacerbate concurrent soil drought and atmospheric aridity. Proc. Natl. Acad. Sci. USA 2019, 116, 18848–18853. [Google Scholar] [CrossRef] [Green Version]
- Keenan, T.F.; Prentice, C.; Canadell, J.G.; Williams, C.A.; Wang, H.; Raupach, M.; Collatz, G.J. Recent pause in the growth rate of atmospheric CO2 due to enhanced terrestrial carbon uptake. Nat. Commun. 2016, 7, 13428. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Fu, B.J.; Wang, S.; Stringer, L.C.; Wang, Y.P.; Li, Z.D.; Liu, Y.X.; Zhou, W.X. Drivers and impacts of changes in China’s drylands. Nat. Rev. Earth Environ. 2021, 2, 858–873. [Google Scholar] [CrossRef]
- Zheng, K.; Wei, J.Z.; Pei, J.Y.; Cheng, H.; Zhang, X.L.; Huang, F.Q.; Li, F.M.; Ye, J.S. Impacts of climate change and human activities on grassland vegetation variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef]
- Chen, Y.P.; Wang, K.B.; Lin, Y.S.; Shi, W.Y.; Song, Y.; He, X.H. Balancing green and grain trade. Nat. Geosci. 2015, 8, 739–741. [Google Scholar] [CrossRef]
- Feng, X.M.; Fu, B.J.; Piao, S.L.; Wang, S.; Ciais, P.; Zeng, Z.Z.; Lü, Y.H.; Zeng, Y.; Li, Y.; Jiang, X.H.; et al. Revegetation in China’s Loess Plateau is approaching sustainable water resource limits. Nat. Clim. Chang. 2016, 6, 1019–1022. [Google Scholar] [CrossRef]
- Yao, Y.B.; Wang, R.Y.; Yang, J.H.; Yue, P.; Lu, D.R.; Xiao, G.J.; Wang, Y.; Liu, L.C. Changes in terrestrial surface dry and wet conditions on the Loess Plateau (China) during the last half century. J. Arid. Land. 2013, 5, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.J.; Wu, Y.P.; Shi, Z.Y.; Yu, M.Z.; Zhao, F.B.; Guan, Y.H. Quantifying spatiotemporal variations in soil moisture driven by vegetation restoration on the Loess Plateau of China. J. Hydrol. 2021, 600, 126580. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant. Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Barbour, M.M.; Schurr, U.; Henry, B.K.; Wong, S.C.; Farquhar, G.D. Variation in the oxygen isotope ratio of phloem sap sucrose from castor bean. Evidence in support of the Péclet effect. Plant Physiol. 2000, 123, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.M.; Roden, J.S.; Farquhar, G.D.; Ehleringer, J.R. Expressing leaf water and cellulose oxygen isotope ratios as enrichment above source water reveals evidence of a Péclet effect. Oecologia 2004, 138, 426–435. [Google Scholar] [CrossRef]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef]
- Gessler, A.; Cailleret, M.; Joseph, J.; Schönbeck, L.; Schaub, M.; Lehmann, M.; Treydte, K.; Rigling, A.; Timofeeva, G.; Saurer, M. Drought induced tree mortality—A tree-ring isotope based conceptual model to assess mechanisms and predispositions. New Phytol. 2018, 219, 485–490. [Google Scholar] [CrossRef]
- Rodriguez-Caton, M.; Andreu-Hayles, L.; Morales, M.S.; Daux, V.; Christie, D.A.; Coopman, R.E.; Alvarez, C.; Rao, M.P.; Aliste, D.; Flores, F.; et al. Different climate sensitivity for radial growth, but uniform for tree-ring stable isotopes along an aridity gradient in Polylepis tarapacana, the world’s highest elevation tree species. Tree Physiol. 2021, 41, 1353–1371. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Li, Q.; Cai, Q.F.; Song, H.M.; Sun, C.F.; Liu, R.S.; Mei, R.C. Asian summer monsoon-related relative humidity recorded by tree ring δ18O during last 205 years. J. Geophys. Res. Atmos. 2019, 124, 9824–9838. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Li, Q.; Song, H.M.; Sun, C.F.; Fang, C.X. An Asian summer monsoon-related relative humidity record from tree-ring δ18O in Gansu Province, North China. Atmosphere 2020, 11, 984. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, M.; Li, Q.; Song, H.M.; Liu, R.S. Tree-ring δ18O-based July–August relative humidity reconstruction on Mt. Shimen, China, for the last 400 years. Atmos. Res. 2020, 243, 105024. [Google Scholar] [CrossRef]
- Li, Q.; Nakatsuka, T.; Kawamura, K.; Liu, Y.; Song, H.M. Hydroclimate variability in the North China Plain and its link with El Niño-Southern Oscillation since 1784 A.D.: Insights from tree-ring cellulose δ18O. J. Geophys. Res. Atmos. 2011, 116, D22106. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, H.Y.; Leavitt, S.; Cressey, E.L.; Quine, T.A.; Shi, J.F.; Shi, S.Y. Tree-ring δ18O identifies similarity in timing but differences in depth of soil water uptake by trees in mesic and arid climates. Agric. For. Meteorol. 2021, 308, 108569. [Google Scholar] [CrossRef]
- Koretsune, S.; Fukuda, K.; Chang, Z.Y.; Shi, F.C.; Ishida, A. Effective rainfall seasons for interannual variation in δ13C and tree-ring width in early and late wood of Chinese pine and black locust on the Loess Plateau, China. J. For. Res. 2009, 14, 88–94. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, X.; Li, M.; Zhang, J.J.; Cao, Y. Climate-growth pattern of Pinus tabulaeformis plantations and their resilience to drought events in the Loess Plateau. For. Ecol. Manag. 2021, 499, 119642. [Google Scholar] [CrossRef]
- Wang, J.; Lu, N.; Fu, B.J. Inter-comparison of stable isotope mixing models for determining plant water source partitioning. Sci. Total Environ. 2019, 666, 685–693. [Google Scholar] [CrossRef]
- Fan, J.; Wang, Q.J.; Jones, S.B.; Shao, M.A. Soil water depletion and recharge under different land cover in China’s Loess Plateau. Ecohydrology 2016, 9, 396–406. [Google Scholar] [CrossRef]
- Zeng, X.M.; Sun, S.W.; Wang, Y.Y.; Chang, Y.X.; Tao, X.X.; Hou, M.Y.; Wang, W.C.; Liu, X.H.; Zhang, L.N. Does resin tapping affect the tree-ring growth and climate sensitivity of the Chinese pine (Pinus tabuliformis) in the Loess Plateau, China? Dendrochronologia 2021, 65, 125800. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics; Springer: New York, NY, USA, 1977. [Google Scholar]
- Palmer, W.C. Weather Bureau Research Paper No. 45. In Meteorological Drought; US Department of Commerce: Washington, DC, USA, 1965. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer: New York, NY, USA, 1990. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Li, X.Q.; Zhang, L.N.; Zeng, X.M.; Wang, K.Y.; Wang, Y.B.; Lu, Q.Q.; Liu, X.H. Different response of conifer and shrubs radial growth to climate in the middle Loess Plateau. Acta Ecol. Sin. 2020, 40, 5685–5697. (In Chinese) [Google Scholar]
- Liu, X.H.; Zhao, L.J.; Voelker, S.; Xu, G.B.; Zeng, X.M.; Zhang, X.W.; Zhang, L.N.; Sun, W.Z.; Zhang, Q.L.; Wu, G.J.; et al. Warming and CO2 enrichment modified the ecophysiological responses of Dahurian larch and Mongolia pine during the past century in the permafrost of northeastern China. Tree Physiol. 2019, 39, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, S.W.; Danzer, S.R. Method for batch processing small wood samples to holocellulose for stable-carbon isotope analysis. Anal. Chem. 1993, 65, 87–89. [Google Scholar] [CrossRef]
- Evans, M.N.; Selmer, K.J.; Breeden, B.T.; Lopatka, A.S.; Plummer, R.E. Correction algorithm for online continuous flow δ13C and δ18O carbonate and cellulose stable isotope analyses. Geochem. Geophys. Geosystems 2016, 17, 3580–3588. [Google Scholar] [CrossRef] [Green Version]
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant Carbon-Water Relations; Academic Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Savard, M.M.; Begin, C.; Marion, J. Response strategies of boreal spruce trees to anthropogenic changes in air quality and rising pCO2. Environ. Pollut. 2020, 261, 114209. [Google Scholar] [CrossRef]
- Barichivich, J.; Peylin, P.; Launois, T.; Daux, V.; Risi, C.; Jeong, J.; Luyssaert, S. A triple tree-ring constraint for tree growth and physiology in a global land surface model. Biogeosciences 2020, 18, 3781–3803. [Google Scholar] [CrossRef]
- Voelker, S.L.; Brooks, J.R.; Meinzer, F.C.; Anderson, R.; Bader, M.K.; Battipaglia, G.; Becklin, K.M.; Beerling, D.; Bert, D.; Betancourt, J.L.; et al. A dynamic leaf gas-exchange strategy is conserved in woody plants under changing ambient CO2: Evidence from carbon isotope discrimination in paleo and CO2 enrichment studies. Glob. Chang. Biol. 2016, 22, 889–902. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.T.W.; Schweingruber, F.H. Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Glob. Chang. Biol. 2004, 10, 2109–2120. [Google Scholar] [CrossRef]
- Seibold, D.R.; McPhee, R.D. Commonality analysis: A method for decomposing explained variance in multiple regression analyses. Hum. Commun. Res. 1979, 5, 355–365. [Google Scholar] [CrossRef]
- Ray-Mukherjee, J.; Nimon, K.; Mukherjee, S.; Morris, D.W.; Slotow, R.; Hamer, M. Using commonality analysis in multiple regressions: A tool to decompose regression effects in the face of multicollinearity. Methods Ecol. Evol. 2014, 5, 320–328. [Google Scholar] [CrossRef]
- Keyimu, M.; Li, Z.S.; Fu, B.J.; Chen, W.L.; Wei, J.S.; Jiao, L.; Gao, G.Y.; Lü, Y.H. Spatial differences in the radial growth responses of black locust (Robinia pseudoacacia Linn.) to climate on the Loess Plateau, China. Dendrochronologia 2021, 67, 125832. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Nakatsuka, T.; Zhang, Q.B.; Ohnishi, K.; Sakai, A.; Kobayashi, O.; Pan, Y.N.; Song, H.M.; Liu, R.S.; et al. Oxygen stable isotopes of a network of shrubs and trees as high-resolution plaeoclimatic proxies in Northwestern China. Agric. For. Meteorol. 2020, 285, 107929. [Google Scholar] [CrossRef]
- Wang, W.Z.; McDowell, N.G.; Liu, X.H.; Xu, G.B.; Wu, G.J.; Zeng, X.M.; Wang, G.X. Contrasting growth responses of Qilian juniper (Sabina przewalskii) and Qinghai spruce (Picea crassifolia) to CO2 fertilization despite common water-use efficiency increases at the northeastern Qinghai–Tibetan plateau. Tree Physiol. 2020, 41, 992–1003. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.C.; Li, Q.; Song, H.M.; Linderholm, H.W.; Leavitt, S.W.; Wang, R.Y.; An, Z.S. Tree-ring stable carbon isotope-based May–July temperature reconstruction over Nanwutai, China, for the past century and its record of 20th century warming. Quat. Sci. Rev. 2014, 93, 67–76. [Google Scholar] [CrossRef]
- Treydte, K.; Boda, S.; Graf Pannatier, E.; Fonti, P.; Frank, D.; Ullrich, B.; Saurer, M.; Siegwolf, R.; Battipaglia, G.; Werner, W.; et al. Seasonal transfer of oxygen isotopes from precipitation and soil to the tree ring: Source water versus needle water enrichment. New Phytol. 2014, 202, 772–783. [Google Scholar] [CrossRef]
- Liu, X.H.; Wang, W.Z.; Xu, G.B.; Zeng, X.M.; Wu, G.J.; Zhang, X.W.; Qin, D.H. Tree growth and intrinsic water-use efficiency of inland riparian forests in northwestern China: Evaluation via δ13C and δ18O analysis of tree rings. Tree Physiol. 2014, 34, 966–980. [Google Scholar] [CrossRef] [Green Version]
- Li, F.L.; Bao, W.K.; Liu, J.H.; Wu, N. Eco-anatomical characteristics of Sophora davidii leaves along an elevation gradient in upper Minjiang River dry valley. Chin. J. Appl. Ecol. 2006, 17, 5–10. (In Chinese) [Google Scholar]
- Liu, X.Y.; Li, J.Y.; Zhai, H.B.; Zhu, G.B. Discussion on drought resistance through hydraulic architecture of trees. J. Beijing For. Univ. 2003, 25, 48–54. (In Chinese) [Google Scholar]
- Prentice, I.C.; Dong, N.; Gleason, S.M.; Maire, V.; Wright, I.J. Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology. Ecol. Lett. 2014, 17, 82–91. [Google Scholar] [CrossRef]
- Wang, W.Z.; Liu, X.H.; An, W.L.; Xu, G.B.; Zeng, X.M. Increased intrinsic water-use efficiency during a period with persistent decreased tree radial growth in northwestern China: Causes and implications. For. Ecol. Manag. 2012, 275, 14–22. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Wanek, W.; Hietz, P. Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species. Trees 2010, 25, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Zhu, H.F.; Liu, X.H.; Liang, E.Y.; Grießinger, J.; Wu, G.J.; Li, X.X.; Bräuning, A. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural alpine timberline on the southeastern Tibetan Plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.; Siegwolf, R. Is the dual-isotope conceptual model fully operational? Tree Physiol. 2012, 32, 1179–1182. [Google Scholar] [CrossRef]
- Driscoll, A.W.; Bitter, N.Q.; Sandquist, D.R.; Ehleringer, J.R. Multidecadal records of intrinsic water-use efficiency in the desert shrub Encelia farinosa reveal strong responses to climate change. Proc. Natl. Acad. Sci. USA 2020, 117, 18161–18168. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Driscoll, A.W.; Szejner, P.; Anderegg, W.R.L.; Ehleringer, J.R. Rapid increases in shrubland and forest intrinsic water-use efficiency during an ongoing megadrought. Proc. Natl. Acad. Sci. USA 2021, 118, e2118052118. [Google Scholar] [CrossRef]
- Xiao, S.C.; Ding, A.J.; Tian, Q.Y.; Han, C.; Peng, X.M. Site- and species-specific climatic responses of two co-occurring shrubs in the temperate Alxa Desert Plateau, northwest China. Sci. Total Environ. 2019, 667, 77–85. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Shao, M.A.; Zhu, Y.J.; Liu, Z.P. Impacts of land use and plant characteristics on dried soil layers in different climatic regions on the Loess Plateau of China. Agric. For. Meteorol. 2011, 151, 437–448. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Index | Mean | SD |
|---|---|---|---|
| P. tabuliformis | δ13Ccor | −22.8 (‰) b | 1.4 (‰) |
| δ18O | 30.8 (‰) a | 1.4 (‰) | |
| iWUE | 107.6 (μmol mol−1) b | 14.8 (μmol mol−1) | |
| S. viciifolia | δ13Ccor | −21.9 (‰) a | 0.8 (‰) |
| δ18O | 29.6 (‰) b | 1.2 (‰) | |
| iWUE | 117.3 (μmol mol−1) a | 11.2 (μmol mol−1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, W.; Liu, X.; Li, X.; Zeng, X.; Zhang, L.; Wang, W.; Xu, G. Strongly Active Responses of Pinus tabuliformis Carr. and Sophora viciifolia Hance to CO2 Enrichment and Drought Revealed by Tree-Ring Isotopes on the Central China Loess Plateau. Forests 2022, 13, 986. https://doi.org/10.3390/f13070986
Ge W, Liu X, Li X, Zeng X, Zhang L, Wang W, Xu G. Strongly Active Responses of Pinus tabuliformis Carr. and Sophora viciifolia Hance to CO2 Enrichment and Drought Revealed by Tree-Ring Isotopes on the Central China Loess Plateau. Forests. 2022; 13(7):986. https://doi.org/10.3390/f13070986
Chicago/Turabian StyleGe, Wensen, Xiaohong Liu, Xiaoqin Li, Xiaomin Zeng, Lingnan Zhang, Wenzhi Wang, and Guobao Xu. 2022. "Strongly Active Responses of Pinus tabuliformis Carr. and Sophora viciifolia Hance to CO2 Enrichment and Drought Revealed by Tree-Ring Isotopes on the Central China Loess Plateau" Forests 13, no. 7: 986. https://doi.org/10.3390/f13070986