Quantifying Vegetation Stability under Drought in the Middle Reaches of Yellow River Basin, China

Abstract
:1. Introduction
2. Materials and Methods
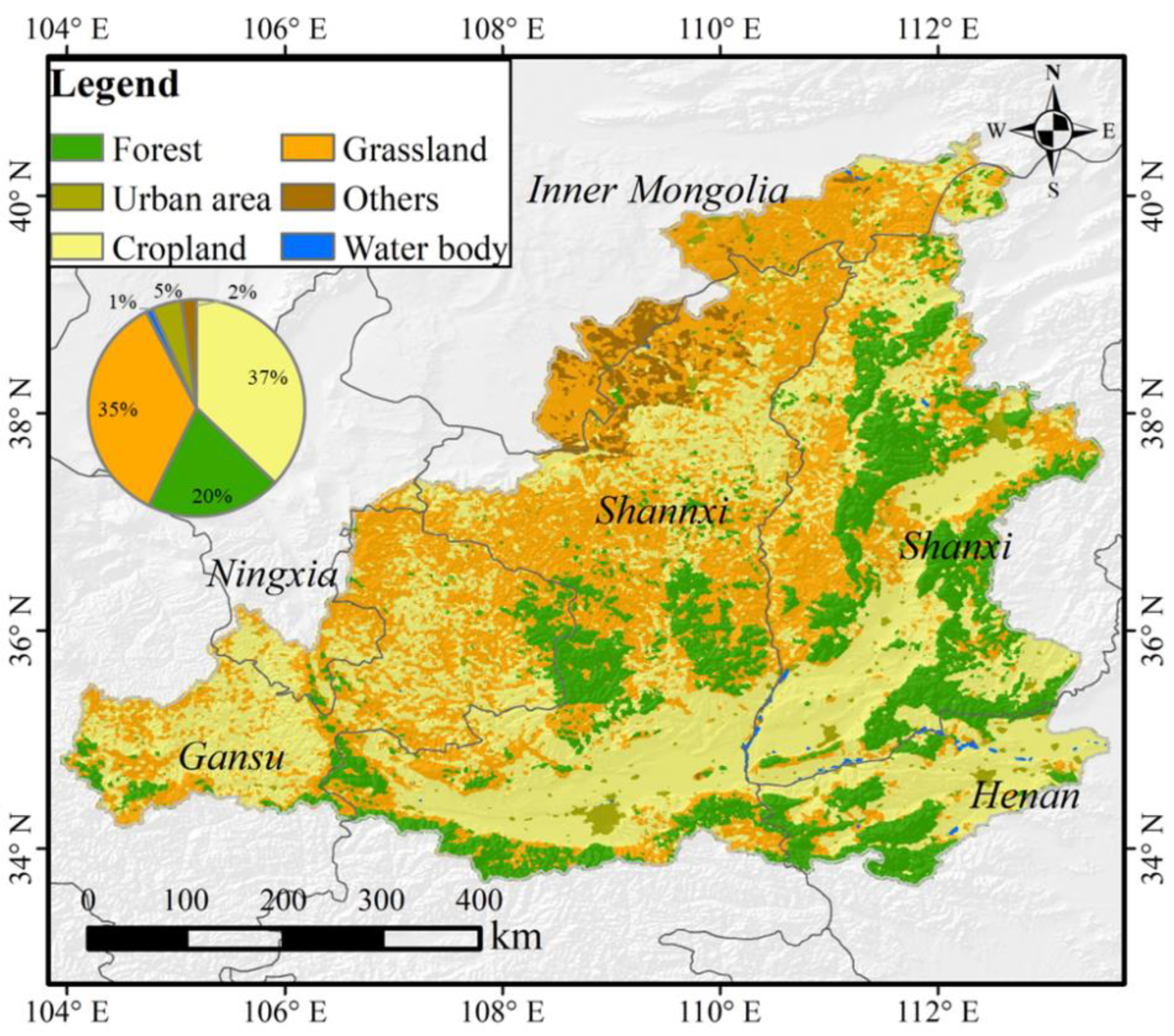
2.1. Study Area
2.2. Data Sources and Processing
2.2.1. Normalized Difference Vegetation Index
2.2.2. Standardized Precipitation Evapotranspiration Index
2.2.3. Land Cover Data
2.3. Method
2.3.1. Resistance and Resilience
2.3.2. Standardized Anomaly
2.3.3. Identification of the Drought Characterization at Grid Cell Level
2.3.4. Identification of the Vegetation Stability Parameters at Grid Cell Level
3. Results
3.1. Qualitative Description of Vegetation Stability
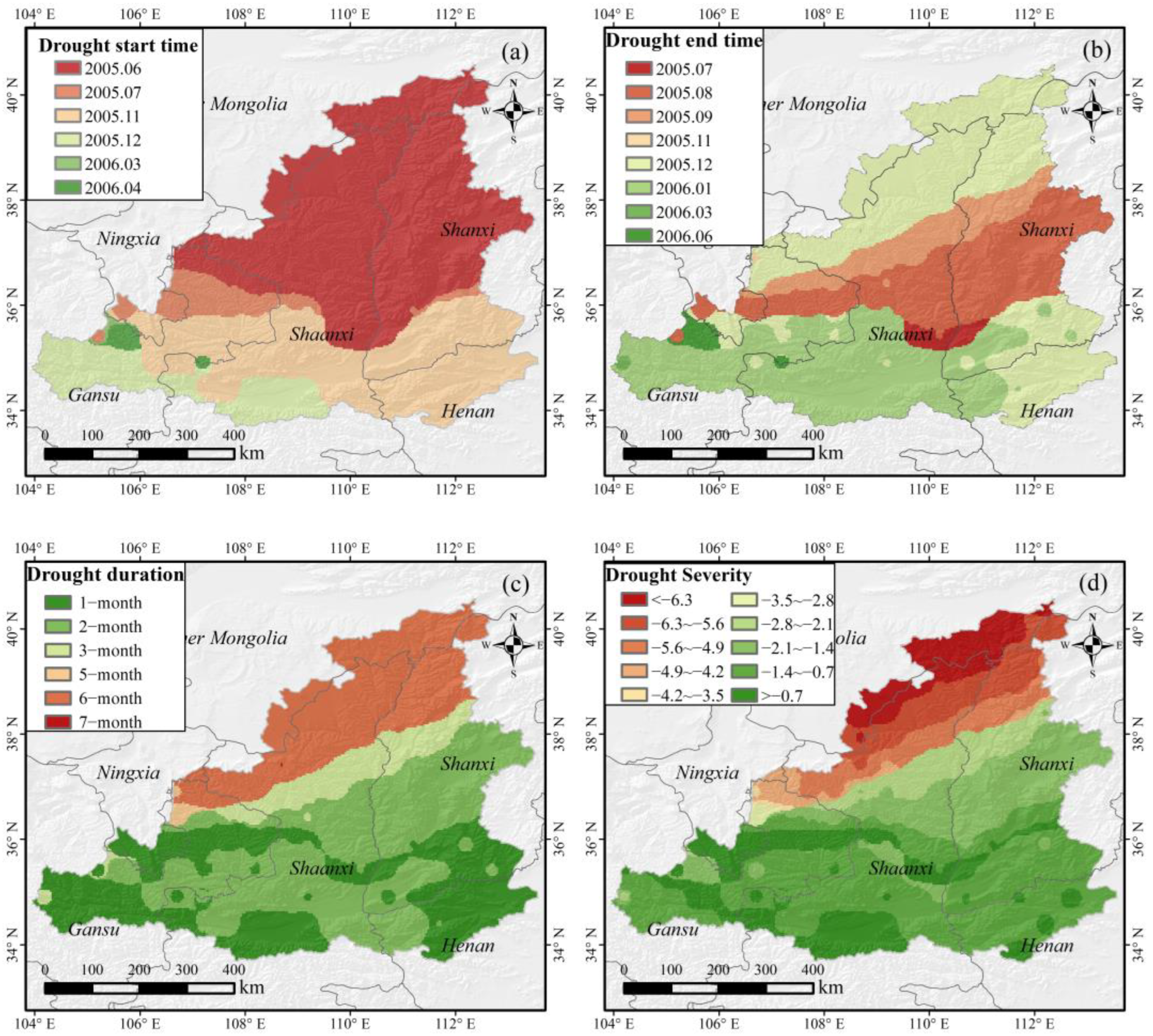
3.2. The Characteristics of the 2005/2006 Drought Event
3.3. Spatial Distribution of Vegetation Stability Parameters
4. Discussion
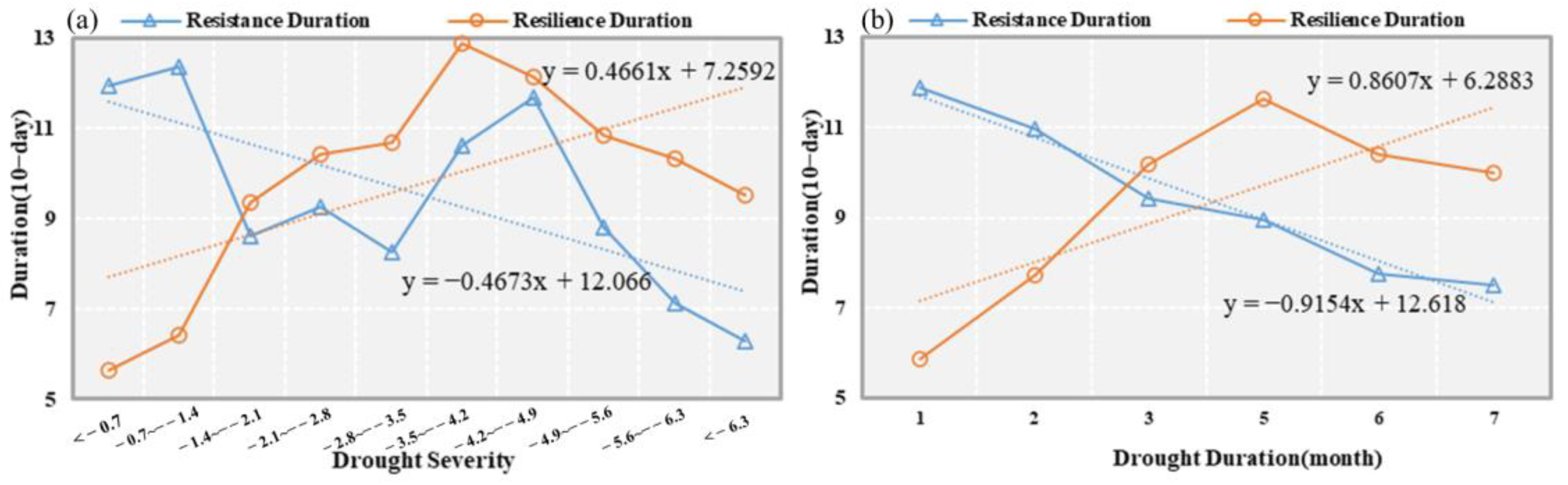
4.1. Impact of Drought Characteristics on Stability
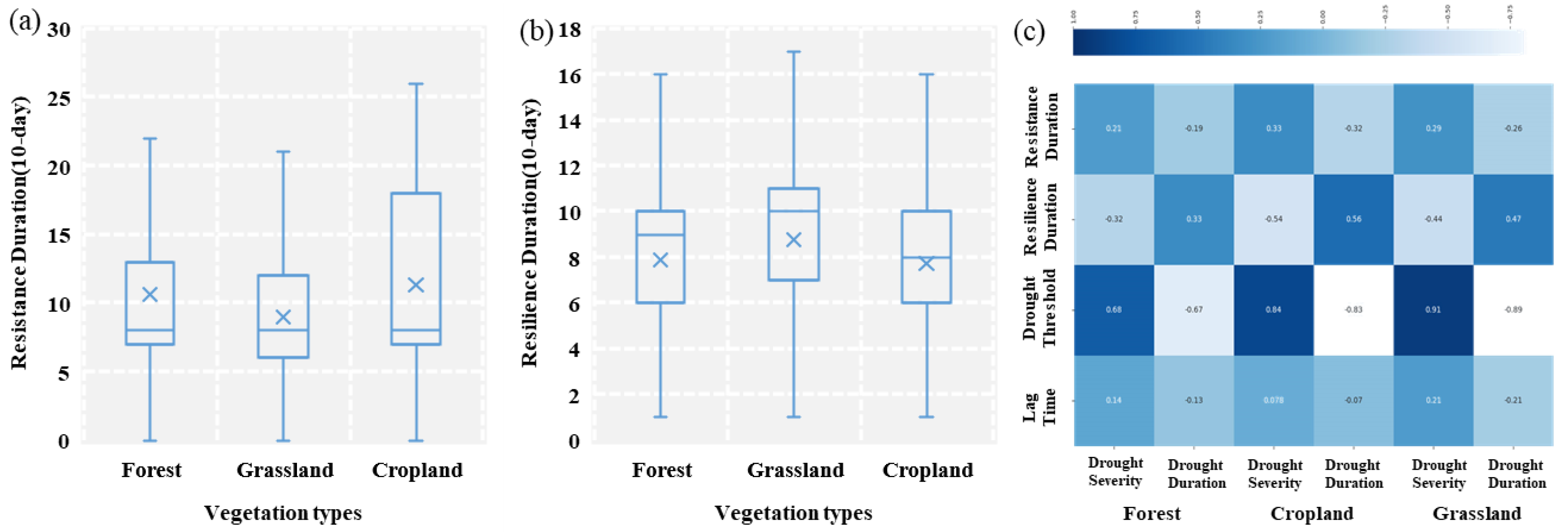
4.2. Divergent Stability in Different Vegetation Types
5. Conclusions
- The vegetation in the study area was mainly less resistance and less resilience (40.9%), which was distributed in the northwest of the study area.
- The start time of the drought is mainly concentrated in the summer and winter of 2005. The drought duration and severity present the similar space distribution characteristics. The northwestern part of the study area has a longer duration drought and severe drought severity, whereas the southeastern part is the opposite.
- Most vegetation (89.4%) can return to the normal level within 100 days after the drought. The average drought threshold in the study area is −2.6, and the lag time of drought on vegetation is concentrated in 1–3 months.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, A. Drought under global warming: A review. Wiley Interdiscip. Rev. Clim. Change 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Miao, C.; Zheng, H.; Duan, Q.; Lei, X.; Li, H. Meteorological and Hydrological Drought on the Loess Plateau, China: Evolutionary Characteristics, Impact, and Propagation. J. Geophys. Res.-Atmos. 2018, 123, 11569–11584. [Google Scholar] [CrossRef]
- Shi, M.; Yuan, Z.; Shi, X.; Li, M.; Chen, F.; Li, Y. Drought assessment of terrestrial ecosystems in the Yangtze River Basin, China. J. Clean. Prod. 2022, 362, 132234. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A. Greening of the Earth and its drivers. Nat. Clim. Change 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Keenan, F.T.; David, Y.H.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Andrew, D.R. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Ruppert, J.C.; Keith, H.; Zalmen, H.; Snyman, H.A.; Marcelo, S.; Walter, W.; Anja, L. Quantifying drylands’ drought resistance and recovery: The importance of drought intensity, dominant life history and grazing regime. Global Chang. Biol. 2015, 21, 1258–1270. [Google Scholar] [CrossRef]
- Zeiter, M.; Schärrer, S.; Zweifel, R.; Newbery, D.M.; Stampfli, A. Timing of extreme drought modifies reproductive output in semi-natural grassland. J. Veg. Sci. 2016, 27, 238–248. [Google Scholar] [CrossRef]
- Jian, D.; Ma, Z.; Chen, L.; Duan, J.; Mitchell, D.; Zheng, Z.; Lv, M.; Zhang, H. Effects of 1.5 °C and 2 °C of warming on regional reference evapotranspiration and drying: A case study of the Yellow River Basin, China. Int. J. Climatol. 2021, 41, 791–810. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.; Stadt, K.J.; Comeau, P.G.; Zhai, L.; Dawson, A.; Alam, S.A. Drought explains variation in the radial growth of white spruce in western Canada. Agr. For. Meteorol. 2017, 233, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Remko, A.D.; Huang, G.; Renee, A.S.; Choat, B.; Anthony, P.O.G.; David, T.T. Elevated [CO2] does not ameliorate the negative effects of elevated temperature on drought-induced mortality in Eucalyptus radiata seedlings. Plant Cell Environ. 2014, 37, 1598–1613. [Google Scholar] [CrossRef]
- Depardieu, C.; Girardin, M.P.; Nadeau, S.; Lenz, P.; Bousquet, J.; Isabel, N. Adaptive genetic variation to drought in a widely distributed conifer suggests a potential for increasing forest resilience in a drying climate. New Phytol. 2020, 227, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.; Li, H.; Adnan, A.M.; Dong, L.; Sun, Y.; Hu, W.; Gong, H.; Zhao, D.; Xiong, J.; Yao, S.; et al. Quantifying Drought Resistance of Drylands in Northern China from 1982 to 2015: Regional Disparity in Drought Resistance. Forests 2022, 13, 100. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Andreas, S.; Bloor, J.M.G.; Markus, F.; Michaela, Z. High land-use intensity exacerbates shifts in grassland vegetation composition after severe experimental drought. Glob. Chang. Biol. 2018, 24, 2021–2034. [Google Scholar]
- Liu, Y.; You, C.; Zhang, Y.; Chen, S.; Zhang, Z.; Li, J.; Wu, Y. Resistance and resilience of grasslands to drought detected by SIF in inner Mongolia, China. Agr. For. Meteorol. 2021, 308, 108567. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J. High ecosystem stability of evergreen broadleaf forests under severe droughts. Global Chang. Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Pablo, G.P.; Nicolas, G.; Juan, G.; Maestre, F.T. Climate mediates the biodiversity–ecosystem stability relationship globally. Proc. Natl. Acad. Sci. USA 2018, 115, 8400–8405. [Google Scholar]
- Yang, J.; Tian, H.; Pan, S.; Chen, G.; Zhang, B.; Shree, D. Amazon drought and forest response: Largely reduced forest photosynthesis but slightly increased canopy greenness during the extreme drought of 2015/2016. Global Chang. Biol. 2018, 24, 1919–1934. [Google Scholar] [CrossRef]
- Zhang, Q.; Yao, Y.; Li, Y.; Huang, J.; Ma, Z.; Wang, Z.; Wang, S.; Wang, Y.; Zhang, Y. Progress and prospect on the study of causes and variation regularity of droughts in China. Acta Meteorol. 2020, 78, 500–521. (In Chinese) [Google Scholar]
- Yang, Q.; Li, Z.; Han, Y.; Gao, H. Responses of Baseflow to Ecological Construction and Climate Change in Different Geomorphological Types in The Middle Yellow River, China. Water 2020, 12, 304. [Google Scholar] [CrossRef] [Green Version]
- Omer, A.; Elagib, N.A.; Ma, Z.; Saleem, F.; Mohammed, A. Water scarcity in the Yellow River Basin under future climate change and human activities. Sci. Total Environ. 2020, 749, 141446. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yu, W. Prediction of Drought Characteristics in Middle Yellow River Basin Based on GM (1,1)-SVR Model. Anhui Agric. Sci. Bull. 2022, 28, 138–142. (In Chinese) [Google Scholar]
- Wei, P.; Zhu, W.; Zhao, Y.; Fang, P.; Zhang, X.; Yan, N.; Zhao, H. Extraction of Kenyan Grassland Information Using PROBA-V Based on RFE-RF Algorithm. Remote Sens. 2021, 13, 4762. [Google Scholar] [CrossRef]
- Michele, M.; Dominique, F.; Riad, B.; Mustapha, D.; Myriam, H.; Ismael, H.; Josh, H.; Mouanis, L.; Raul, L.-L.; Hamid, M.; et al. Evaluating NDVI Data Continuity Between SPOT-VEGETATION and PROBA-V Missions for Operational Yield Forecasting in North African Countries. IEEE Trans. Geosci. Remote Sens. 2016, 54, 795–804. [Google Scholar]
- Wang, W.; Guo, B.; Zhang, Y.; Zhang, L.; Ji, M.; Xu, Y.; Zhang, X.; Zhang, Y. The sensitivity of the SPEI to potential evapotranspiration and precipitation at multiple timescales on the Huang-Huai-Hai Plain, China. Theor. Appl. Climatol. 2020, 143, 87–99. [Google Scholar] [CrossRef]
- Askarimarnani, S.S.; Kiem, A.S.; Twomey, C.R. Comparing the Performance of Drought Indicators in Australia from 1900 to 2018. Int. J. Climatol. 2020, 41, 912–934. [Google Scholar] [CrossRef]
- Musei, S.K.; Nyaga, J.M.; Dubow, A.Z. SPEI-based spatial and temporal evaluation of drought in Somalia. J. Arid Environ. 2021, 184, 104296. [Google Scholar] [CrossRef]
- Xu, Q.; Jiao, Y.; Liu, C.; Liu, Z.; Ding, Y.; Zhang, H.; Tao, Y.; Zhang, Z. The spatial patterns and impact factors of stable oxygen and hydrogen isoscapes in pond water: A case study on the water-source forests of the Hani terraced fields in Yunnan, China. J. Hydrol. 2021, 603, 127097. [Google Scholar] [CrossRef]
- Yin, J.; Yuan, Z.; Li, T. The Spatial-Temporal Variation Characteristics of Natural Vegetation Drought in the Yangtze River Source Region, China. Int. J. Environ. Res. Public Health 2021, 18, 1613. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhu, X.; Wu, J.; Gu, L.; Ma, G.; Liu, X. Retrieval and analysis of coal fire temperature in Wuda coalfield, Inner Mongolia, China. Chinese Geogr. Sci. 2011, 21, 159–166. [Google Scholar] [CrossRef]
- Li, M.; Chu, R.; Sha, X.; Xie, P.; Ni, F.; Wang, C.; Jiang, Y.; Shen, S.; Towfiqul, I.A.R.M. Monitoring 2019 Drought and Assessing Its Effects on Vegetation Using Solar-Induced Chlorophyll Fluorescence and Vegetation Indexes in the Middle and Lower Reaches of Yangtze River, China. Remote Sens. 2022, 14, 2569. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Chen, A.; Gao, M.; Slette, I.J.; Piao, S. The impact of the 2009/2010 drought on vegetation growth and terrestrial carbon balance in Southwest China. Agr. For. Meteorol. 2019, 269, 239–248. [Google Scholar] [CrossRef]
- Pi, G.; He, Z.; Zhang, L.; Yang, M.; You, M. Response of Vegetation to Meteorological Drought in Watershed at different Time Scales—A Case Study of Guizhou Province. Res. Soil Water Conserv. 2022, 29, 277–284. (In Chinese) [Google Scholar]
- Ding, Y.; Li, Z.; Peng, S. Global analysis of time-lag and -accumulation effects of climate on vegetation growth. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Wei, X.; He, W.; Zhou, Y.; Ju, W.; Xiao, J.; Li, X.; Liu, Y.; Xu, S.; Bi, W.; Zhang, X.; et al. Global assessment of lagged and cumulative effects of drought on grassland gross primary production. Ecol. Indic. 2022, 136, 108646. [Google Scholar] [CrossRef]
- Gu, X.; Guo, E.; Yin, S.; Wang, Y.; Na, R.; Wan, Z. Assessment of the Cumulative and Lagging Effects of Drought on Vegetation Growth in Inner Mongolia. Acta Agrestia Sin. 2021, 29, 1301–1310. (In Chinese) [Google Scholar]
- Zhao, A.; Yu, Q.; Feng, L.; Zhang, A.; Pei, T. Evaluating the cumulative and time-lag effects of drought on grassland vegetation: A case study in the Chinese Loess Plateau. J. Environ. Manag. 2020, 261, 110214. [Google Scholar] [CrossRef]
- Brendan, C.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.H.; Evans, J.P.; Mccabe, M.F. The Impact of Observed Vegetation Changes on Land—Atmosphere Feedbacks During Drought. J. Hydrometeorol. 2014, 15, 759–776. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Impacts of vegetation on drought trends. Chinese J. Atmos. Sci. 2016, 40, 142–156. (In Chinese) [Google Scholar]
- Jones, J.A.; Creed, I.F.; Hatcher, K.L.; Warren, R.J.; Adams, M.B.; Benson, M.H.; Boose, E.; Brown, W.A.; Campbell, J.L.; Covich, A.; et al. Ecosystem Processes and Human Influences Regulate Streamflow Response to Climate Change at Long-Term Ecological Research Sites. BioScience 2012, 62, 390–404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Guo, L.; Liang, C.; Zhao, L.; Wang, J.; Zhan, C.; Jiang, S. Encounter risk analysis of crop water requirements and effective precipitation based on the copula method in the Hilly Area of Southwest China. Agric. Water Manag. 2022, 266, 107571. [Google Scholar] [CrossRef]
- Huang, M.; Wang, X.; Keenan, T.F.; Piao, S. Drought timing influences the legacy of tree growth recovery. Global Chang. Biol. 2018, 24, 3546–3559. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Global Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [Green Version]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef]
- Stuart-Haëntjens, E.; de Boeck, H.J.; Lemoine, N.P.; Mänd, P.; Kröel-Dulay, G.; Schmidt, I.K.; Jentsch, A.; Stampfli, A.; Anderegg, W.R.L.; Bahn, M.; et al. Mean annual precipitation predicts primary production resistance and resilience to extreme drought. Sci. Total Environ. 2018, 636, 360–366. [Google Scholar] [CrossRef]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, M.; Ma, Z.; Yang, Q.; LV, M.; Clark, R. Assessment of an Evapotranspiration Deficit Drought Index in Relation to Impacts on Ecosystems. Adv. Atmos. Sci. 2019, 36, 1273–1287. [Google Scholar] [CrossRef]
- Srinidhi, J.; Jew, D.; Ashutosh, S.; Budhaditya, H.; Manish, K.G. Probabilistic evaluation of vegetation drought likelihood and its implications to resilience across India. Global Planet. Chang. 2019, 176, 23–35. [Google Scholar]
- Corey, L.; Pedram, R.; Navin, R. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Liu, J.; Shen, L.; Wang, Z.; Duan, S.; Wu, W.; Peng, X.; Wu, C.; Jiang, Y. Response of plants water uptake patterns to tunnels excavation based on stable isotopes in a karst trough valley. J. Hydrol. 2019, 571, 485–493. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Zhou, S.; Ciais, P.; McCarthy, H.; Luo, Y. Canopy and physiological controls of GPP during drought and heat wave. Geophys. Res. Lett. 2016, 43, 3325–3333. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Range | Resistance Level | Resilience Range | Resilience Level |
|---|---|---|---|
| <10 | less resistance | <0.8 | less resilience |
| 10–14 | fair resistance | 0.8–1.2 | fair resilience |
| >14 | more resistance | >1.2 | more resilience |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Chen, F.; Ding, H.; Li, Y.; Shi, M. Quantifying Vegetation Stability under Drought in the Middle Reaches of Yellow River Basin, China. Forests 2022, 13, 1138. https://doi.org/10.3390/f13071138
Shi X, Chen F, Ding H, Li Y, Shi M. Quantifying Vegetation Stability under Drought in the Middle Reaches of Yellow River Basin, China. Forests. 2022; 13(7):1138. https://doi.org/10.3390/f13071138
Chicago/Turabian StyleShi, Xiaoliang, Fei Chen, Hao Ding, Yi Li, and Mengqi Shi. 2022. "Quantifying Vegetation Stability under Drought in the Middle Reaches of Yellow River Basin, China" Forests 13, no. 7: 1138. https://doi.org/10.3390/f13071138