
Cold Tolerance in Pinewood Nematode Bursaphelenchus xylophilus Promoted Multiple Invasion Events in Mid-Temperate Zone of China


Abstract
:1. Introduction
2. Materials and Methods
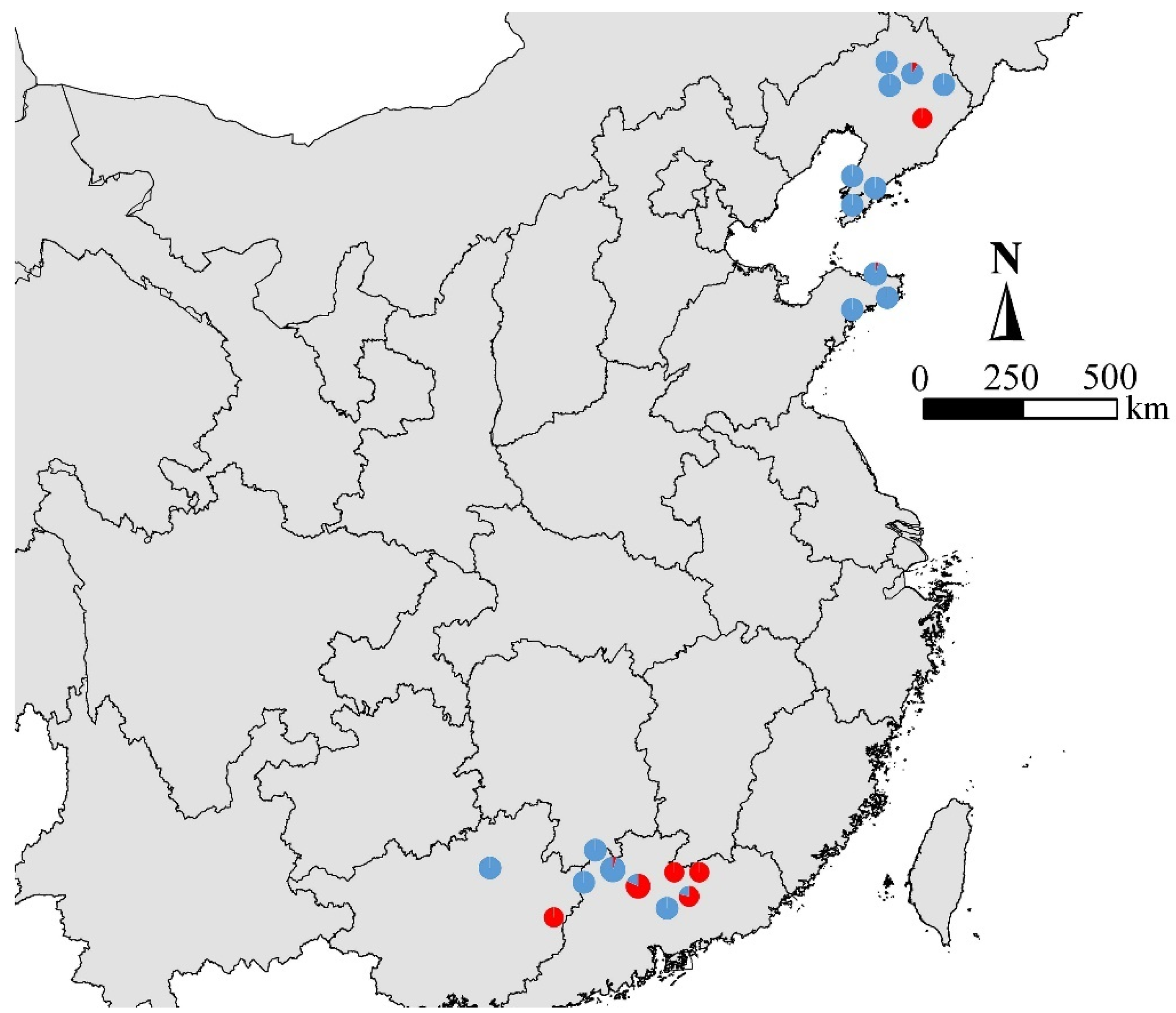
2.1. Nematode Collection
2.2. Test of Survival under Low Temperatures
2.3. Test of Survival under Gradient Cooling
2.4. Statistical Analysis
2.5. Whole Genome Sequencing and Quality Control
2.6. Variant Identification and Annotation
2.7. Phylogenetic and Population Structure Analyses
3. Results
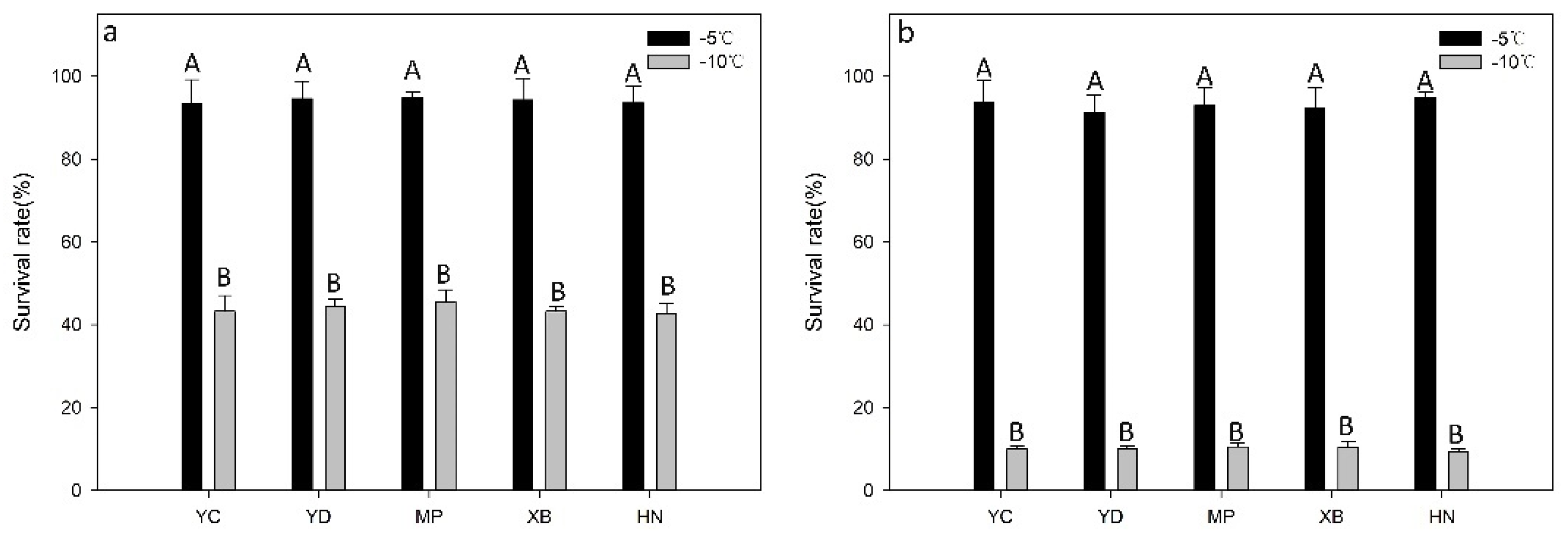
3.1. Effect of Low Temperatures on the Survival Rates of Different Nematode Isolates
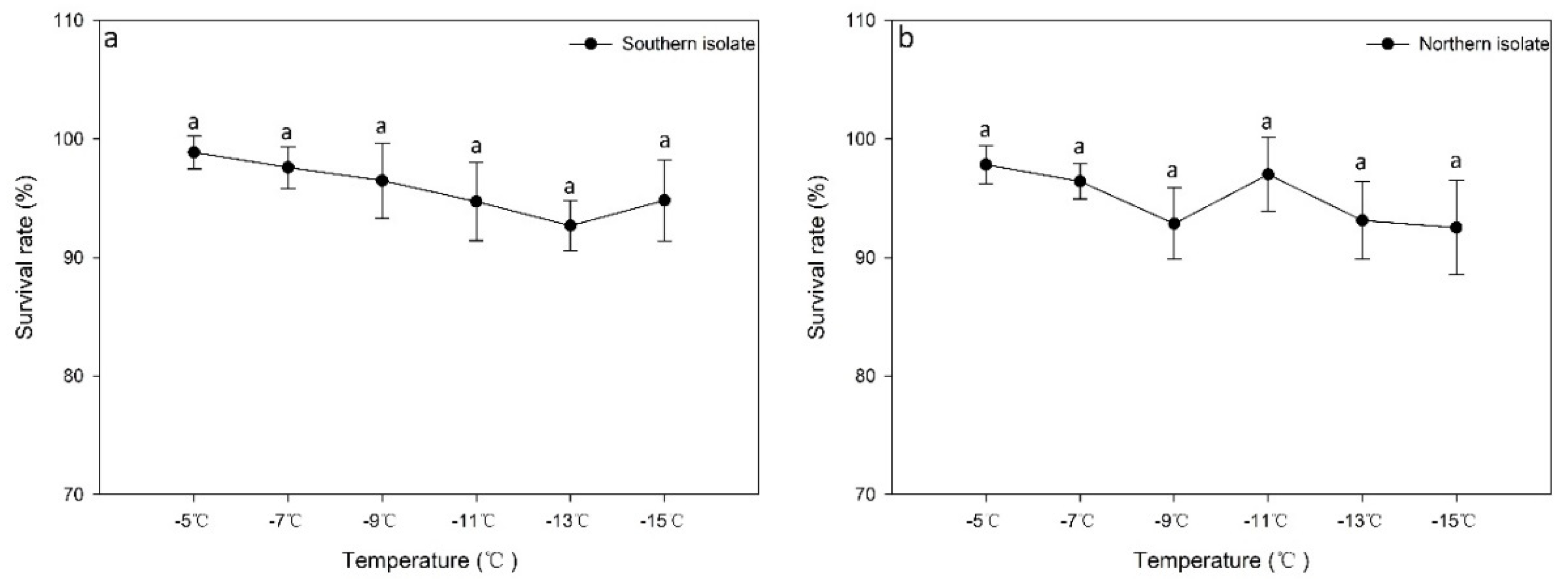
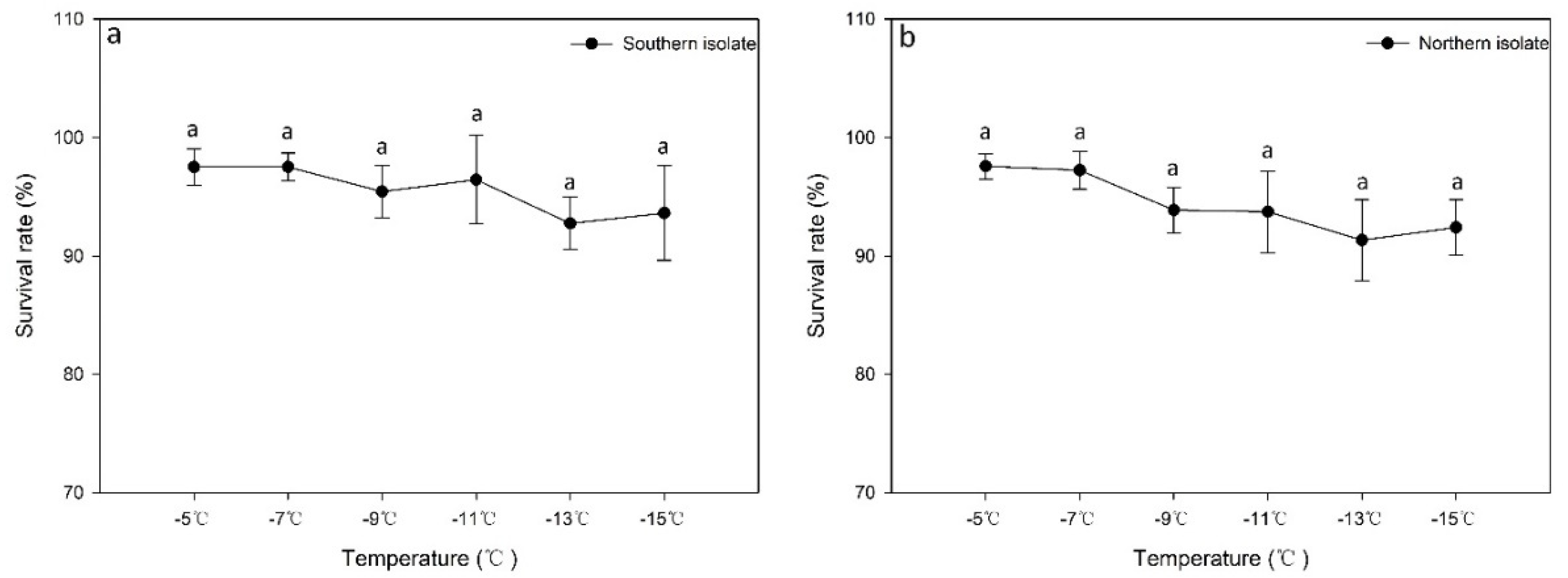
3.2. Effect of Gradient Cooling on the Survival Rate and Activity of Different Nematode Isolates
3.3. Summary of Whole Genome Resequencing Data and Variant Calling
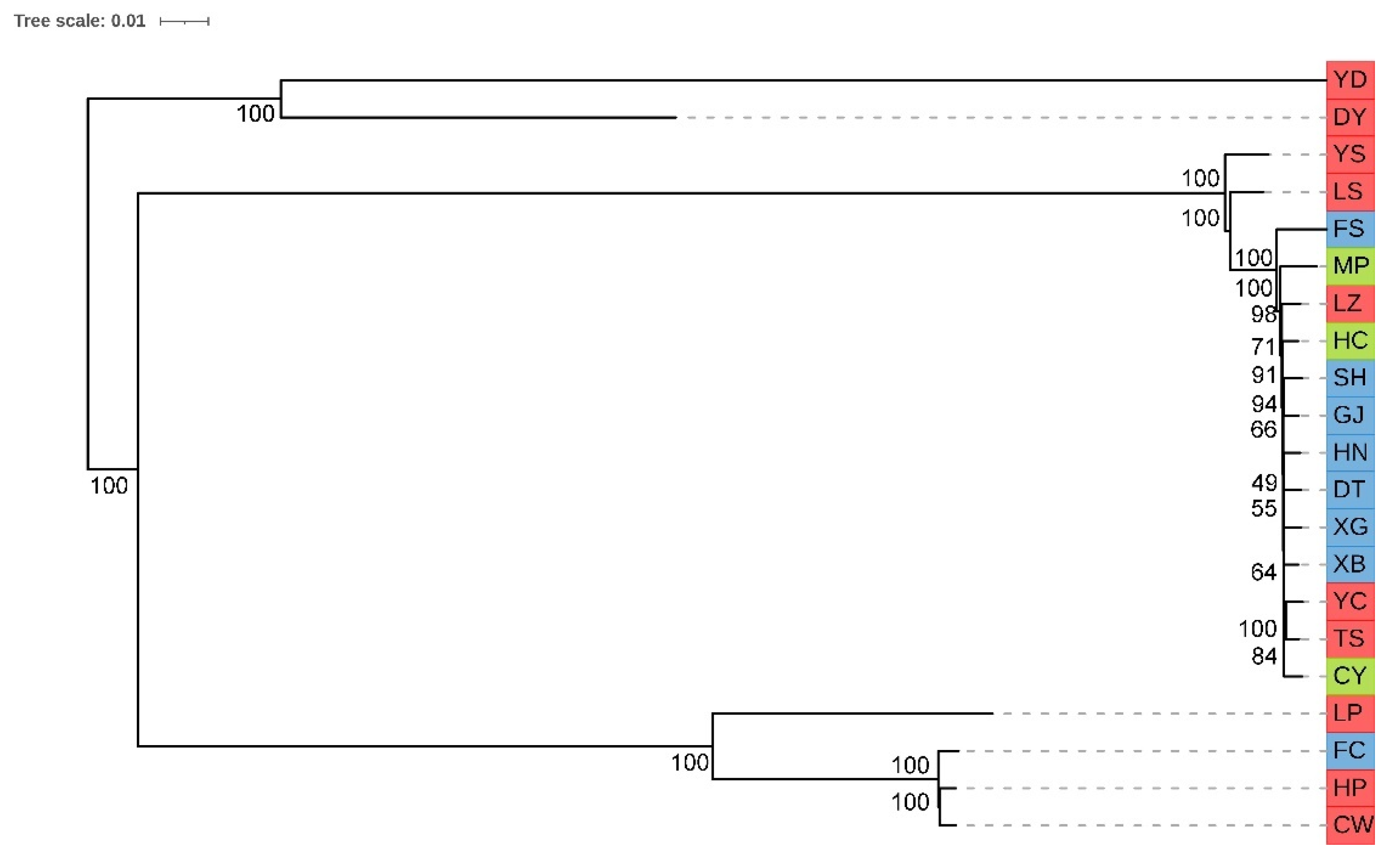
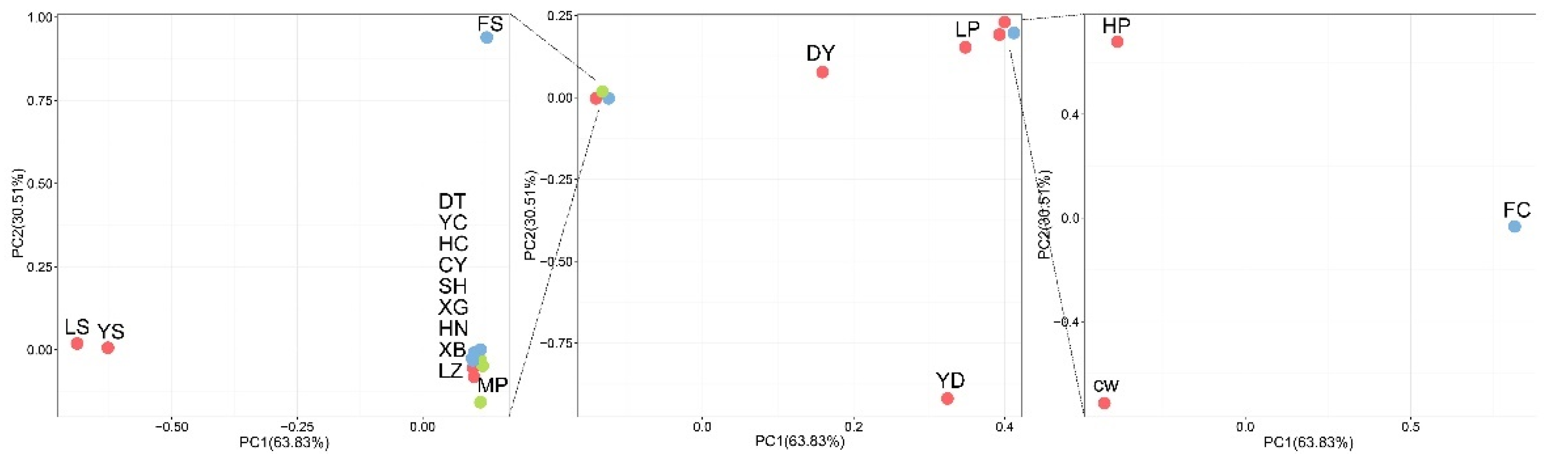
3.4. Phylogenetic Analysis and Principal Component Analysis of 21 Pinewood Nematode Isolates
3.5. Population Structure of 21 Pinewood Nematode Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [Green Version]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar] [PubMed]
- Ye, J. Epidemic status of pine wilt disease in China and its prevention and control techiques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. [Google Scholar]
- Sun, H.; Zhou, Y.; Li, X.; Zhang, Y.; Wang, Y. Occurrence of major forestry pests in China in 2020 and prediction of occurrence trend in 2021. For. Pest Dis. 2021, 40, 45–48. [Google Scholar]
- Yu, H.; Wu, H.; Zhang, X.; Wang, L.; Zhan, X.; Song, Y. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. [Google Scholar]
- Pan, L.; Li, Y.; Liu, Z.; Meng, F.; Chen, J.; Zhang, X. Isolation and identification of pine wood nematode in Pinus koraiensis in Fengcheng, Liaoning Province. For. Pest Dis. 2019, 38, 1–4. [Google Scholar]
- Yu, H.; Wu, H.; Huang, R.; Wang, J.; Zhang, R.; Song, Y. Separation and identification of Bursaphelenchus xylophilus from Pinus sylvestris var. mongolica in Fushun city. For. Pest Dis. 2020, 39, 6–10. [Google Scholar]
- Ishibashi, N.; Kondo, E. Occurrence and survival of the dispersal forms of pine wood nematode, Bursaphelenchus lignicolus. Appl. Entomol. Zool. 1977, 12, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, Y.; Pan, L.; Meng, F.; Zhang, X. Cold adaptive potential of pine wood nematodes overwintering in plant hosts. Biol. Open 2019, 8, bio041616. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Liu, W.; Wang, F.; Ma, L.; Li, D.; Wang, B.; Hao, X. Fat accumulation in Bursaphelenchus xylophilus by positively regulating Bx-SCD under low temperature. J. Northeast For. Univ. 2017, 45, 89–93. [Google Scholar]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F.; Jiang, S.; Wang, J. Trehalose in pine wood nematode participates in DJ3 formation and confers resistance to low-temperature stress. BMC Genom. 2021, 22, 524. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F. Genetic characteristics of Bursaphelenchus xylophilus third-stage dispersal juveniles. Sci. Rep. UK 2021, 11, 3908. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wei, W.; Kulhavy, D.L.; Zhang, X.Y.; Sun, J. Low temperature induces two growth-arrested stages and change of secondary metabolites in Bursaphelenchus xylophilus. Nematology 2007, 9, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Cui, R.; Li, Y.; Feng, Y.; Zhang, X. Investigation of pinewood nematodes in Pinus tabuliformis Carr. under low-temperature conditions in Fushun, China. Forests 2020, 11, 993. [Google Scholar] [CrossRef]
- Pan, L.; Cui, R.; Li, Y.; Zhang, W.; Bai, J.; Li, J.; Zhang, X. Third-stage dispersal juveniles of Bursaphelenchus xylophilus can resist low-temperature stress by entering cryptobiosis. Biology 2021, 10, 785. [Google Scholar] [CrossRef]
- Kong, Q.; Ding, X.; Chen, Y.; Ye, J. Comparison of morphological indexes and the pathogenicity of Bursaphelenchus xylophilus in northern and southern China. Forests 2021, 12, 310. [Google Scholar] [CrossRef]
- Pimentel, C.S.; Ayres, M.P. Latitudinal patterns in temperature-dependent growth rates of a forest pathogen. J. Therm. Biol. 2018, 72, 39–43. [Google Scholar] [CrossRef]
- Zheng, J.; Bian, J.; Ge, Q.; Zhixin, H.; Yin, Y.; Liao, Y. The climate regionalization in China for 1981–2010. Chin. Sci. Bull. 2013, 50, 3088–3099. (In Chinese) [Google Scholar]
- Cheng, X.; Xie, P.; Cheng, F.; Xu, R.; Xie, B. Competitive displacement of the native species Bursaphelenchus mucronatus by an alien species Bursaphelenchus xylophilus (Nematoda: Aphelenchida: Aphelenchoididae): A case of successful invasion. Biol. Invasions 2008, 11, 205–213. [Google Scholar] [CrossRef]
- Wang, Y.; Yamada, T.; Sakaue, D.; Suzuki, K. Variations in life history parameters and their influence on rate of population increase of different pathogenic isolates of the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2005, 7, 459–467. [Google Scholar] [CrossRef]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar] [PubMed]
- Liu, H.B.; Rui, L.; Feng, Y.Q.; Wu, X.Q. Autophagy contributes to resistance to the oxidative stress induced by pine reactive oxygen species metabolism, promoting infection by Bursaphelenchus xylophilus. Pest Manag. Sci. 2020, 76, 2755–2767. [Google Scholar] [CrossRef] [PubMed]
- Wharton, D.A.; Raymond, M.R. Cold tolerance of the Antarctic nematodes Plectus murrayi and Scottnema lindsayae. J. Comp. Physiol. B 2015, 185, 281–289. [Google Scholar] [CrossRef]
- Hayashi, M.; Wharton, D.A. The oatmeal nematode Panagrellus redivivus survives moderately low temperatures by freezing tolerance and cryoprotective dehydration. J. Comp. Physiol. B 2011, 181, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Wharton, D.A.; Marshall, C.J. Cold tolerance of an Antarctic nematode that survives intracellular freezing: Comparisons with other nematode species. J. Comp. Physiol. B 2008, 178, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.R.; Wharton, D.A. The ability of the Antarctic nematode Panagrolaimus davidi to survive intracellular freezing is dependent upon nutritional status. J. Comp. Physiol. B 2013, 183, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Hardy, O.J.; Vekemans, X. Spagedi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Loiselle, B.A.; Sork, V.L.; Nason, J.; Graham, C. Spatial genetic structure of a tropical understory shrub, Psychotria officinalis (Rubiaceae). Am. J. Bot. 1995, 82, 1420–1425. [Google Scholar] [CrossRef]
- Dayi, M.; Sun, S.; Maeda, Y.; Tanaka, R.; Yoshida, A.; Tsai, I.J.; Kikuchi, T.; Rokas, A. Nearly complete genome assembly of the pinewood nematode Bursaphelenchus xylophilus strain Ka4C1. Microbiol. Resour. Announc. 2020, 9, e01002-20. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [Green Version]
- Jikumaru, S.; Togashi, K. Resistance of an indigenous biological system against expansion of the invasive nematode, Bursaphelenchus xylophilus, in cool areas of Japan. Nematol. Int. J. Fundam. Appl. Nematol. Res. 2008, 10, 679–687. [Google Scholar] [CrossRef]
- Wang, F. Pine wilt in meteorological hazard environments. Nat. Hazards 2014, 72, 723–741. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X. Analysis on the trend of invasion and expansion of Bursaphelenchus xylophilus. For. Pest Dis. 2018, 37, 1–4. [Google Scholar]
- Huang, J.; Xi, X.; Ding, X.; Ye, J. Study on the population differentiation of Bursaphelenchus xylophilus in Guangdong Province by SNP markers. J. Nanjing For. Univ. Nat. Sci. Ed. 2019, 43, 25–31. [Google Scholar]
- Wang, Q.; Ding, X.; Ye, J.; Shi, X. Study on the genetic differentiation of Bursaphelenchus xylophilus in East China based on SNP markers. J. Nanjing For. Univ. Nat. Sci. Ed. 2021, 1, 1–10. [Google Scholar]
- Ding, X.; Guo, Y.; Ye, J.; Wu, X.; Lin, S.; Chen, F.; Zhu, L.; Huang, L.; Song, X.; Zhang, Y.; et al. Population differentiation and epidemic tracking of Bursaphelenchus xylophilus in China based on chromosome—Level assembly and whole—Genome sequencing data. Pest Manag. Sci. 2022, 78, 1213–1226. [Google Scholar] [CrossRef]
- Rutherford, T.A.; Webster, J.M. Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Ohsawa, M.; Akiba, M. Possible altitude and temperature limits on pine wilt disease: The reproduction of vector sawyer beetles (Monochamus alternatus), survival of causal nematode (Bursaphelenchus xylophilus), and occurrence of damage caused by the disease. Eur. J. For. Res. 2014, 133, 225–233. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A.; Pan, H.; Fang, G.; Ye, J.; Zhang, Y.; Sun, J. Role of human-mediated dispersal in the spread of the pinewood nematode in China. PLoS ONE 2009, 4, e4646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, J.; Simões, M.J.; Gomes, P.; Barroso, C.; Pinho, D.; Conceição, L.; Fonseca, L.; Abrantes, I.; Pinheiro, M.; Egas, C.; et al. Assessment of the geographic origins of pinewood nematode isolates via single nucleotide polymorphism in effector genes. PLoS ONE 2013, 8, e83542. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Locality | Host | Annual Average Temperature 1 |
|---|---|---|---|
| DT | Dengta, Liaoning | Pinus rigida | 2–8 °C |
| FC | Fengcheng, Liaoning | P. koraiensis | 2–8 °C |
| XB | Xinbing, Liaoning | P. koraiensis | 2–8 °C |
| FS | Fushun, Liaoning | P. tabulaeformis | 2–8 °C |
| GJ | Ganjingzi, Liaoning | P. tabulaeformis | 2–8 °C |
| HN | Hunnan, Liaoning | P. tabulaeformis | 2–8 °C |
| SH | Shahekou, Liaoning | P. thunbergii | 2–8 °C |
| XG | Xigang, Liaoning | P. thunbergii | 2–8 °C |
| CY | Chengyang, Shandong | P. thunbergii | 10–15 °C |
| HC | Huancui, Shandong | P. thunbergii | 10–15 °C |
| MP | Mouping, Shandong | P. thunbergii | 10–15 °C |
| CW | Cangwu, Guangxi | P. massoniana | 19–24 °C |
| TS | Liuzhou, Guangxi | P. massoniana | 19–24 °C |
| LS | Lianshan, Guangdong | P. massoniana | 19–24 °C |
| YD | Yingde, Guangdong | P. massoniana | 19–24 °C |
| DY | Dongyuan, Guangdong | P. massoniana | 19–24 °C |
| HP | Heping, Guangdong | P. massoniana | 19–24 °C |
| LP | Lianping, Guangdong | P. massoniana | 19–24 °C |
| YS | Yangshan, Guangdong | P. massoniana | 19–24 °C |
| LZ | Lianzhou, Guangdong | P. massoniana | 19–24 °C |
| YC | Yuancheng, Guangdong | P. massoniana | 19–24 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Tao, J.; Zong, S. Cold Tolerance in Pinewood Nematode Bursaphelenchus xylophilus Promoted Multiple Invasion Events in Mid-Temperate Zone of China. Forests 2022, 13, 1100. https://doi.org/10.3390/f13071100
Li Z, Tao J, Zong S. Cold Tolerance in Pinewood Nematode Bursaphelenchus xylophilus Promoted Multiple Invasion Events in Mid-Temperate Zone of China. Forests. 2022; 13(7):1100. https://doi.org/10.3390/f13071100
Chicago/Turabian StyleLi, Zhenxiao, Jing Tao, and Shixiang Zong. 2022. "Cold Tolerance in Pinewood Nematode Bursaphelenchus xylophilus Promoted Multiple Invasion Events in Mid-Temperate Zone of China" Forests 13, no. 7: 1100. https://doi.org/10.3390/f13071100