
Potential European Geographical Distribution of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Scolytinae) under Current and Future Climate Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Modelling Species Distribution
3. Results
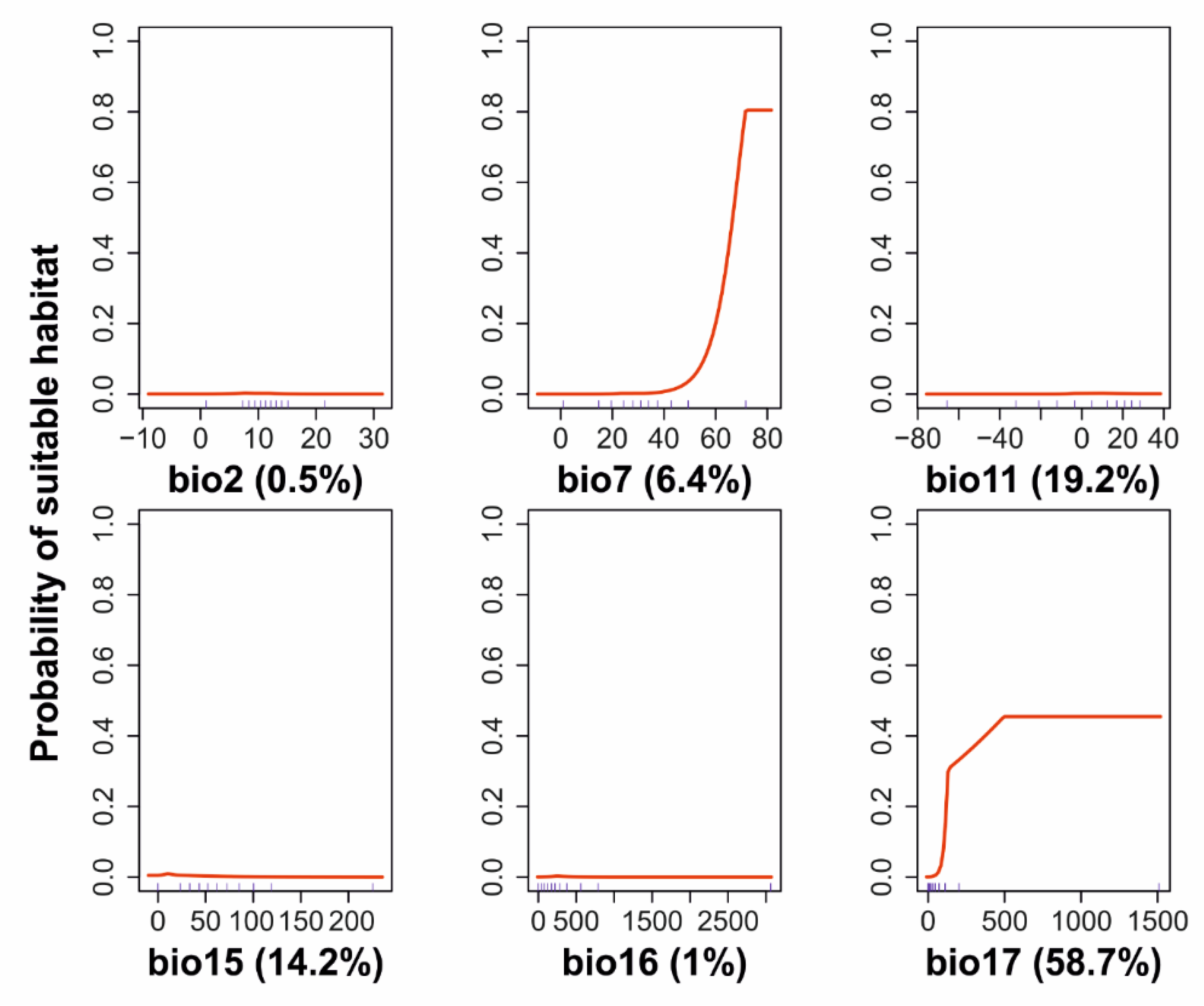
3.1. G. materiarius Distribution Model
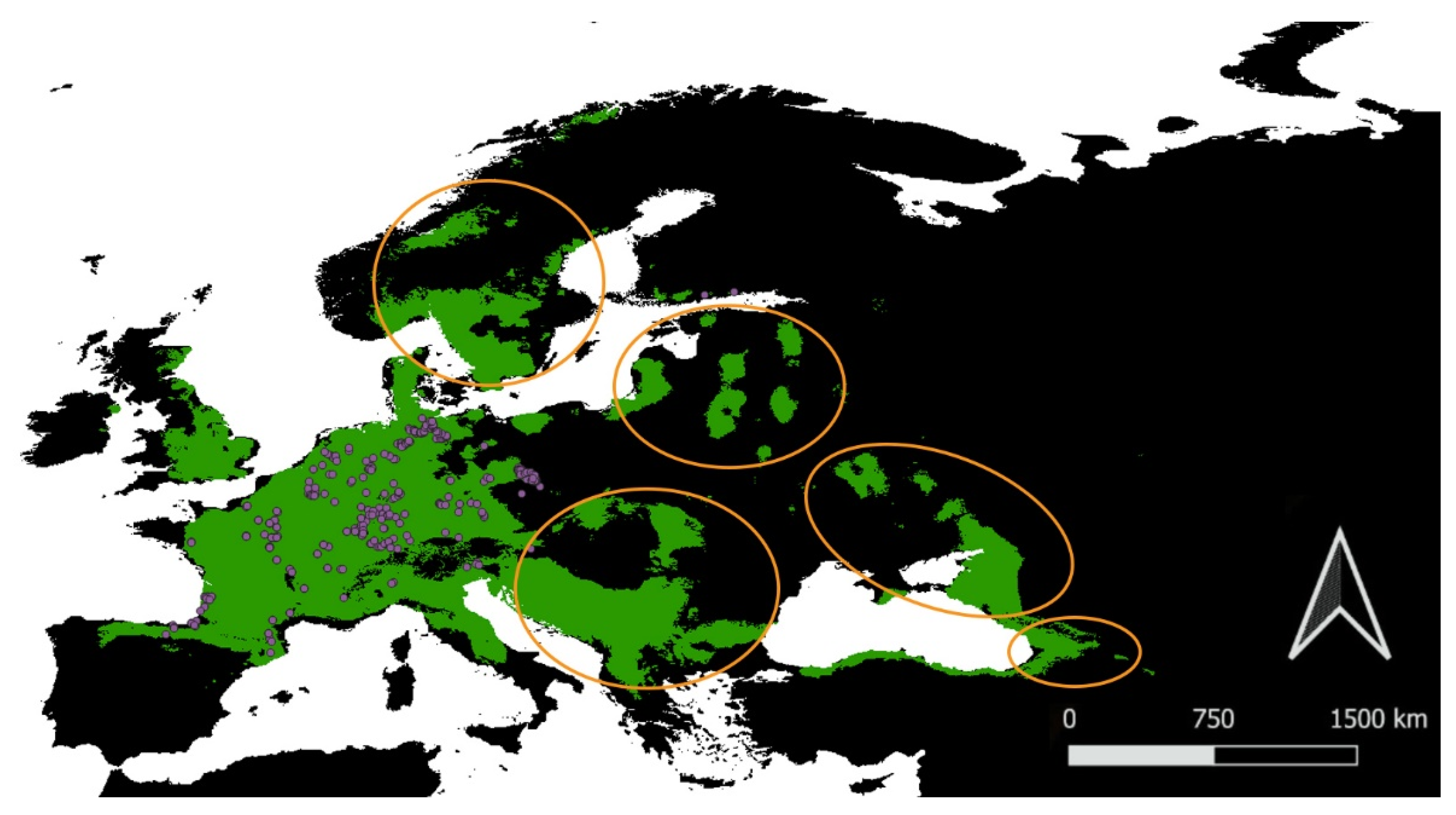
3.2. Current Potential Distribution
3.3. Predicted Range Shifts
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perrings:, C. Mitigation and adaptation strategies for the control of biological invasions. Ecol. Econ. 2005, 52, 315–325. [Google Scholar] [CrossRef]
- Lenzner, B.; Leclère, D.; Franklin, O.; Seebens, H.; Roura-Pascual, N.; Obersteiner, M.; Dullinger, S.; Essl, F. A framework for global twenty-first century scenarios and models of biological invasions. BioScience 2019, 69, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Brondizio, E.S.; Settele, J.; Díaz, S.; Ngo, H.T. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019; pp. 1–1148. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Brockerhoff, E.G.; Kalisz, S.; Nuñez, M.A.; Wardle, D.A.; Wingfield, M.J. Biological invasions in forest ecosystems. Biol. Invasions 2017, 19, 3437–3458. [Google Scholar] [CrossRef]
- Zenni, R.D.; Essl, F.; García-Berthou, E.; McDermott, S.M. The economic costs of biological invasions around the world. NeoBiota 2021, 67, 1–9. [Google Scholar] [CrossRef]
- Kirkendall, L.R.; Faccoli, M. Bark beetles and pinhole borers (Curculionidae, Scolytinae, Platypodinae) alien to Europe. ZooKeys 2010, 56, 227–251. [Google Scholar] [CrossRef] [Green Version]
- Faccoli, M.; Gallego, D.; Branco, M.; Brockerhoff, E.G.; Corley, J.; Coyle, D.R.; Hurley, B.P.; Jactel, H.; Lakatos, F.; Lantschner, V.; et al. A first worldwide multispecies survey of invasive Mediterranean pine bark beetles (Coleoptera: Curculionidae, Scolytinae). Biol. Invasions 2020, 22, 1785–1799. [Google Scholar] [CrossRef]
- Grégoire, J.-C.; Raffa, K.F.; Lindgren, B.S. Economics and politics of bark beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 585–613. [Google Scholar] [CrossRef]
- Kirkendall, L.R. Invasive bark beetles (Coleoptera, Curculionidae, Scolytinae) in Chile and Argentina, including two species new for South America, and the correct identity of the Orthotomicus species in Chile and Argentina. Diversity 2018, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Barnouin, T.; Soldati, F.; Roques, A.; Faccoli, M.; Kirkendall, L.R.; Mouttet, R.; Daubree, J.B.; Noblecourt, T. Bark beetles and pinhole borers recently or newly introduced to France (Coleoptera: Curculionidae, Scolytinae and Platypodinae). Zootaxa 2020, 4877, 51–74. [Google Scholar] [CrossRef]
- Inward, D.J.G. Three new species of ambrosia beetles established in Great Britain illustrate unresolved risks from imported wood. J. Pest Sci. 2020, 93, 117–126. [Google Scholar] [CrossRef]
- Hulme, P.E. Invasion pathways at a crossroad: Policy and research challenges for managing alien species introductions. J. Appl. Ecol. 2015, 52, 1418–1424. [Google Scholar] [CrossRef]
- Sauvard, D.; Branco, M.; Lakatos, F.; Faccoli, M.; Kirkendall, L. Weevils and Bark Beetles (Coleoptera, Curculionoidea). Chapter 8.2. BioRisk 2010, 4, 219–266. [Google Scholar] [CrossRef]
- Knížek, M.; Beaver, R. Taxonomy and systematics of bark and ambrosia beetles. In Bark and Wood Boring Insects in Living Trees in Europe, A Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J., Evans, H.F., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 41–54. [Google Scholar]
- Beaver, R.A.; Wilding, N.; Collins, N.; Hammond, P.; Webber, J. Insect-fungus relationships in the bark and ambrosia beetles. In Insect-Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 121–143. [Google Scholar]
- Hulcr, J.; Mogia, M.; Isua, B.; Novotny, V. Host specificity of ambrosia and bark beetles (Col.; Curculionidae: Scolytinae and Platypodinae) in a New Guinea rainforest. Ecol. Entomol. 2007, 32, 762–772. [Google Scholar] [CrossRef]
- Gomez, D.F.; Lin, W.; Gao, L.; Li, Y. New host plant records for the Euwallacea fornicatus (Eichhoff) species complex (Coleoptera: Curculionidae: Scolytinae) across its natural and introduced distribution. J. Asia-Pac. Entomol. 2019, 22, 338–340. [Google Scholar] [CrossRef]
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Reding, M.E. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, 97–166. [Google Scholar] [CrossRef]
- Tisserat, N.; Cranshaw, W.; Putnam, M.L.; Pscheidt, J.; Leslie, C.A.; Murray, M.; Hoffman, J.; Barkley, Y.; Alexander, K.; Seybold, S.J. Thousand cankers disease is widespread in black walnut in the western United States. Plant Health Prog. 2011, 12, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, D.; Duncan, R.E.; Ploetz, J.N.; Campbell, A.F.; Ploetz, R.C.; Peña, J.E. Lateral transfer of a phytopathogenic symbiont among native and exotic ambrosia beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; De Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Geiser, D.M. An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on avocado and other plant hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef]
- Vannini, A.; Contarini, M.; Faccoli, M.; Valle, M.D.; Rodriguez, C.M.; Mazzetto, T.; Guarneri, D.; Vettraino, A.M.; Speranza, S. First report of the ambrosia beetle Xylosandrus compactus and associated fungi in the Mediterranean maquis in Italy, and new host–pest associations. EPPO Bull. 2017, 47, 100–103. [Google Scholar] [CrossRef]
- Hirschheydt, J. Der amerikanische Nutzholz-borkenkäfer Gnathotrichus materiarius (Fitch) hat die Schweiz erreicht. Entomol. Gesellsch. 1992, 65, 33–37. [Google Scholar]
- Mazur, A.; Witkowski, R.; Góral, J.; Rogowski, G. Occurrence of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera, Curculionidae, Scolytinae) in South-Western Poland. Folia For. Pol. 2018, 60, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Balachowsky, A. Faune de France, Volume 50: Coléoptères scolytides; Lechevalier: Paris, France, 1949; pp. 1–320. [Google Scholar]
- Knížek, M. Scolytinae. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2011; Curculionoidea I; pp. 204–251. [Google Scholar]
- Aurenhammer, S.; von Komposch, C.; Holzer, E.; Holzschuh, C.; Holzinger, W.E. Xylobionte Käfergemeinschaften (Insecta: Coleoptera) im Bergsturzgebiet des Dobratsch (Schütt, Kärnten). Carinth. II 2015, 205, 439–502. [Google Scholar]
- Moucheron, B.; Warzee, N. Gnathotrichus materiarius (Fitch, 1858): Un scolyte nordamericain a surveiller, nouveau pour la faune Belge (Coleoptera, Scolytidae). Lambillionea 2006, 106, 610–612. [Google Scholar]
- Knížek, M. Faunistic records from the Czech Republic—272. Coleoptera: Curculionidae: Scolytinae. Klapalekiana 2009, 45, 22. [Google Scholar]
- Valkama, H.; Martikainen, P.; Räty, M. First record of North American ambrosia beetle Gnathotrichus materiarius (Fitch) (Coleoptera, Scolytidae) in Finland—A new potential forest pest? Entomol. Fenn. 1997, 8, 193–195. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Iturrondobeitia, J.C.; Goldarazena, A.A. Primera cita de la Península Ibérica de Gnathotrichus materiarius (Fitch, 1858) y Xylosandrus germanus (Blandford, 1894) (Coleoptera: Scolytinae). Boletín de La SEA 2007, 40, 527–532. [Google Scholar]
- Doom, D. Notes on Gnathotrichus materiarius (Col. Scolytidae), a timber beetle new to the Netherlands. Entomol. Ber. 1967, 27, 143–148. [Google Scholar]
- Schedl, K.E. Ein für Deutschland und Holland neuer Borkenkäfer. Anzeig. Schädling. 1966, 39, 118–120. [Google Scholar] [CrossRef]
- Witkowski, R.; Góral, J.; Nowik, K.; Rogowski, G.; Skałecka, K.; Mazur, A. Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Curculionidae, Scolytinae) new species of beetle in the Polish fauna. Acta Sci. Pol. Silv. Colendar. Ratio Ind. Lignar. 2016, 15, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Jurc, M.; Bojovic, S.; Fernández, M.F.; Jurc, D. The attraction of cerambycids and other xylophagous beetles, potential vectors of Bursaphelenchus xylophilus, to semiochemicals in Slovenia. Phytoparasitica 2012, 40, 337–349. [Google Scholar] [CrossRef]
- Gillerfors, G. Skalbaggar införda till Sverige med importerad massaved. Entomol. Tidskr. 1988, 9, 42–45. [Google Scholar]
- Faccoli, M. The North American Gnathotricus materiarius (Fitch) (Coleoptera Scolytidae): An ambrosia beetle new to Italy. Redia 1998, 81, 151–154. [Google Scholar]
- Brockerhoff, E.G.; Bain, J.; Kimberley, M.; Knížek, M. Interception frequency of exotic bark and ambrosia beetles (Coleoptera: Scolytinae) and relationship with establishment in New Zealand and worldwide. Can. J. For. Res. 2006, 36, 289–298. [Google Scholar] [CrossRef]
- Kirkendall, L.R. The evolution of mating systems in bark and ambrosia beetles (Coleoptera: Scolytidae and Platypodidae). Zool. J. Linn. Soc. 1983, 77, 293–352. [Google Scholar] [CrossRef] [Green Version]
- Flechtmann, C.A.H.; Berisford, C.W. Identification of sulcatol, a potential pheromone of the ambrosia beetle Gnathotrichus materiarius (Col.; Scolytidae). J. Appl. Entomol. 2003, 127, 189–194. [Google Scholar] [CrossRef]
- Jordal, B.H.; Cognato, A.I. Molecular phylogeny of bark and ambrosia beetles reveals multiple origins of fungus farming during periods of global warming. BMC Ecol. Evol. 2012, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Batra, L.R. Contributions to our knowledge of ambrosia fungi II. Endomycopsis fasciculate nom. nov. Am. J. Bot. 1963, 50, 481–487. [Google Scholar] [CrossRef]
- Batra, L.R. Habitat and Nutrition of Dipodascus and Cephaloascus. Mycologia 1963, 55, 508–520. [Google Scholar] [CrossRef]
- Postner, M. Scolytidae, Borkenkäfer. In Die Forstschädlinge Europas: Ein Handbuch in 5 Bänden; Band 2: Käfer; Schwenke, W., Ed.; Verlag Paul Parey: Berlin/Heidelberg, Germany, 1974; pp. 334–482. [Google Scholar]
- Kirschner, R. Diversität mit Borkenkäfern Assoziierter Filamentser Mikropilze. Fakultät für Biologie. Ph.D. Thesis, Eberhard-Karls Universität, Tübingen, Germany, 1998. [Google Scholar]
- Bleich, O.; Gürlich, S.; Köhler, F. Verzeichnis und Verbreitungsatlas der Käfer Deutschlands. Available online: http://www.coleoweb.de/ (accessed on 19 June 2019).
- Dengler, K. Bemerkenswerte Baumschädlinge vor und in der Stadt Rottenburg. Allg. Forst 1990, 45, 361–365. [Google Scholar]
- Lindgren, B.S.; Fraser, R.G. Control of ambrosia beetle damage by mass trapping at a dryland log sorting area in British Columbia. For. Chron. 1994, 70, 159–163. [Google Scholar] [CrossRef]
- Raffa, K.F.; Gregoire, J.C.; Lindgren, B.S. Natural history and ecology of bark beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 1–40. [Google Scholar] [CrossRef]
- Shore, T.L. Ambrosia beetles. In Forest Pest Leaflet; Forestry Canada: Burnaby, BC, Canada; BC Ministry of Forests: Victoria, BC, Canada, 1998; Volume 72, pp. 1–4. [Google Scholar]
- Daterman, G.E.; Overhulser, D.L. Forest Insect Disease Leaflet; USDA Forest Service: Corvallis, OR, USA, 2002; pp. 1–8.
- IPCC. Summary for Policymakers in Climate Change 2013: The Physical Science Basis, Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press & Assessment: Cambridge, UK; UKNY: Westbury, NY, USA, 2013. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; Garcia, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffen, W.; Broadgate, W.; Deutsch, L.; Gaffney, O.; Ludwig, C. The trajectory of the Anthropocene: The Great Acceleration. Anthr. Rev. 2015, 2, 81–98. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; John, C.; Coulson, J.C.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef]
- Berg, E.E.; Henry, J.D.; Fastie, C.L.; De Volder, A.D.; Matsuoka, S.M. Spruce beetle outbreaks on the Kenai Peninsula, Alaska, and Kluane National Park and Reserve, Yukon Territory: Relationship to summer temperatures and regional differences in disturbance regimes. For. Ecol. Manag. 2006, 227, 219–232. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Ranger, C.M.; Reding, M.E.; Persad, A.B.; Herms, D.A. Ability of stress-related volatiles to attract and induce attacks by Xylosandrus germanus and other ambrosia beetles. Agric. For. Entomol. 2010, 12, 177–185. [Google Scholar] [CrossRef]
- Gely, C.; Laurance, S.G.W.; Stork, N.E. How do herbivorous insects respond to drought stress in trees? Biol. Rev. 2020, 95, 434–448. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Young, N.E. Assessing forest vulnerability and the potential distribution of pine beetles under current and future climate scenarios in the Interior West of the US. For. Ecol. Manag. 2011, 262, 307–316. [Google Scholar] [CrossRef]
- Preisler, H.K.; Hicke, J.A.; Ager, A.A.; Hayes, J.L. Climate and weather influences on spatial temporal patterns of mountain pine beetle populations in Washington and Oregon. Ecology 2012, 93, 2421–2434. [Google Scholar] [CrossRef]
- Creeden, E.P.; Hicke, J.A.; Buotte, P.C. Climate, weather, and recent mountain pine beetle outbreaks in the western United States. For. Ecol. Manag. 2014, 312, 239–251. [Google Scholar] [CrossRef]
- Lantschner, M.V.; Atkinson, T.H.; Corley, J.C.; Liebhold, A.M. Predicting North American Scolytinae invasions in the Southern Hemisphere. Ecol. Appl. 2017, 27, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, O.; Karaceylan, I.B.; Sen, I. Maximum entropy modeling (maxent) of current and future distributions of Ips mannsfeldi (Wachtl, 1879) (Curculionidae: Scolytinae) in Turkey. Appl. Ecol. Environ. Res. 2018, 16, 2527–2535. [Google Scholar] [CrossRef]
- Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Mandelshtam, M.Y.; Musolin, D.L.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agric. For. Entomol. 2019, 21, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Urvois, T.; Auger-Rozenberg, M.A.; Roques, A.; Rossi, J.P.; Kerdelhue, C. Climate change impact on the potential geographical distribution of two invading Xylosandrus ambrosia beetles. Sci. Rep. 2021, 11, 1339. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.H.; Smith, W.D. Spatio-Temporal Analysis of Xyleborus glabratus (Coleoptera: Circulionidae: Scolytinae) Invasion in Eastern U.S. Forests. Environ. Entomol. 2008, 37, 442–452. [Google Scholar] [CrossRef]
- Ge, X.; Jiang, C.; Chen, L.; Qiu, S.; Zhao, Y.; Wang, T.; Zong, S. Predicting the potential distribution in China of Euwallacea fornicatus (Eichhoff) under current and future climate conditions. Sci. Rep. 2017, 7, 906. [Google Scholar] [CrossRef] [Green Version]
- Kirkendall, L.R.; Dal Cortivo, M.; Gatti, E. First record of the ambrosia beetle, Monarthrum mali (Curculionidae, Scolytinae) in Europe. J. Pest Sci. 2008, 81, 175–178. [Google Scholar] [CrossRef]
- Tremblay, E.; Espinosa, B.; Mancini, D.; Caprio, G. Un coleottero proveniente dal Sudamerica minaccia i pioppi. Inf. Agrar. 2000, 56, 89–90. [Google Scholar]
- Atkinson, T.H. Bark and Ambrosia Beetles. Available online: https://www.barkbeetles.info/regional_chklist_target_species.php?lookUp=3080#table (accessed on 23 November 2018).
- GBIF. GBIF Occurrence download for Gnathotrichus materiarius Fitch. Retrieved from Global Biodiversity Information Facility. Available online: https://doi.org/10.15468/dl.3yenre (accessed on 2 February 2021).
- Atkinson, T.H.; Rabaglia, R.J.; Peck, S.B.; Foltz, J.L. New records of Scolytidae and Platypodidae (Coleoptera) from the United States and the Bahamas. Coleopt. Bull. 1991, 45, 152–164. [Google Scholar]
- Coyle, D.R.; Booth, D.C.; Wallace, M.S. Ambrosia beetle (Coleoptera: Scolytidae) species, flight, and attack on living eastern cottonwood trees. J. Econ. Ent. 2005, 98, 2049–2057. [Google Scholar] [CrossRef]
- Dodge, H.R. The bark beetles of Minnesota (Coleoptera: Scolytidae). University of Minnesota Agricultural Experiment Station. Tech. Bull. 1938, 132, 60. [Google Scholar]
- Goldarazena, A.; Bright, D.E.; Hishinuma, S.M.; López, S.; Seybold, S.J. First record of Pityophthorus solus (Blackman, 1928) in Europe. EPPO Bull. 2014, 44, 65–69. [Google Scholar] [CrossRef]
- Helm, C.; Molano-Flores, B. New records of native and non-native bark and ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) in Illinois. Great Lakes Entomol. 2015, 48, 137–149. [Google Scholar]
- Hines, J.W.; Heikkenen, H.J. Beetles attracted to severed Virginia pine (Pinus virginiana Mill.). Environ. Entomol. 1977, 6, 123–127. [Google Scholar] [CrossRef]
- LaBonte, J.R.; Mudge, A.D.; Johnson, K.J.R. Nonindigenous woodboring Coleoptera (Cerambycidae, Curculionidae: Scolytinae) new to Oregon and Washington, 1999-2002: Consequences of the intracontinental movement of raw wood products and solid wood packing materials. Proc. Ent. Soc. Wash. 2005, 107, 554–564. [Google Scholar]
- Mudge, A.D.; LaBonte, J.R.; Johnson, K.J.R.; LaGasa, E.H. Exotic woodboring Coleoptera (Micromalthidae, Scolytidae) and Hymenoptera (Xiphydriidae) new to Oregon and Washington. Proc. Ent. Soc. Wash. 2001, 103, 1011–1019. [Google Scholar]
- Oliver, J.B.; Mannion, C.M. Ambrosia beetle (Coleoptera: Scolytidae) species attacking chestnut and captured in ethanol-baited traps in middle Tennessee. Environ. Entomol. 2001, 30, 909–918. [Google Scholar] [CrossRef]
- Rabaglia, R.J.; & Valenti, M.A. Annotated list of the bark and ambrosia beetles (Coleoptera: Scolytidae) of Delaware, with new distribution records. Proc. Entomol. Soc. Wash. 2003, 105, 312–319. [Google Scholar]
- Reed, S.E.; Muzika, R.M. The influence of forest stand and site characteristics on the composition of exotic dominated ambrosia beetle communities (Coleoptera: Curculionidae: Scolytinae). Environ. Entomol. 2010, 39, 1482–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeper, R.A.; Hazen, C.R.; Helsel, D.K.; Bunce, M.A. Studies on Michigan ambrosia fungi. Michigan Bot. 1980, 19, 69–73. [Google Scholar]
- Weber, B.C.; MacPherson, J.E. Seasonal Flight Patterns Of Scolytidae (Coleoptera) In Black-Walnut Plantations In North-Carolina And Illinois. Coleopt. Bull. 1991, 45, 4556. [Google Scholar]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982; pp. 1–1356. [Google Scholar]
- Wood, S.L.; Bright, D.E. A Catalog of Scolytidae and Platypodidae (Coleoptera), Part 2. Taxonomic Index (Volumes A, B); Brigham Young University: Provo, UT, USA, 1992; pp. 1–1553. [Google Scholar]
- Szołtys, H.; Plewa, R.; Grzywocz, J. Nowe stanowiska Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Curculionidae, Scolytinae) w południowej Polsce. Acta Entomol. Sil. 2019, 27, 1–2. [Google Scholar]
- Bernabo, E. Osservazione sulla presenza in Italia di Xylosandrus germanus (Blandford, 1894) e Gnathotrichus materiarius (Fitch, 1858)(Coleoptera: Scolytidae). Riv. Piemont. Di Stor. Nat. 2000, 21, 255–262. [Google Scholar]
- Witkowski, R. Ecology and Phenology of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera, Scolytinae) in Conditions of Expansion in Poland. Ph.D. Thesis, Poznań University of Life Sciences, Poznań, Poland, 2020. [Google Scholar]
- Rocchini, D.; Garzon-Lopez, C.X. Cartograms tool to represent spatial uncertainty in species distribution. Res. Ideas Outcomes 2017, 3, e12029. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic Predictors for Supporting Ecological Applications in the Conterminous United States; U.S. Department of the Interior: Washington, DC, USA; U.S. Geological Survey: Reston, VA, USA, 2012; pp. 1–10.
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Riahi, K.; Vuuren, D.P.; van Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Change 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Anibaba, Q.A.; Dyderski, M.K.; Jagodziński, A.M. Predicted range shifts of invasive giant hogweed (Heracleum mantegazzianum) in Europe. Sci. Total Environ. 2022, 825, 154053. [Google Scholar] [CrossRef] [PubMed]
- Polaina, E.; Soultan, A.; Pärt, T.; Recio, M.R. The future of invasive terrestrial vertebrates in Europe under climate and land-use change. Environ. Res. Lett. 2021, 16, 044004. [Google Scholar] [CrossRef]
- Seidl, R.; Klonner, G.; Rammer, W.; Essl, F.; Moreno, A.; Neumann, M.; Dullinger, S. Invasive alien pests threaten the carbon stored in Europe’s forests. Nat. Commun. 2018, 9, 1626. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. Version 1.3-3URL. 2020. Available online: https://CRAN.R-project.org/package=dismo (accessed on 17 December 2020).
- Hijmans, R.J.; van Etten, J.; Sumner, M.; Cheng, J.; Baston, D.; Bevan, A.; Bivand, R.; Busetto, L.; Canty, M.; Fasoli, B.; et al. Raster: Geographic Data Analysis and Modeling. Version 3.3-13. 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 17 December 2020).
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–466. [Google Scholar] [CrossRef] [Green Version]
- Goberville, E.; Beaugrand, G.; Hautekèete, N.-C.; Piquo, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef]
- Thuiller, W.; Guéguen, M.; Renaud, J.; Karger, D.N.; Zimmermann, N.E. Uncertainty in ensembles of global biodiversity scenarios. Nat. Commun. 2019, 10, 1446. [Google Scholar] [CrossRef]
- Paź-Dyderska, S.; Jagodziński, A.M.; Dyderski, M.K. Possible changes in spatial distribution of walnut (Juglans regia L.) in Europe under warming climate. Reg. Environ. Chang. 2021, 21, 18. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Global Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, I. Time lags in biological invasions with regard to the success and failure of alien species. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 15–38. [Google Scholar]
- Rassati, D.; Faccoli, M.; Battisti, A.; Marini, L. Habitat and climatic preferences drive invasions of non-native ambrosia beetles in deciduous temperate forests. Biol. Invasions 2016, 18, 2809–2821. [Google Scholar] [CrossRef]
- Bentz, B.J.; Jönsson, A.M. Modeling Bark Beetle Responses to Climate Change. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 533–553. [Google Scholar] [CrossRef]
- Kelsey, R.G.; Gallego, D.; Sánchez-García, F.J.; Pajares, J.A. Ethanol accumulation during severe drought may signal tree vulnerability to detection and attack by bark beetles. Can. J. For. Res. 2014, 44, 554–561. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Jactel, H.; Koricheva, J.; Castagneyrol, B. Responses of forest insect pests to climate change: Not so simple. Curr. Opin. Insect Sci. 2019, 35, 103–108. [Google Scholar] [CrossRef]
- Dodds, K.J. Effects of trap height on captures of arboreal insects in pine stands of northeastern United States of America. Can. Entomol. 2014, 146, 80–89. [Google Scholar] [CrossRef]
- Thurm, E.A.; Hernandez, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Zlatanov, T.M.; Hladnik, D.; Balic, B.; Freudenschuss, A.; et al. Alternative tree species under climate warming in managed European forests. For. Ecol. Manag. 2018, 430, 485–497. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; Krokene, P.; Liebhold, A.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.; Schelhaas, M.-J.; Seidl, R.; Svoboda, M.; et al. Living with Bark Beetles: Impacts, Outlook and Management Options; European Forest Institute: Joensuu, Finland, 2019; pp. 1–55. [Google Scholar]
- Richardson, D.M.; Whittaker, R.J. Conservation biogeography–foundations, concepts and challenges. Divers. Distrib. 2010, 16, 313–320. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Parameter |
|---|---|
| BIO2 | Mean Diurnal Range (Mean of monthly (max–temp–min temp)) (°C) |
| BIO7 | Temperature Annual Range (BIO5-BIO6) (°C) |
| BIO11 | Mean Temperature of Coldest Quarter (°C) |
| BIO15 | Precipitation Seasonality (Coefficient of Variation: mean/SD × 100) (%) |
| BIO16 | Precipitation of Wettest Quarter (mm) |
| BIO17 | Precipitation of Driest Quarter (mm) |
| Shared Socioeconomic Pathway (SSP) and Timeline | Range Expansion (Absent/Present) (%) | Persistence (Present/Present) (%) | Range Contraction (Present/Absent) (%) | Net Shift (Expansion–Contraction) |
|---|---|---|---|---|
| SSP126 2041–2060 | 7.53 | 11.35 | 0.25 | 7.28 |
| SSP245 2041–2060 | 9.09 | 11.27 | 0.32 | 8.77 |
| SSP370 2041–2060 | 10.39 | 11.15 | 0.45 | 9.94 |
| SSP585 2041–2060 | 12.53 | 11.13 | 0.47 | 12.06 |
| SSP126 2061–2080 | 8.04 | 11.29 | 0.31 | 7.73 |
| SSP245 2061–2080 | 11.88 | 11.14 | 0.46 | 11.43 |
| SSP370 2061–2080 | 16.27 | 11.05 | 0.55 | 15.72 |
| SSP585 2061–2080 | 18.86 | 10.87 | 0.72 | 18.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witkowski, R.; Dyderski, M.K.; Bełka, M.; Mazur, A. Potential European Geographical Distribution of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Scolytinae) under Current and Future Climate Conditions. Forests 2022, 13, 1097. https://doi.org/10.3390/f13071097
Witkowski R, Dyderski MK, Bełka M, Mazur A. Potential European Geographical Distribution of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Scolytinae) under Current and Future Climate Conditions. Forests. 2022; 13(7):1097. https://doi.org/10.3390/f13071097
Chicago/Turabian StyleWitkowski, Radosław, Marcin K. Dyderski, Marta Bełka, and Andrzej Mazur. 2022. "Potential European Geographical Distribution of Gnathotrichus materiarius (Fitch, 1858) (Coleoptera: Scolytinae) under Current and Future Climate Conditions" Forests 13, no. 7: 1097. https://doi.org/10.3390/f13071097