
Characterization of the Complete Chloroplast Genome Sequence of the Socotra Dragon`s Blood Tree (Dracaena cinnabari Balf.)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Isolation
2.2. Library Preparing and Sequencing
2.3. Genome Assembly and Annotation
2.4. Genome Comparative Analysis
2.5. Identification of Simple Sequence Repeats and Phylogenetic Inference
2.6. Phylogenetic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marrero, A.; Almeida, R.; Gonzalez-Martin, M. A new species of the wild dragon tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic implications. J. Linn. Soc. Bot. 1998, 128, 291–314. [Google Scholar] [CrossRef]
- Maděra, P.; Forrest, A.; Hanáček, P.; Vahalík, P.; Gebauer, R.; Plichta, R.; Jupa, R.; Van Rensburg, J.J.; Morris, M.; Nadezhdina, N.; et al. What We Know and What We Do Not Know about Dragon Trees? Forests 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.G.; Morris, M. Ethnoflora of the Soqotra Archipelago, 1st ed.; Atkinson, R., Ed.; Royal Botanic Gardens: Edinburgh, UK, 2004; p. 759. [Google Scholar]
- Al-Okaishi, A. Exploring the Historical Distribution of Dracaena cinnabari Using Ethnobotanical Knowledge on Socotra Island, Yemen. J. Ethnobiol. Ethnomed. 2021, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a Single Tree Species Will Lead to an Overall Decline in Plant Diversity: Effect of Dracaena cinnabari Balf. f. on the Vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- Vasconcelos, R.; Pujol-Buxó, E.; Llorente, G.A.; Saeed, A.; Carranza, S. Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests 2020, 11, 353. [Google Scholar] [CrossRef] [Green Version]
- Adolt, R.; Pavlis, J. Age Structure and Growth of Dracaena cinnabari Populations on Socotra. Trees Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will Dragonblood Survive the next Period of Climate Change? Current and Future Potential Distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Habrova, H.; Cermak, Z.; Pavlis, J. Dragon’s Blood Tree—Threatened by Overmaturity, Not by Extinction: Dynamics of a Dracaena cinnabari Woodland in the Mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth Dynamics of Endemic Dracaena cinnabari Balf. f. of Socotra Island Suggest Essential Elements for a Conservation Strategy. Biologia 2019, 74, 339–349. [Google Scholar] [CrossRef]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth Dynamics of Dracaena cinnabari under Controlled Conditions as the Most Effective Way to Protect Endangered Species. Saudi J. Biol. Sci. 2017, 24, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Sap Flow Measurements in a Socotra Dragon’s Blood Tree (Dracaena cinnabari) in Its Area of Origin. Trop. Plant Biol. 2018, 11, 107–118. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- Miller, A. Dracaena cinnabari, Dragon’s Blood Tree. IUCN Red List Threat. Species 2004, e.T30428A9548491. [Google Scholar] [CrossRef]
- Klimko, M.; Nowińska, R.; Wilkin, P.; Wiland-Szymańska, J. Comparative Leaf Micromorphology and Anatomy of the Dragon Tree Group of Dracaena (Asparagaceae) and Their Taxonomic Implications. Plant Syst. Evol. 2018, 304, 1041–1055. [Google Scholar] [CrossRef] [Green Version]
- Lautenschläger, T.; Frommherz, L.; Monizi, M.; Neinhuis, C.; Henle, T.; Förster, A. Anatomy and Nutritional Value of Dracaena Camerooniana Baker—An African Wild Vegetable. S. Afr. J. Bot. 2021, 145, 149–156. [Google Scholar] [CrossRef]
- Jura-Morawiec, J.; Wiland-Szymańska, J. A Novel Insight into the Structure of Amphivasal Secondary Bundles on the Example of Dracaena draco L. Stem. Trees Struct. Funct. 2014, 28, 871–877. [Google Scholar] [CrossRef]
- Jura-Morawiec, J. Formation of Amphivasal Vascular Bundles in Dracaena draco Stem in Relation to Rate of Cambial Activity. Trees Struct. Funct. 2015, 29, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Wang, X.; Chen, J.; Lu, Y.; Wu, N.; Kang, W.; Zheng, Q. Studies on the Chemical Constituents of Chloroform Extract of Dracaena Cochinchinensis. Acta Pharm. Sin. 1998, 33, 755–758. [Google Scholar]
- Lu, P.L.; Morden, C.W.; Manning, J. Phylogenetic Relationships among Dracaenoid Genera (Asparagaceae: Nolinoideae) Inferred from Chloroplast DNA Loci. Syst. Bot. 2014, 39, 90–104. [Google Scholar] [CrossRef]
- Edwards, C.E.; Bassüner, B.; Birkinshaw, C.; Camara, C.; Lehavana, A.; Lowry, P.P.; Miller, J.S.; Wyatt, A.; Jackson, P.W. A Botanical Mystery Solved by Phylogenetic Analysis of Botanical Garden Collections: The Rediscovery of the Presumed-Extinct Dracaena Umbraculifera. Oryx 2018, 52, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Chai, X.; Rao, Y.; Tu, X.; Du, S. The Complete Chloroplast Genome Sequence of Asparagus (Asparagus officinalis L.) and Its Phylogenetic Positon within Asparagales. Int. J. Plant Biol. Res. 2017, 5, 1075. [Google Scholar]
- Li, J.R.; Li, S.F.; Wang, J.; Dong, R.; Zhu, H.W.; Li, N.; Deng, C.L.; Gao, W.J. Characterization of the Complete Chloroplast Genome of Asparagus Setaceus. Mitochondrial DNA Part B Resour. 2019, 4, 2639–2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, G.; Park, S.; Lee, E.M.; Park, S.J. Evidence of Mitochondrial DNA in the Chloroplast Genome of Convallaria Keiskei and Its Subsequent Evolution in the Asparagales. Sci. Rep. 2019, 9, 5028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Song, M.; Guan, Y.; Ma, X. Species Identification of Dracaena Using the Complete Chloroplast Genome as a Super-Barcode. Front. Pharmacol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Celiński, K.; Kijak, H.; Wiland-Szymańska, J. Complete Chloroplast Genome Sequence and Phylogenetic Inference of the Canary Islands Dragon Tree (Dracaena draco L.). Forests 2020, 11, 309. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L. Isolation of Plants DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an Integrated Plastome Sequence Annotator and Analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and Accurate Annotation of Organelle Genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. TRNAscan-SE 2.0: Improved Detection and Functional Classification of Transfer RNA Genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-Web: A Web Server for Microsatellite Prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Liu, Y.; Yuan, Q.; Sun, J.; Guo, L. Chloroplast genome variation and phylogenetic relationships of Atractylodes species. BMC Genomics 2021, 22, 103. [Google Scholar] [CrossRef]
- Xu, J.; Shen, X.; Liao, B.; Xu, J.; Hou, D. Comparing and phylogenetic analysis chloroplast genome of three Achyranthes species. Sci Rep. 2020, 10, 10818. [Google Scholar] [CrossRef]
- Chen, Q.; Hu, H.; Zhang, D. DNA Barcoding and Phylogenomic Analysis of the Genus Fritillaria in China Based on Complete Chloroplast Genomes. Front. Plant Sci. 2022, 13, 764255. [Google Scholar] [CrossRef]
- Hong, Z.; He, W.; Liu, X.; Tembrock, L.R.; Wu, Z.; Xu, D.; Liao, X. Comparative Analyses of 35 Complete Chloroplast Genomes from the Genus Dalbergia (Fabaceae) and the Identification of DNA Barcodes for Tracking Illegal Logging and Counterfeit Rosewood. Forests 2022, 13, 626. [Google Scholar] [CrossRef]
- Sokołowska, J.; Fuchs, H.; Celiński, K. New Insight into Taxonomy of European Mountain Pines, Pinus Mugo Complex, Based on Complete Chloroplast Genomes Sequencing. Plants 2021, 10, 1331. [Google Scholar] [CrossRef]
- Gu, L.; Su, T.; Luo, G.L.; Hu, G.X. The complete chloroplast genome sequence of Heteropolygonatum ginfushanicum (Asparagaceae) and phylogenetic analysis. Mitochondrial DNA Part B 2021, 6, 1799–1802. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Choi, M.-J.; Kim, S.-H.; Choi, H.-J.; Kim, S.-C. Plastome Characterization and Phylogenomic Analysis Yield New Insights into the Evolutionary Relationships among the Species of the Subgenus Bryocles (Hosta; Asparagaceae) in East Asia. Plants 2021, 10, 1980. [Google Scholar] [CrossRef] [PubMed]
- Takawira-Nyenya, R.; Mucina, L.; Cardinal-Mcteague, W.M.; Thiele, K.R. Sansevieria (Asparagaceae, Nolinoideae) is a herbaceous Glade within Dracaena: Inference from non-coding plastid and nuclear DNA sequence data. Phytotaxa 2018, 376, 254–276. [Google Scholar] [CrossRef]
- Ge, J.; Cai, L.; Bi, G.Q.; Chen, G.; Sun, W. Characterization of the Complete Chloroplast Genomes of Buddleja colvilei and B. sessilifolia: Implications for the Taxonomy of Buddleja L. Molecules 2018, 23, 1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Zou, P.; Jiang, N.; Fang, Y.; Liu, G. Comparative Analysis of the Complete Chloroplast Genomes of Nine Paphiopedilum Species. Front. Genet. 2022, 12, 772415. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Wiland-Szymańska, J.; Buczkowska-Chmielewska, K.; Drapikowska, M.; Maślak, M.; Myszczyński, K.; Szczecińska, M.; Ślipiko, M.; Sawicki, J. The Complete Chloroplast Genome of a Rare Orchid Species Liparis loeselii (L.). Conserv. Genet. Resour. 2018, 10, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H. Microsatellites: Simple Sequences with Complex Evolution. Nat. Rev. Genet. 2004, 5, 435–554. [Google Scholar] [CrossRef]
- Oliveira, E.J.; Pádua, J.G.; Zucchi, M.I.; Vencovsky, R.; Vieira, M.L.C. Origin, Evolution and Genome Distribution of Microsatellites. Genet. Mol. Biol. 2006, 29, 249–307. [Google Scholar] [CrossRef]
- Scarcelli, N.; Barnaud, A.; Eiserhardt, W.; Treier, U.A.; Seveno, M.; d’Anfray, A.; Pintaud, J.C. A set of 100 chloroplast DNA primer pairs to study population genetics and phylogeny in monocotyledons. PLoS ONE 2011, 6, e19954. [Google Scholar] [CrossRef] [Green Version]
- Vendramin, G.G.; Lelli, L.; Rossi, P.; Morgante, M. A set of primers for the amplification of 20 chloroplast microsatellites in Pinaceae. Mol. Ecol. 1996, 5, 595–598. [Google Scholar] [CrossRef] [PubMed]
- González-Martinez, S.C.; Robledo-Arnuncio, J.J.; Collada, C.; Díaz, A.; Williams, C.G.; Alía, R.; Cervera, M.T. Cross-amplification and sequence variation of microsatellite loci in Eurasian hard pines. Theor Appl Genet. 2004, 109, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Celiński, K.; Pawlaczyk, E.M.; Wojnicka-Półtorak, A.; Chudzińska, E.; Prus-Głowacki, W. Cross-species amplification and characterization of microsatellite loci in Pinus mugo Turra. Biologia 2013, 68, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, C.; Lee, Y.M.; Kim, J.H. Development of chloroplast microsatellite markers for the endangered Maianthemum bicolor (Asparagaceae s.l.). Appl. Plant Sci. 2016, 4, 1600032. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
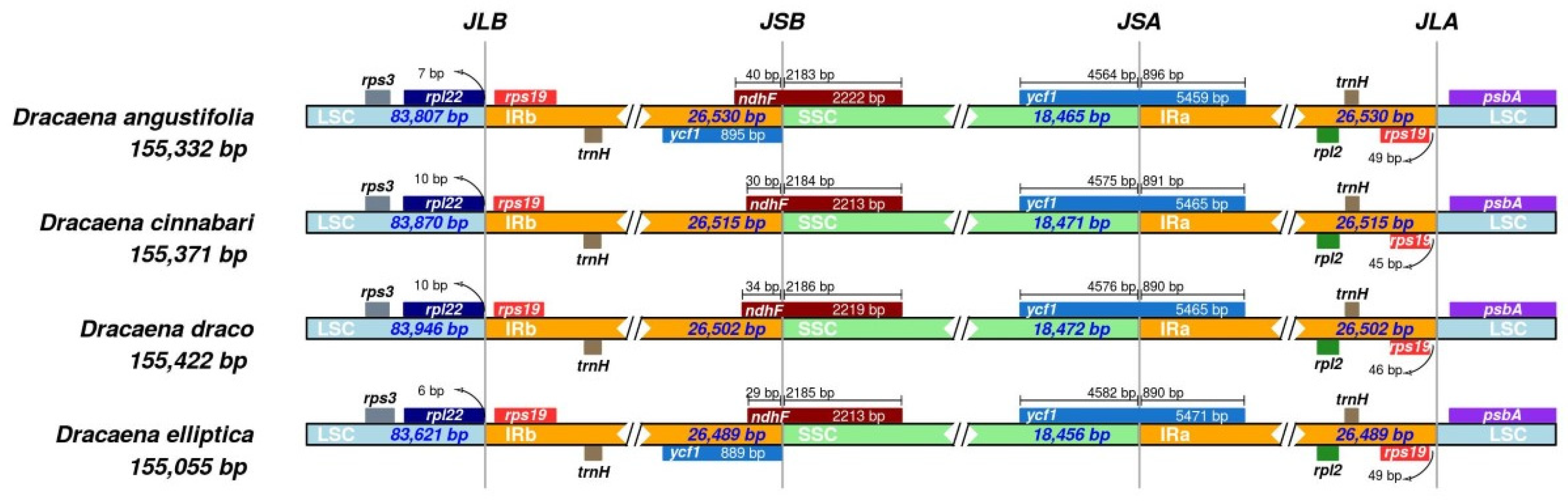{kind=link}
{kind=link}
| Species | D. angustifolia | D. cinnabari | D. draco | D. elliptica |
|---|---|---|---|---|
| Total length (bp) | 155,332 | 155,371 | 155,422 | 155,055 |
| IR length (bp) | 53,060 | 53,030 | 53,004 | 52,978 |
| LSC length (bp) | 83,807 | 83,870 | 83,946 | 83,621 |
| SSC length (bp) | 18,465 | 18,471 | 18,472 | 18,456 |
| Total gene number | 130 | 132 | 132 | 130 |
| rRNA | 8 | 8 | 8 | 8 |
| tRNA | 38 | 38 | 38 | 38 |
| GC content (%) | 37.5 | 37.5 | 37.6 | 37.5 |
| GenBan accession | MN200193 | OM961177 | MN990038 | MN200196 |
| Reference | [25] | This study | [26] | [25] |
| No. | Group of Genes | Name of Genes | Number |
|---|---|---|---|
| 1 | Photosystem I | psaA, psaB, psaC, psaI, psaJ | 5 |
| 2 | Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbNpsbT, psbZ | 15 |
| 3 | Cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN, | 6 |
| 4 | ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI, | 6 |
| 5 | NADH dehydrogenase | ndhA *, ndhB *(×2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | 12 |
| 6 | RubisCO large subunit | rbcL | 1 |
| 7 | RNA polymerase | rpoA, rpoB, rpoC1 *, rpoC2 | 4 |
| 8 | Ribosomal proteins –small units (SSU) | rps2, rps3, rps4, rps7(×2), rps8, rps11, rps12 *(×2), rps14, rps15, rps16 *, rps18, rps19(×2), | 15 |
| 9 | Ribosomal proteins –large units (LSU) | rpl2 *(×2), rpl14, rpl16 *, rpl20, rpl22, rpl23(×2), rpl32, rpl33, rpl36, | 11 |
| 10 | Other genes/ Miscellaneous | accD, ccsA, cemA, clpP **, infA, matK, | 6 |
| 11 | Protein of unknown function | ycf1, ycf2(×2), ycf3 **, ycf4 | 5 |
| 12 | Transfer RNAs | trnA-UGC(x2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnH-GUG(×2), trnI-CAU(×2), trnI-GAU *(×2), trnK-UUU *, trnL-CAA(×2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU(×2), trnP-UGG, trnQ-UUG, trnR-ACG(×2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC(×3), trnV-UAC *, trnW-CCA, trnY-GUA | 38 |
| 13 | Ribosomal RNAs | rrn4.5(×2), rrn5(×2), rrn16(×2), rrn23(×2) | 8 |
| Total | 132 |
| Species | Indices | Motif | Total | |
|---|---|---|---|---|
| Mononucleotide | Dinucleotide | |||
| Total counts | 38 | 4 | 42 | |
| D. angustifolia | Total Repeat Length (bp) | 409 | 54 | |
| Mean Length (bp) | 10.76 | 13.5 | ||
| Total counts | 34 | 6 | 40 | |
| D. cinnabari | Total Repeat Length (bp) | 363 | 78 | |
| Mean Length (bp) | 10.68 | 13 | ||
| Total counts | 56 | 4 | 60 | |
| D. draco | Total Repeat Length (bp) | 638 | 50 | |
| Mean Length (bp) | 11.39 | 12.5 | ||
| Total counts | 44 | 3 | 47 | |
| D.elliptica | Total Repeat Length (bp) | 473 | 36 | |
| Mean Length (bp) | 10.75 | 12 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celiński, K.; Sokołowska, J.; Fuchs, H.; Maděra, P.; Wiland-Szymańska, J. Characterization of the Complete Chloroplast Genome Sequence of the Socotra Dragon`s Blood Tree (Dracaena cinnabari Balf.). Forests 2022, 13, 932. https://doi.org/10.3390/f13060932
Celiński K, Sokołowska J, Fuchs H, Maděra P, Wiland-Szymańska J. Characterization of the Complete Chloroplast Genome Sequence of the Socotra Dragon`s Blood Tree (Dracaena cinnabari Balf.). Forests. 2022; 13(6):932. https://doi.org/10.3390/f13060932
Chicago/Turabian StyleCeliński, Konrad, Joanna Sokołowska, Hanna Fuchs, Petr Maděra, and Justyna Wiland-Szymańska. 2022. "Characterization of the Complete Chloroplast Genome Sequence of the Socotra Dragon`s Blood Tree (Dracaena cinnabari Balf.)" Forests 13, no. 6: 932. https://doi.org/10.3390/f13060932