
Changes in the Microbiological Properties of Soils along the Gradient of the Altitude Zone of Mount Kivaka in Eastern Fennoscandia, Russia


Abstract
:1. Introduction
2. Materials and Methods
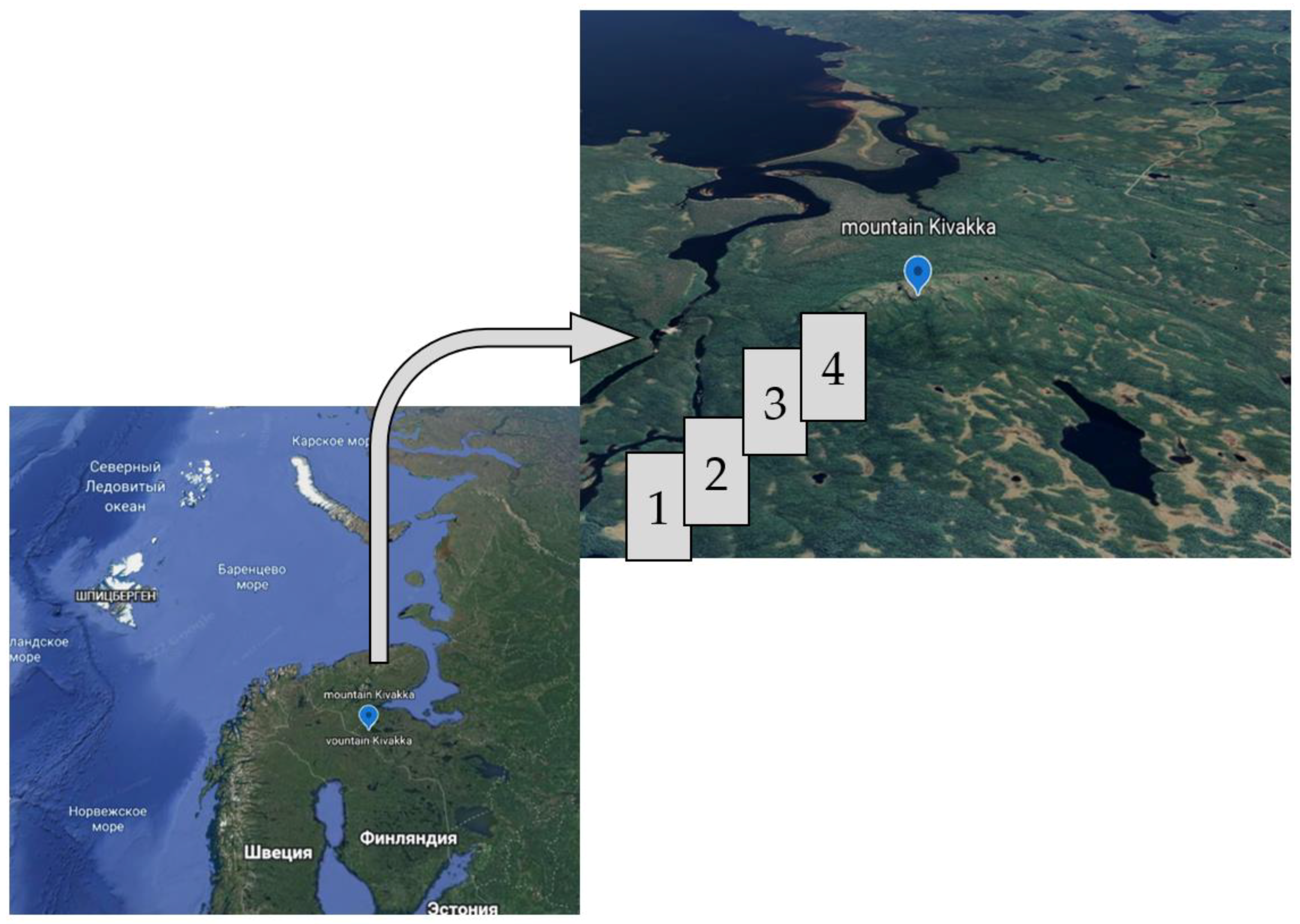
2.1. Study Area
- taiga forest ecosystem (TF),
- forest of mountain (FM),
- mixed forest (MF), boundary between taiga forest and forest mountain
- forest tundra, (FT), subgoltsy zone (Table 1).
2.2. Soil Sampling and Microbiological Analysis
2.3. Statistical Analysis
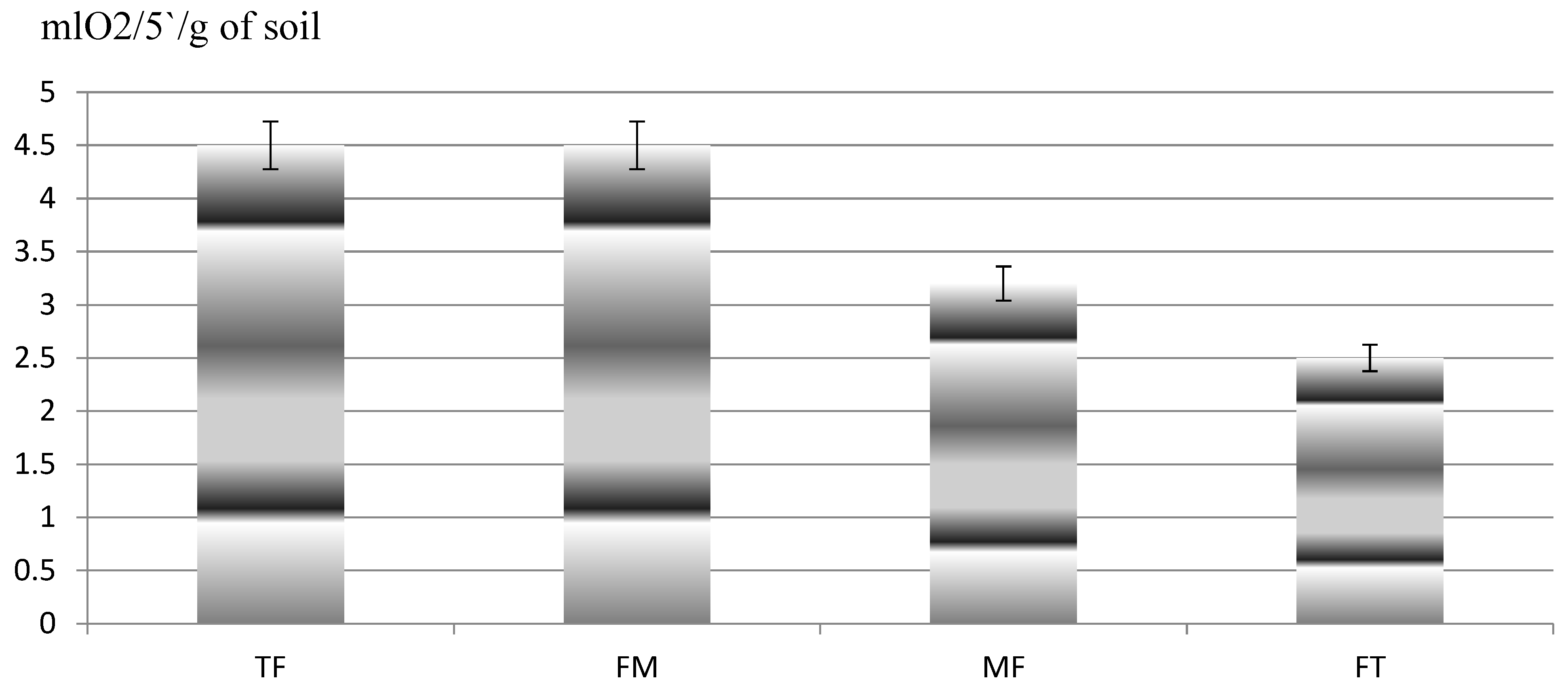
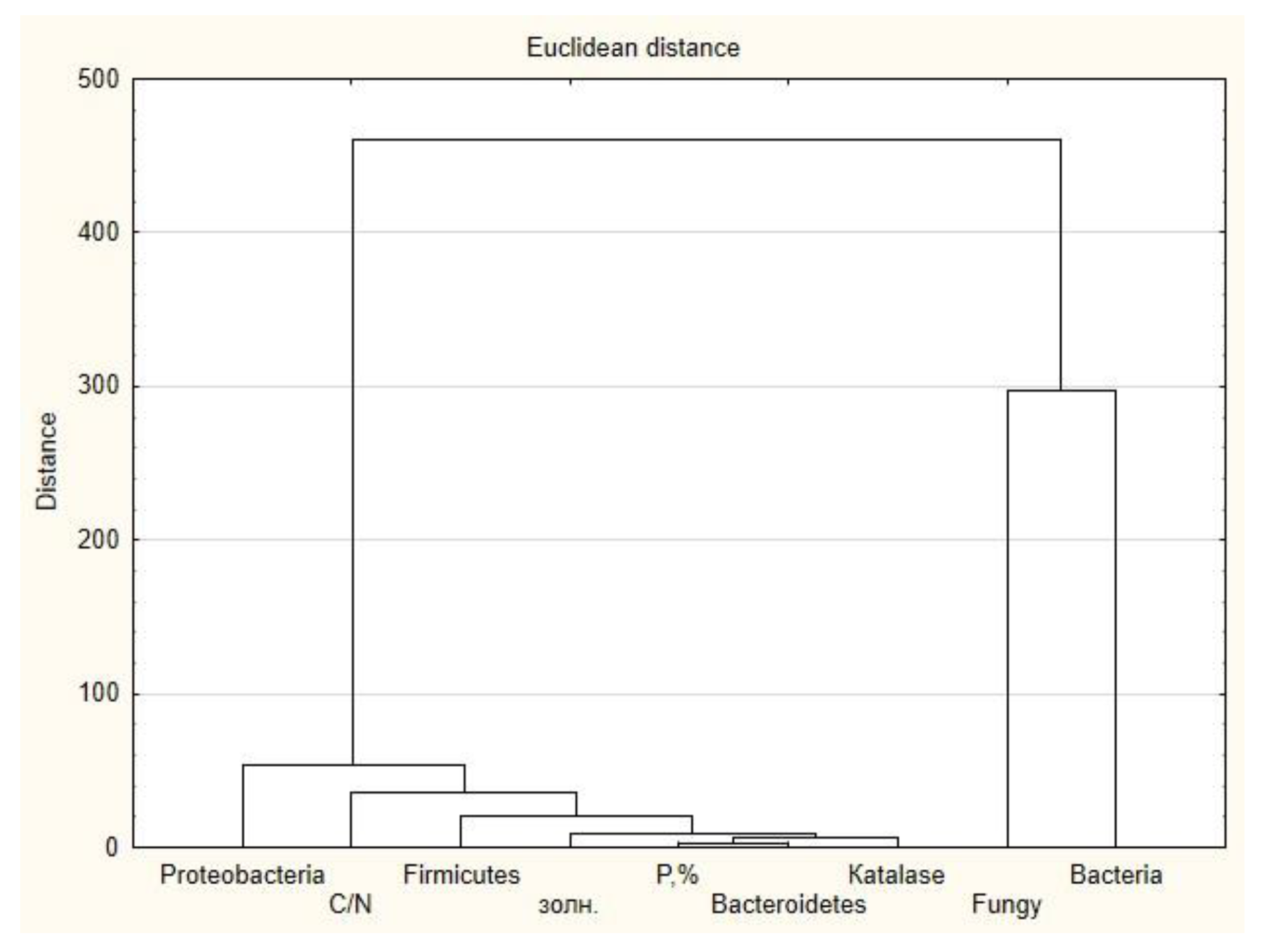
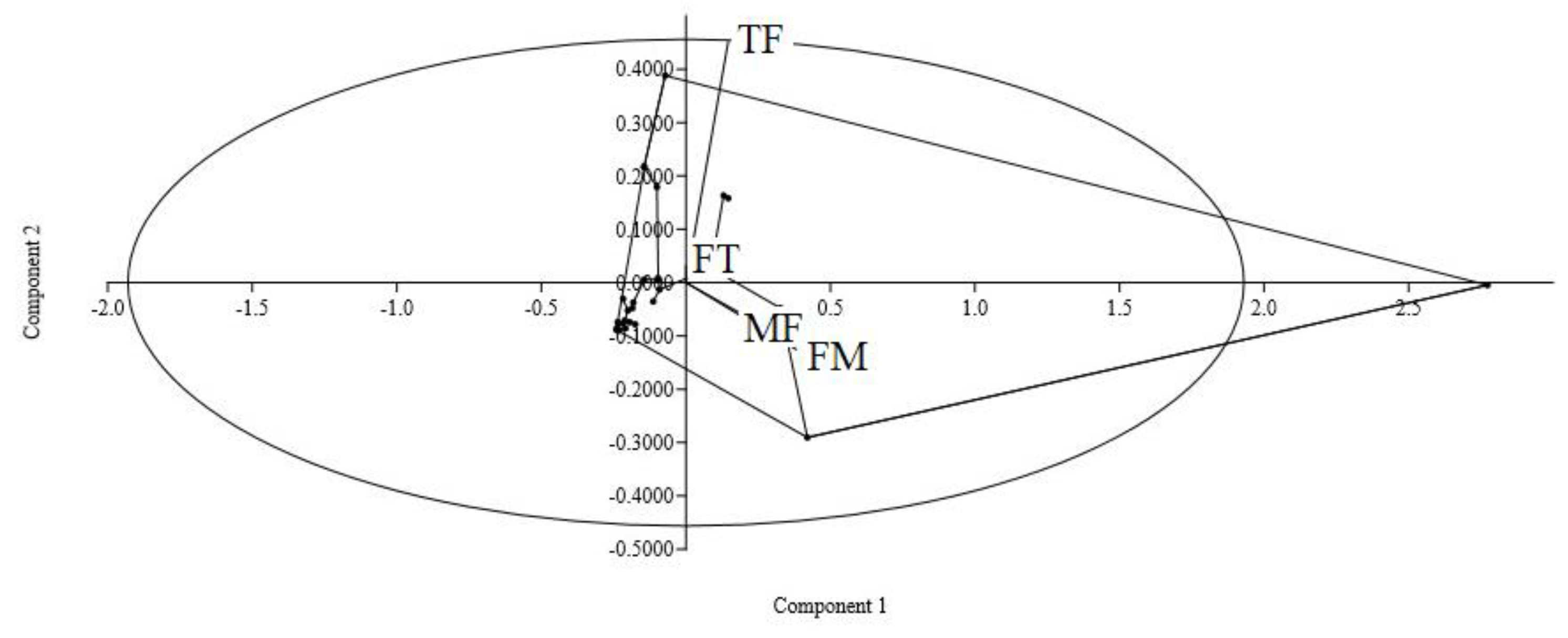
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Andersen, R.; Grasset, L.; Thormann, M.N.; Rochefort, L.; Francez, A.-J. Changes in microbial community structure and function following Sphagnum peatland restoration. Soil Biol. Biochem. 2010, 42, 291–301. [Google Scholar] [CrossRef]
- Bailey, V.L.; Smith, J.L.; Bolton, H. Fungal-to-bacterial ratios in soils investigated for enhanced C sequestration. Soil Biol. Biochem. 2002, 34, 997–1007. [Google Scholar] [CrossRef]
- Bao, K.; Yu, X.; Jia, L.; Wang, G. Recent carbon accumulation in Changbai Mountain peatlands, Northeast China. Mt. Res. Dev. 2010, 30, 33–41. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.L.; Laval, B.; Lim, D.S.S.; Slater, G.F. Autotrophic and heterotrophic associated biosignatures in modern freshwater microbialites over seasonal and spatial gradients. Org. Geochem. 2014, 67, 8–18. [Google Scholar] [CrossRef]
- Bruggen, A.H.C.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]
- Bychkova, Y.V.; Koptev-Dvornikov, E.V.; Kononkova, N.N.; Kameneva, E.E. Composition of rockforming minerals in the Kivakka layered massif, northern Karelia, and systematic variations in the chemistries of minerals in the rhythmic layering subzone. Geochem. Int. 2007, 45, 131–151. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697, PMCID:PMC92043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, P.R.; Ehrlich, A.H. Extinction: The Causes and Consequences of the Disappearance of Species; Random House: New York, NY, USA, 1981; 320p. [Google Scholar]
- Djukic, I.; Zehetner, F.; Mentler, A.; Gerzabek, M.H. Microbial community composition and activity in different Alpine vegetation zones. Soil Biol. Biochem. 2010, 42, 155–161. [Google Scholar] [CrossRef]
- Joyce, L.A.; Talbert, M.; Sharp, D.; Stevenson, J. Historical and projected climate in the northern rockies region. In Climate Change and Rocky Mountain Ecosystems. Advances in Global Change Research; Halofsky, J., Peterson, D., Eds.; Springer: New York, NY, USA, 2018; pp. 17–23. [Google Scholar]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Hanzelová, M.; Vido, J.; Jaroslav, S.; Nalevanková, P.; Perhacova, Z. Microorganisms in summer snow patches in selected high mountain ranges of Slovakia. Biologia 2018, 73, 1177–1186. [Google Scholar] [CrossRef]
- Ivashchenko, K.; Sushko, S.; Selezneva, A.; Ananyeva, N.; Zhuravleva, A.; Kudeyarov, V.; Makarov, M.; Blagodatsky, S. Soil microbial activity along an altitudinal gradient: Vegetation as a main driver beyond topographic and edaphic factors. Appl. Soil Ecol. 2021, 168, 104197. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Kuan, H.L.; Ritz, K.; Glover, L.A.; McCaig, A.E.; Fenwick, C. The relationship between microbial community structure and functional stability, tested experimentally in an upland pasture soil. Microb. Ecol. 2004, 47, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Yu, W.; Dutta, S.; Gao, H. Soil microbial community composition and function are closely associated with soil organic matter chemistry along a latitudinal gradient. Geoderma 2021, 383, 114744. [Google Scholar] [CrossRef]
- Kotas, P.; Šantrůčková, H.; Elster, J.; Kaštovská, E. Soil microbial biomass, activity and community composition along altitudinal gradients in the High Arctic (Billefjorden, Svalbard). Biogeosciences 2018, 15, 1879–1894. [Google Scholar] [CrossRef] [Green Version]
- Lavrov, M. Hyperbasites and Stratification of Peridotite-Gabbro-Norite Intrusions of the Precambrian in the North Karelia; Nauka: Leningrad, Russia, 1979; pp. 51–101. [Google Scholar]
- Loreau, M. Biodiversity and ecosystem functioning: Recent theoretical advances. Oikos 2000, 91, 3–17. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Allen, C.D.; Marshall, L. Growth, carbon-isotope discrimination, and drought-associated mortality across a Pinus ponderosa elevational transect. Glob. Chang. Biol. 2010, 16, 399–415. [Google Scholar] [CrossRef]
- Medvedeva, M.V.; Akhmetova, G.V.; Fedorets, N.G.; Yakovlev, A.S.; Raevskii, B.V.; Travin, V.V. Soils of Low-Mountain Landscapes of North Karelia. Eurasian Soil Sci. 2018, 2, 131–140. [Google Scholar] [CrossRef]
- Montpellier, E.; Soul, P.; Knapp, P.; Maxwell, J. Dendroclimatic assessment of Ponderosa Pine radial growth along elevational transects in Western Montana, U.S.A. Forests 2019, 10, 1094. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Marhan, S.; Kandeler, E.; Poll, C. Carbon flow from litter through soilmicroorganisms: From incorporation rates to mean residence times in bacteria and fungi. Soil Biol. Biochem. 2017, 115, 187–196. [Google Scholar] [CrossRef]
- Naeem, S.; Li, S. Biodiversity enhances ecosystem reliability. Nature 1997, 390, 507–509. [Google Scholar] [CrossRef]
- Niklinska, M.; Klimek, B. Effect of temperature on the respiration rate of forest soil organic layer along an elevation gradient in the Polish Carpathians. Biol. Fertil. Soils 2007, 43, 511–518. [Google Scholar] [CrossRef]
- Osipov, G.A. Chromato-Mass Spectrometric Study of Microorganisms and Their Communities. Autoref. diss. d. biol.; Nauk: Moscow, Russia, 1995; 62p. [Google Scholar]
- Osipov, G.A. Method for Determining the Generic (Species) Composition of the Association of Microorganisms. RF Patent No. 2086642 C12N 1/00, 1/20, C12Q ¼, 24 December 1993. [Google Scholar]
- Rautio, P.; Fürst, A.; Stefan, K.; Raitio, H.; Bartels, U. Sampling and analysis of needles and leaves. 19 pp. Manual Part XII. In Manual and Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects to Air Pollution on Forests; UNECE ICP Forest Programme Coordinating Centre: Hamburg, Germany, 2010. [Google Scholar]
- Plant Guide. Available online: https://plants.usda.gov/plantguide/pdf/pg_pipo.pdf (accessed on 7 September 2019).
- Rehfeldt, G.E. Genetic differentiation among populations of Pinus ponderosa from the upper Colorado River Basin. Bot. Gaz. 1990, 1, 125–137. [Google Scholar] [CrossRef]
- Rudkovskaya, O.A.; Rayevsky, B.V.; Ilyinov, A.A.; Medvedeva, M.V. Comparative assessment of biodiversity of plant communities of the low mountains of the National Park “Paanayarvi”. Lesn. Vestn. 2016, 20, 26–35. [Google Scholar]
- Segura, J.H.; Nilsson, M.B.; Schleucher, J.; Haei, M.; Sparrman, T.; Székely, A.; Bertilsson, S.; Öquist, M.G. Microbial utilization of simple carbon substratesin boreal peat soils at low temperatures. Soil Biol. Biochem. 2019, 135, 438–448. [Google Scholar] [CrossRef]
- Segura, J.H.; Nilsson, M.B.; Haei, M.; Sparrman, T.; Mikkola, J.-P.; Gräsvik, J.; Schleucher, J.; Öquist, M.G. Microbial mineralization of cellulose in frozen soils. Nat. Commun. 2017, 8, 1154. [Google Scholar] [CrossRef] [Green Version]
- Shekhovtsova, N.; Marakaev, O.; Pervushina, K.; Osipov, G. The underground organ microbial complexes of moorland spotted orchid Dactylorhiza maculata (L.) Soó (Orchidaceae). Adv. Biosci. Biotechnol. 2013, 04, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Siles, J.A.; Cajthaml, T.; Filipova, A.; Minerbi, S.; Margesin, R. Altitudinal, seasonal and inter annual shifts in microbial communities and chemical composition of soil organic matter in Alpine forest soils. Soil Biol. Biochem. 2017, 112, 1–13. [Google Scholar] [CrossRef]
- Siles, J.A.; Cajthaml, T.; Minerbi, S.; Margesin, R.; Max, H. Effect of altitude and season on microbial activity, abundance and community structure in Alpine forest soils. FEMS Microbiol. Ecol. 2016, 92, fiw008. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Gunina, A.; Luo, Y.; Wang, J.; He, J.; Kuzyakov, Y.; Hemp, A.; Classen Aim, T.; Ge, Y. Contrasting patterns and drivers of soil bacterial and fungal diversity across a mountain gradient. Environ. Microbiol. 2020, 22, 3287–3301. [Google Scholar] [CrossRef]
- Sundh, I.; Nilsson, M.; Borga, P. Variation in microbial community structure in two boreal peatlands as determined by analysis of phospholipid fatty acid profiles. Am. Soc. Microbiol. 1997, 63, 1476–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundqvist, M.K.; Liu, Z.; Giesler, R.; Wardle, D.A. Plant and microbial responses to nitrogen and phosphorus addition across an elevational gradient in subarctic tundra. Ecology 2014, 95, 1819–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorobyeva, L.A. Chemical Analysis of Soils: Textbook; Publishing House of Moscow State University: Moscow, Russia, 1998; p. 272p. [Google Scholar]
- Xu, M.; Li, X.L.; Cai, X.B.; Gai, J.P.; Li, X.L.; Christie, P.; Zhang, J.L. Soil microbial community structure and activity along a montane elevational gradient on the Tibetan. Plateau. Eur. J. Soil Biol. 2014, 64, 6–14. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.; Cao, Y.; Jiang, M.; Wang, G.; Dong, Y. The effects of hummock-hollow microtopography on soil organic carbon stocks and soil labile organic carbon fractions in a sedge peatland in Changbai Mountain. China. Catena 2021, 201, 105204. [Google Scholar] [CrossRef]
- Willers, C.; van Rensburg, P.J.; Claassens, S. Phospholipid fatty acid profiling of microbial communities—a review of interpretations and recent applications. J. Appl. Microbiol. 2015, 119, 1207–1218. [Google Scholar] [CrossRef]
- Wu, Y.P.; Ma, B.; Zhou, L.; Wang, H.Z.; Xu, J.M.; Kemmitt, S.; Brookes, P.C. Changes in the soil microbial community structure with latitude in eastern China, based on phospholipid fatty acid analysis. Appl. Soil Ecol. 2009, 3, 234–240. [Google Scholar] [CrossRef]
- WRB, IUSS Working Group WRB. World Reference Base for Soil Resources 2014: Update 2015, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, Word Soil Resources Report 106; Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Xu, Z.W.; Zhang, T.Y.; Wang, S.Z.; Wang, Z.C. Soil pH and C/N ratio determines spatial variations in soil microbial communities and enzymatic activities of the agricultural ecosystems in Northeast China: Jilin Province case. Appl. Soil Ecol. 2020, 155, 103629. [Google Scholar] [CrossRef]
- Xu, Z.W.; Yu, G.R.; Zhang, X.Y.; Ge, J.P.; He, N.P.; Wang, Q.F.; Wang, D. The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain. Appl. Soil Ecol. 2015, 86, 19–29. [Google Scholar] [CrossRef]
- Yu, Z.; Loisel, J.; Brosseau, D.P.; Beilman, D.W.; Hunt, S.J. Global peatland dynamics since the Last Glacial Maximum. Geophys. Res. Lett. 2010, 37. Available online: https://agupubs.onlinelibrary.wiley.com/doi/full/10.1029/2010GL043584 (accessed on 31 December 2010). [CrossRef]
- Zaitlin, B.; Turkington, K.; Parkinson, D.S.; Clayton, G. Effects of tillage and inorganic fertilizers on culturable soil actinomycete communities and inhibion of fungi by specific actinomycetes. Appl. Soil Ecol. 2004, 26, 53–62. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Zhang, B.; Liang, C.; He, H.; Zhang, X. Variations in soil microbial communities and residues along an altitude gradient on the northern slope of Changbai mountain, China. PLoS ONE 2013, 8, e66184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cregg, B.M. Growth and physiological responses to varied environments among populations of Pinus ponderosa. For. Ecol. Manag. 2005, 219, 1–12. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, M.; Zhao, Y.; Wang, G.; Xue, Z.; Jiang, M. Variations in Soil Microbial Communities in the Sedge-Dominated Peatlands along an Altitude Gradient on the Northern Slope of Changbai Mountain, China. Available online: https://www.researchgate.net/publication/353195243_Variations_in_soil_microbial_communities_in_the_sedge-dominated_peatlands_along_an_altitude_gradient_on_the_northern_slope_of_Changbai_Mountain_China (accessed on 19 April 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Altitude m a.s.l. | Soil Type (WRB, 2015) | Dominant Vegetation | Ash, % | C/N | P % |
|---|---|---|---|---|---|
| Taiga forest (TF) | |||||
| 208 | Skeletic Albic Podzol | Picea abies, Vaccinium myrtillus, Avenella flexuosa (L.) Drej., Gymnocarpium dryopteris (L.) Newm., Linnaea borealis L., Luzula pilosa (L.) Willd., Maianthemum bifolium (L.) F. W. Schmidt, Melampyrum pratense L., Vaccinium vitis-idaea L., Pleurozium schreberi (Brid.) Mitt. (40%), Hylocomium splendens (Hedw.) Schimp. | 8.2 | 35.8 | 0.09 |
| Forest of mountain (FM) | |||||
| 213 | Skeletic Histic Podzol | Picea abies, Vaccinium myrtillus, Vacinium vitis-idaea, Pleurozium schreberi, Sphagnum | 6.6 | 27.1 | 0.08 |
| Mix forest (MF) | |||||
| 450 | Hyperskeletic Entic Podzol | Betula czerepanovii, Salix lapponum L., Sorbus aucuparia, Vaccinium myrtillus, Empetrum hermaphroditum Hagerup, Pleurozium schreberi, Hylocomium splendens, Dicranum | 11.4 | 27.5 | 0.11 |
| Forest tundra (FT) | |||||
| 470 | Hyperskeletic Albic Podzol | Picea obovata, Pinus sylvestris L., Betula czerepanovii, Betula nana L., Salix phylicifolia L., Empetrum hermaphroditum, Vaccinium myrtillus, Vaccinium uliginosum L., Arctous alpina (L.) Niedenzu, Pleurozium schreberi, Cladonia stellaris (Opiz) Pouzar & Vezda, Dicranum, Juncus trifidus L., Loiseleuria procumbens (L.) Desv.), Arctous alpina, Empetrum hermaphroditum, Lycopodium lagopus (Laest. ex Hartm.) Zinserl. ex Kuzen., Pinguicula villosa L. и дp.) | 4.2 | 29.5 | 0.12 |
| Biotypes of Microorganisms | TF | FM | MF | FT |
|---|---|---|---|---|
| Acetobacter-Rhodobacter group | 6.05 | 8.52 | 11.49 | 9.36 |
| Aeromonas hydrophila | 0.00 | 39.50 | 14.94 | 8.61 |
| Agrobacterium radiobacter | 3.56 | 0.00 | 4.11 | 1.86 |
| Caulobacter | 30.66 | 173.12 | 52.74 | 0.72 |
| Pseudomonas fluorescens | 4.01 | 4.66 | 3.54 | 2.58 |
| Pseudomonas putida | 0.54 | 0.89 | 0.91 | 6.90 |
| Pceudomonas vesicularis | 0.54 | 0.72 | 0.61 | 0.00 |
| Methylococcus/Clostridium sp. | 9.50 | 0.00 | 0.00 | 8.71 |
| Sphingobacterium spiritovorum | 0.35 | 1.26 | 0.94 | 0.48 |
| Sphingomonas adgesiva | 0.81 | 1.98 | 1.05 | 0.84 |
| Sphingomonas capsulata | 0.83 | 2.09 | 2.25 | 1.93 |
| Xanthomonas sp. | 1.64 | 1.46 | 1.53 | 1.46 |
| Specific iron-reduction bacteria | 0.36 | 0.00 | 0.19 | 0.15 |
| Acetobacterium sp. | 2.02 | 2.05 | 1.04 | 0.37 |
| Bacillus subtilis | 2.60 | 2.70 | 4.78 | 3.07 |
| Bacillus sp. | 10.56 | 9.24 | 11.95 | 5.28 |
| C.pasteureanum | 9.50 | 0.00 | 0.00 | 8.71 |
| C.perfringens | 0.10 | 0.04 | 0.04 | 0.05 |
| Eubacterium lentum | 1.39 | 0.97 | 1.00 | 0.82 |
| Clostridium OPA * | 1.82 | 0.00 | 0.33 | 2.46 |
| Butyrivibrio 1-4-11 | 0.00 | 0.12 | 0.49 | 0.67 |
| Butyrivibrio 7S-14-3 | 3.48 | 4.61 | 4.21 | 2.46 |
| Bifidobacterium sp. | 0.45 | 0.00 | 0.00 | 0.16 |
| Corynebacterium sp. | 6.47 | 32.97 | 9.18 | 1.08 |
| Mycobacterium sp. | 0.00 | 0.00 | 0.00 | 0.00 |
| Mикoбaктepии пo 10Me18 | 15.39 | 0.00 | 0.93 | 1.64 |
| Nocardiopsis | 10.83 | 20.49 | 2.34 | 0.00 |
| Propionibacterium sp. | 9.09 | 2.04 | 1.75 | 0.00 |
| Propionibacterium freudenreichii | 6.60 | 0.00 | 0.00 | 0.00 |
| Pseudonocardia sp. | 2.94 | 2.26 | 3.40 | 2.14 |
| Rhodococcus equi | 54.34 | 31.02 | 9.46 | 3.89 |
| Rhodococcus terrae | 8.55 | 4.72 | 6.01 | 6.41 |
| Streptomyces-Nocardiopsis | 13.19 | 13.21 | 21.31 | 7.66 |
| Bacteroides fragilis | 0.26 | 0.00 | 0.00 | 0.00 |
| Bacteroides hypermegas | 0.05 | 0.05 | 0.06 | 0.06 |
| Bacteroides ruminicola | 0.36 | 0.46 | 0.54 | 0.42 |
| Cytophaga sp. | 0.41 | 0.73 | 1.33 | 1.33 |
| Ruminococcus sp. + ** | 6.22 | 4.26 | 5.00 | 7.07 |
| WolInella-Acholeplasma-Roseomonas-Burkholderia | 1.62 | 3.75 | 4.41 | 2.40 |
| Micromonospora sp. | 2.74 | 1.75 | 0.99 | 1.17 |
| Chlamydia sp. | 0.00 | 0.00 | 0.00 | 0.00 |
| Bradyrhizobium, Mezorhizobium | 14.14 | 12.98 | 4.44 | 3.66 |
| Micrococcus/Arthrobacter sp. | 7.90 | 8.03 | 6.25 | 3.70 |
| Eubacterium sp. | 0.20 | 0.17 | 0.15 | 0.03 |
| Nocardia carnea | 3.89 | 5.79 | 7.33 | 5.46 |
| Actinomadura roseola | 1.02 | 1.00 | 4.20 | 3.20 |
| Aspergillus sp. | 4.01 | 20.81 | 7.37 | 10.43 |
| Enterobacteriaceae (ceмeйcтвo) | 2.08 | 0.00 | 0 | 0 |
| Butyrivibrio 1-2-13 | 3.11 | 0.58 | 0.79 | 0.32 |
| Enterococcus | 0.49 | 1.24 | 0.85 | 0.56 |
| Riemirella | 0.00 | 10.10 | 7.79 | 6.2 |
| Ochrobactrum | 0.83 | 1.03 | 3.81 | 0.47 |
| The amount | 267.5 | 433.4 | 228 | 137 |
| Fungy on 18:2, mkg/g | 226 | 524 | 257 | 401 |
| Yeasts on 10h16, cells/g × 106 | 0.06 | 0.02 | 0.02 | 0.01 |
| Protozoa | 0.00 | 21.00 | 8.75 | 6.98 |
| Eucariotes | 0.00 | 0.00 | 0.00 | 0.01 |
| Planta | 3.97 | 11.31 | 6.79 | 3.80 |
| N° | Bacterial Genus | Research Sites, N° | |||
|---|---|---|---|---|---|
| TF | FM | MF | FT | ||
| Proteobacteria | |||||
| 1 | Acetobacter | 3.14 | 2.50 | 6.75 | 10.38 |
| 2 | Aeromonas | 0.00 | 11.61 | 8.78 | 9.55 |
| 3 | Agrobacterium | 1.85 | 0.00 | 2.41 | 2.07 |
| 4 | Caulobacter | 15.91 | 50.90 | 30.98 | 0.80 |
| 5 | Methylococcus | 4.93 | 0.00 | 0.00 | 9.66 |
| 6 | Pceudomonas | 2.64 | 1.84 | 2.97 | 10.51 |
| 7 | Sphingomonas | 0.85 | 1.20 | 1.94 | 3.07 |
| 8 | Xanthomonas sp. | 0.85 | 0.43 | 0.90 | 1.62 |
| 9 | Specific iron-reduction bacteria | 0.19 | 0.00 | 0.11 | 0.16 |
| Firmicutes | |||||
| 10 | Acetobacterium | 1.05 | 0.60 | 0.61 | 0.41 |
| 11 | Bacillus | 6.83 | 3.51 | 9.83 | 9,26 |
| 12 | Clostridium | 5.93 | 0.01 | 0.22 | 12.44 |
| 13 | Eubacterium | 0.72 | 0.28 | 0.58 | 0.91 |
| Actinobacteria | |||||
| 14 | Bifidobacterium | 0.23 | 0.00 | 0.00 | 0.18 |
| 15 | Butyrivibrio | 1.80 | 1.39 | 2.76 | 3.47 |
| 16 | Corynebacterium | 3.36 | 9.69 | 5.39 | 1.20 |
| 17 | Propionibacterium | 8.14 | 0.60 | 1.03 | 0.00 |
| 18 | Pseudonocardia | 1.53 | 0.66 | 2.00 | 2.37 |
| 19 | Rhodococcus | 32.64 | 10.51 | 9.09 | 11.43 |
| 20 | Streptomyces | 6.85 | 3.89 | 12.52 | 8.50 |
| Bacteroidetes | |||||
| 21 | Bacteroides fragilis | 0.14 | 0.00 | 0.00 | 0.00 |
| 22 | Bacteroides hypermegas | 0.02 | 0.02 | 0.04 | 0.07 |
| 23 | Bacteroides ruminicola | 0.19 | 0.14 | 0.32 | 0.47 |
| 24 | Cytophaga sp. | 0.22 | 0.21 | 0.78 | 1.47 |
| Total bacterial cells number × 106 | 192.68 | 340.11 | 170.24 | 90.17 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medvedeva, M.V.; Bakhmet, O.N. Changes in the Microbiological Properties of Soils along the Gradient of the Altitude Zone of Mount Kivaka in Eastern Fennoscandia, Russia. Forests 2022, 13, 849. https://doi.org/10.3390/f13060849
Medvedeva MV, Bakhmet ON. Changes in the Microbiological Properties of Soils along the Gradient of the Altitude Zone of Mount Kivaka in Eastern Fennoscandia, Russia. Forests. 2022; 13(6):849. https://doi.org/10.3390/f13060849
Chicago/Turabian StyleMedvedeva, Maria V., and Olga N. Bakhmet. 2022. "Changes in the Microbiological Properties of Soils along the Gradient of the Altitude Zone of Mount Kivaka in Eastern Fennoscandia, Russia" Forests 13, no. 6: 849. https://doi.org/10.3390/f13060849