1. Introduction
Since 2012, there has been extensive mortality of the Norway spruce (
Picea abies L. Karst.) due to insect attacks [
1,
2]. The most harmful pest is the European spruce or eight-toothed bark beetle (
Ips typographus L.)—a secondary pest that primarily affects weakened and depressed trees. A change in the hydrological regime due to global warming and acid rain facilitated the weakening of spruce. It should be borne in mind that the spruce in the Białowieża Forest in Poland (which means further Forest) is located in the south of the northeastern part of the range [
1]. Therefore, it is especially susceptible to changes in hydrology. The bark beetle actively attacks even healthy and robust trees during mass reproduction. This negative phenomenon occurs in the Forest, in Poland, including areas with rich soil conditions that fully meet the ecological and biological requirements of
A. alba [
3,
4,
5]. This species is natural to the Forest but has survived only in a small refugium, in the Tisovik tract in the Belarusian part of the Forest [
6,
7]. It lies 500 km northeast of the edge of the continuous range of silver fir in Poland and 120 km from the nearest natural island habitat in the Jata Reserve [
8]. At present, it is the rarest coniferous species in the Forest.
Taking into account the autochthonous nature of the silver fir from Tisovik, it is reasonable to assume that the
A. alba does not pose a threat to biodiversity, but, on the contrary, enhances it. This is unlike, for example, a very aggressive invasive species,
Quercus rubra (L.), which displaces
Q. robur (L.) and thus poses a real threat to the stability of forest ecosystems [
9,
10].
It is interesting to study the possibility of including silver fir as a candidate for artificial reforestation in the Forest and adjacent forest districts. According to our previous research, silver fir has exhibited high growth rates and vigor in stands of different ages and origins [
11,
12]. These performance measures justify establishing a seed base of silver fir for targeted afforestation of those areas freed up by spruce mortality.
A half-sib family is a collection of plants obtained from seeds from free pollination from one tree. In this context, the study of the unique gene pool of autochthonous silver fir, presented in the form of half-sib families (what we understand as “provenance”) in the Hajnówka Forest District (FD), is necessary to identify the most promising genotypes. These promising genotypes could be preferred seed sources, especially for establishing seed plantations. Therefore, the studies of the first stage [
11,
12], continued in this research, were conducted using traditional forest mensuration and phenotypic analyses.
It is essential to note that despite its small size—approximately 20 trees at present [
7,
13,
14]—the autochthonous population of silver fir in the Tisovik tract is similar in the degree of polymorphism and heterozygosity (based on isoenzymatic and DNA analyses) compared to the populations from the main distribution area in the Carpathians [
15]. The list of publications on the genetics of fir in the Carpathians, without comparison with the population from Tisovik, is quite extensive [
16,
17,
18]. Unfortunately, only this publication provides a genetic comparison of the population from Tisovik with populations from the main area. This small population shows no inbreeding effect [
15], suggesting the value of a detailed genetic study of half-sib families, especially in selecting candidates for preferred seed trees. A study by Pawlaczyk et al. [
19] showed an excess of heterozygotes on a half-sib plantation in the Hajnówka FD, which is the offspring of such a small population as the fir group in the Tisovik tract. The authors attribute this to a “bottleneck effect” and assume that pollination of the remnant firs in Tisovik could have been carried out, among other things, from two stands (artificial, of unknown geographical origin) in the Polish part of the Białowieża Forest. Based on genetic analysis, Mejnartowicz [
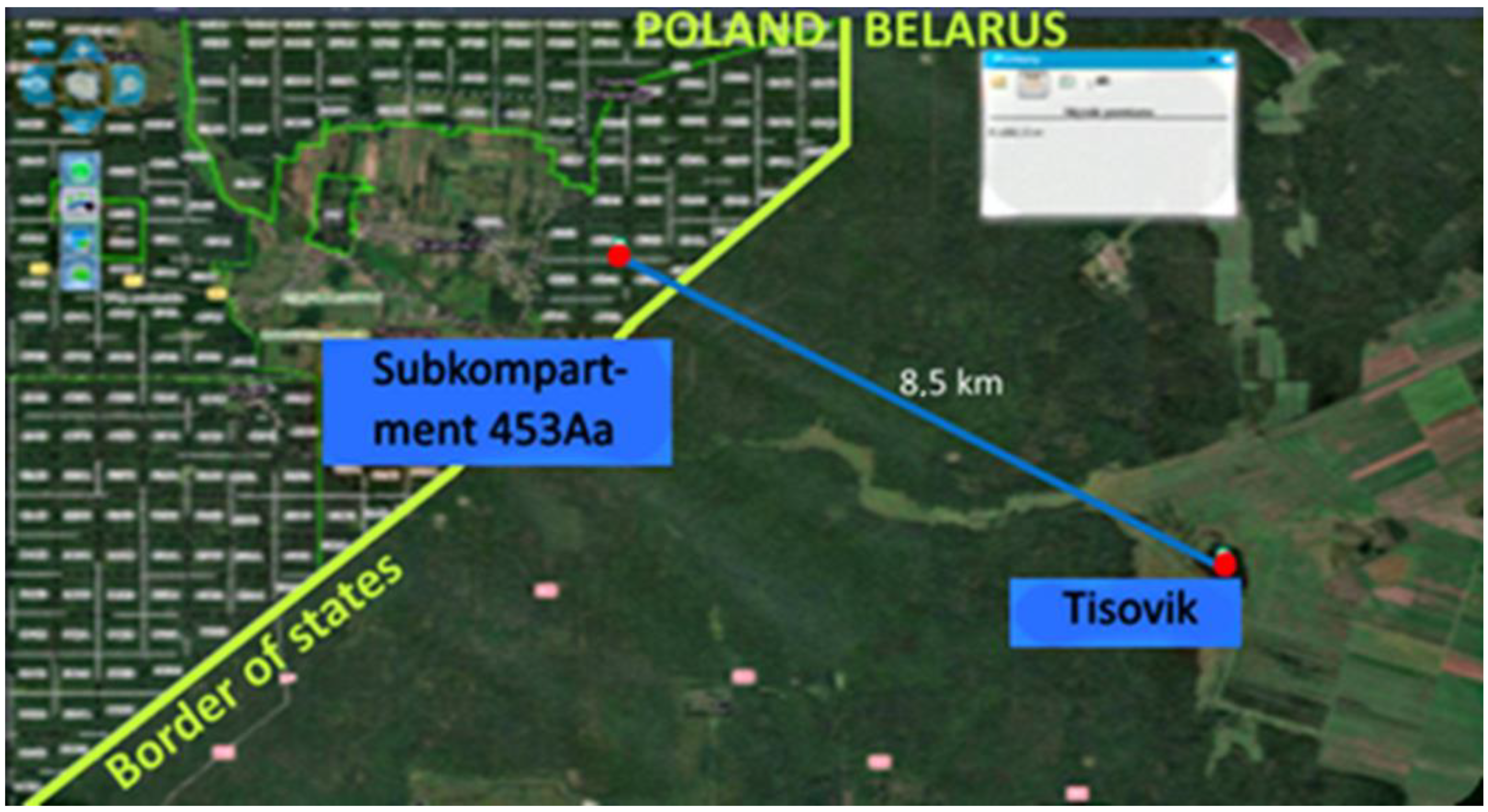
20] became convinced that these two stands are not descendants of autochthonous fir from the Tisovik tract (even though the nearest stand is located 8.5 km in a straight line from this refuge). If the assumption of Pawlaczyk et al. [
19] is correct, then not only the progeny of local cross-pollinated silver fir from Tisovik is represented in the half-sib plantation, but also individuals that arose as a result of gene flow together with pollen from these two stands of unknown origin, which are located in two forest subcompartments, 453Aa and 498 Ci of the Białowieża FD. We were unsuccessful in finding documents on the history of their appearance (between 1920 and 1939) in Polish archives, likely due to the destruction occurring during the Second World War. They might exist in some archives in Belarus, which are not available to us at the moment.
It would be logical to assume that there is a hereditary high degree of differentiation of DBH (diameter at breast height) between the families of the studied silver fir half-sibs and within families. Confirming or refuting this assumption was one of our research objectives, presented below. Note that at this stage, the genetic factor is conventionally represented as the membership of trees in one of the families of half-sibs (provenance). In addition, only the next step will be a study based on the evaluation of DNA polymorphism of the identified best specimens for this phenotype (DBH) trait.
Competitive interactions between trees occur at all stages of the forest stand dynamics, but they are the most pronounced at the second—stem—exclusion stage of forest formation [
21]. The density and spacing of the trees determine the competitive influence experienced by any single tree. These interactions, in turn, influence the spatial structure of the stand, affecting both productivity and species diversity of forest communities. Optimising the spatial structure of forest stands as a function of age classes, and taking into account the biological characteristics of the breed is one of the most effective methods of increasing a stand’s productivity and stability. Thus, regulating the spatial distribution of trees is the most important tool for managing artificial forest ecosystems [
22].
The development of stand growth modeling raises the question of how to simulate the initial states of these models and, in particular, how to simulate realistic spatial structures [
23]. The parameters of the spatial structure of forest stands determine and characterize them quantitatively. However, only understanding how to change the spatial structure effectively is crucial for forest management [
24]. Knowledge of the spatial structure of stands can be used to develop silvicultural regimes to increase the area of semi-natural forests established as even-aged plantations [
25]. Growth models of stands considering their spatial characteristics can predict productivity and evaluate changes in forest fund parameters and are recommended for use in forestry [
26].
Thus, the aim of this study was a comparative evaluation of the influence of the provenance factor and spatial structure on the growth of trees in a 22-year-old artificial stand of autochthonous silver fir from the Tisovik tract.
The objectives of this study were as follows:
4. Discussion
We note that the established significant variability of average DBH of half-sibs (
Table 1) occurs in the absence of differences in edapho-climatic conditions, characteristics of the forest cultivated area, age, and type of planting material, timing, method of establishment, and maintenance of the plantation. The provenance factor’s influence is reflected by the membership of trees in a particular half-sib family. However, within each half-sib, a certain amount of phenotypic variability occurs. This variability might be due, among other things, to possible cross-fertilization with the influence of paternal components, which may differ genetically. Moreover, as noted above (see section “Introduction,” [
19]), trees not only from Tisovik but also from stands of unknown origin in the Polish part of the Forest may participate in pollination. The closest location (subcompartment 453Aa) is 8.5 km from the refugium (
Figure 6). If this is the case, then given the growing aridity of the climate, an increase in genetic mixing is regarded as a positive factor. However, the opposite process is also possible—with pollen from Tisovik pollinating the macrostrobils on trees in sub-compartment 453Aa.
As shown above, the parameters of phenotypic variability differ in different half-siblings, which may evolve into actual genetic differences. This potential for future genetic variability suggests the need for maximum conservation of each tree—a potential source of heritable valuable economic traits—in thinning.
Note that the biological features of fir fully contribute to this outcome. On the one hand, due to the pronounced taproot system of fir, the growing space of individual trees is relatively small. On the other hand, this species is characterized by exceptional shade tolerance [
44,
45,
46]. These results suggest that managers reduce harvest impacts of future silvicultural and increase genetic opportunities by minimizing felling intensity at the cleaning stage. For example, in 2015, the marked tree in
Figure 7 was initially designated for felling because of its reduced growth compared to the neighboring trees. However, since this experimental site was assigned to the UNESCO protected zone II, where any felling is prohibited, this tree was preserved. This “reprieve” was borne out by its subsequent performance similar to neighboring trees by 2021.
In this regard, we also note that, according to Goncharenko [
47], the fir in Tisovik “despite its isolation and small size, has not yet experienced the influence of inbreeding, and although only 20 trees were preserved, as a result of crosses (cross-pollination) between them, one can obtain 13,122 genetically different offspring”.
Nevertheless, the issue of the manifestation of inbreeding in the Tisovik tract should be approached more carefully and, perhaps, not so optimistically [
48]. Because the population, represented by only about 20 trees [
7,
12,
47], is very small, and they live in a very small area. In addition, perhaps it is the effect of inbreeding that can explain the lowest survival rate in family No. 5—28.3 (only 73 trees out of 258 survived) [
12], as well as the significant differences in diameter both between sibs and within sibs, as can be seen from our data. Although, in the latter case, the effect of inbreeding, even if it takes place, is manifested against the background of the natural process of biological differentiation of trees (Kraft classes or social classes of trees position in canopy [
48]).
Moreover, for an exceptionally shade-tolerant fir, it is characteristic that not only trees of IV, but even sometimes Va of Kraft classes are not doomed to fail. Most of them are not in a state of gradual mortality. On the contrary, their condition corresponds to a live plant, only with very slow growth rates. In addition, this is precisely why the statistical analysis showed a large standard deviation and, accordingly, the coefficient of variation of DBH (
Table 1,
Table 2 and
Table 4). In natural stands, when the dominant trees approach the climax stage, it is the representatives of the lower tiers that begin to take over their role [
44,
45,
46].
To obtain a genetically richer silver fir population originating from the seeds of the planned vegetative seed plantation created by grafting, we must first select the best phenotypes from all sibs.
Therefore, when creating it, you should not focus only on the top nine trees identified at the moment. According to Barzdajn [
49], who was involved in the restitution of silver fir in Sudety, clones should number at least 100. This conclusion implies that phenotypic studies must be repeated periodically to identify the best trees (
Table 10). In our opinion, a frequency of 5 years will be necessary. These remeasurements will make it possible to record the emergence of new candidates for superior trees since, as we noted earlier, there is a pronounced positive dynamics of tree growth [
12] (
Figure 2 and
Figure 3). Additionally, two more plots are awaiting their turn at this object. The most critical condition for the success of future vegetative seed plantations is the maximum representation of all the best phenotypes from all half-sibs families.
The closest positive correlation of DBH occurs with the distance to the nearest neighboring tree in the southern direction (0.69) and the lowest (0.45) in the eastern direction (
Table 9). These results suggest that the creation of conditions for an increase in illumination in the stand without excessive felling of trees (i.e., an increase in the distance to a neighboring tree in the southern direction with the highest intensity of solar radiation) has a positive effect on the growth of trees (an increase in their DBH). As it turns out,
A. alba, which is a classically shade-tolerant species, responds positively to improving light conditions under these conditions, even at a young age.
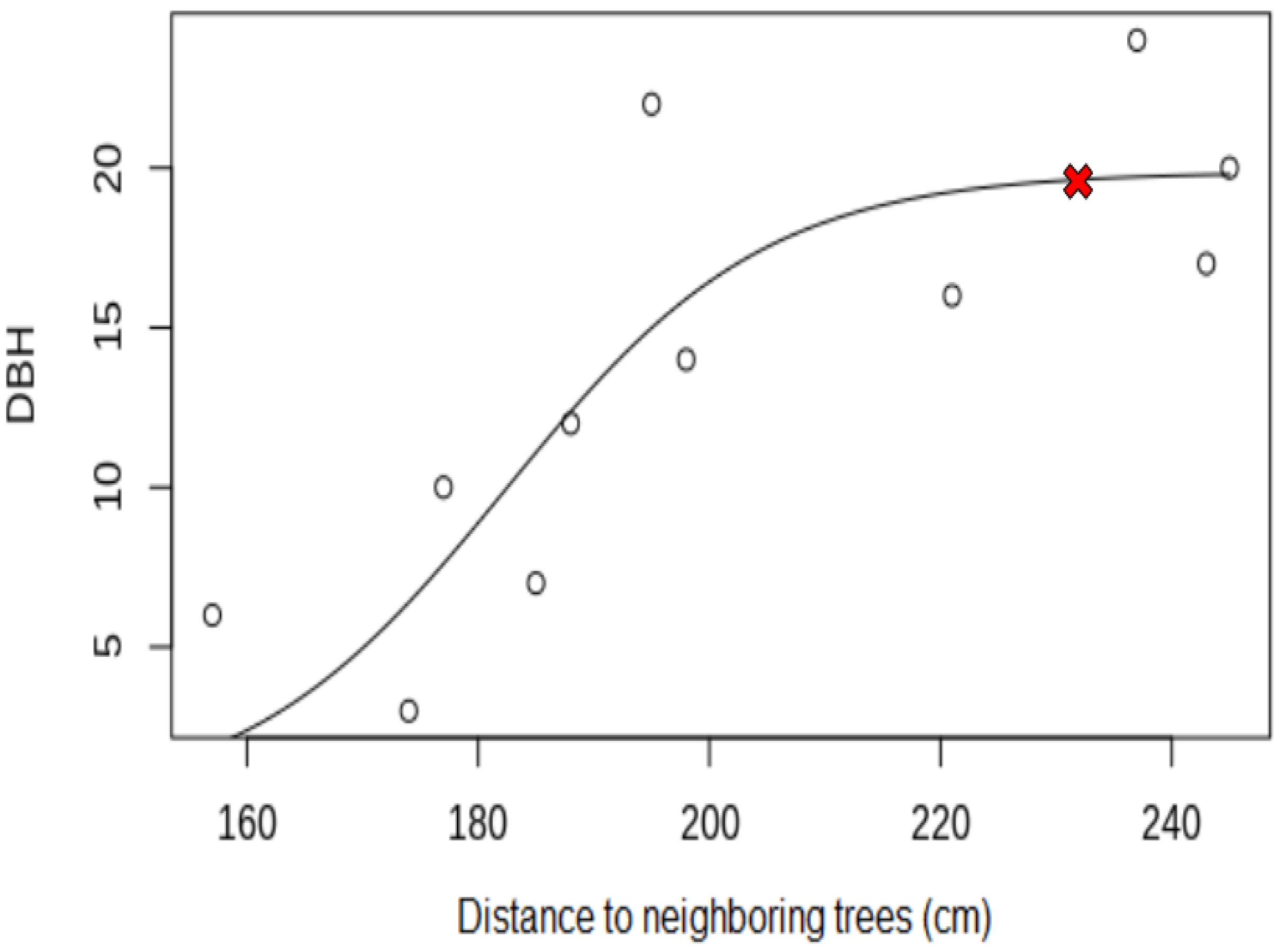
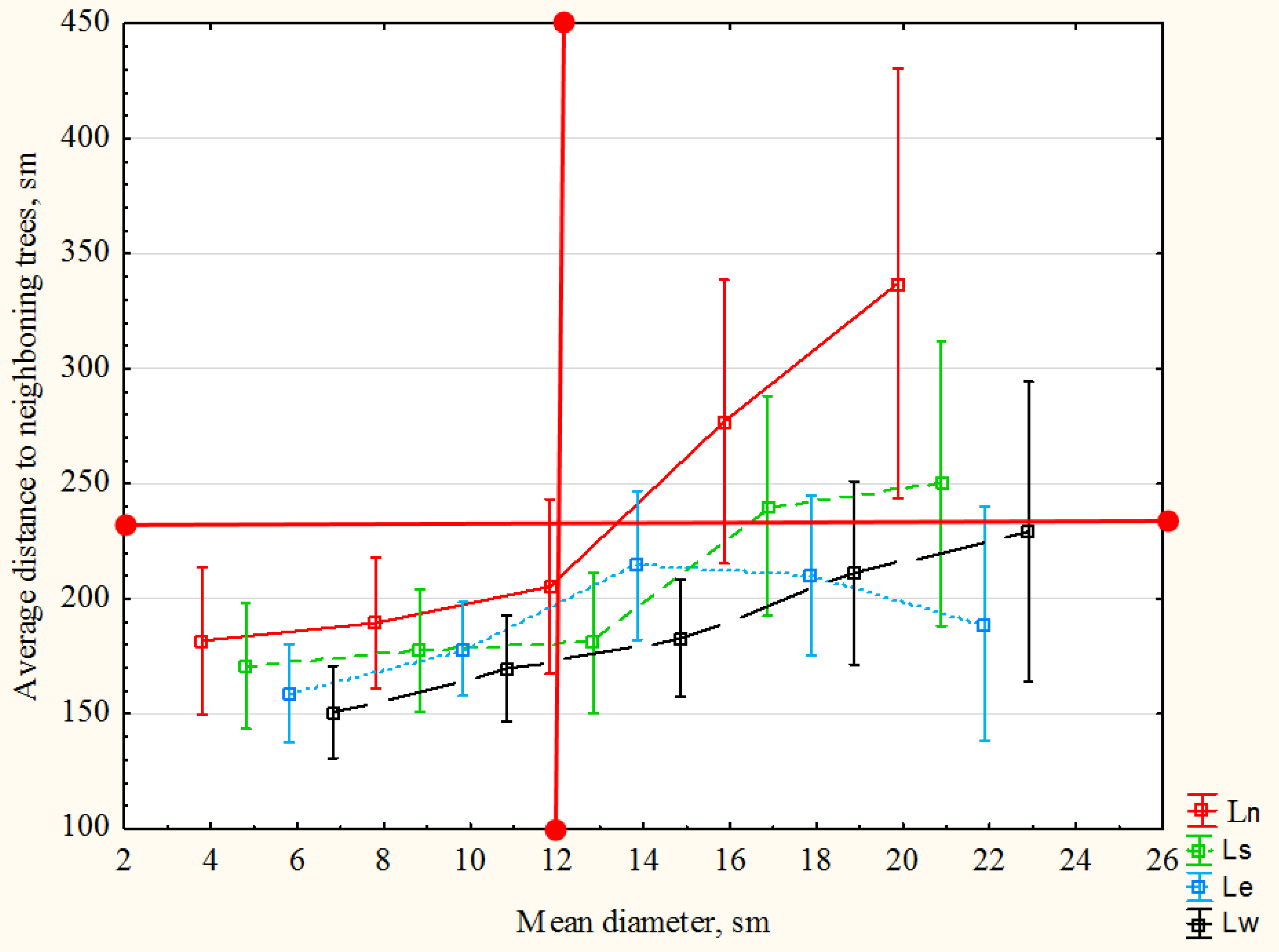
The relationship between the DBH and the average distance to neighboring trees (
Figure 8) suggests a certain “threshold value” of the average distance between trees in a stand of a given age. Beyond this threshold, further thinning will not lead to an increase in the productivity of the stand nor an improvement in its general condition. At the moment, this threshold indicator is 2.3 m (
Figure 8). However, this distance is not permanent and will change with the age of the stand.
Thus, 2.3 m is a “threshold,” which we believe determines the end of the influence of spatial structure on DBH increase. This factor ceases to be limiting [
50,
51]. Even though the curve of the graph continues to move very slightly upward, from a silvicultural point of view, a further increase in the average distance will lead to an unnecessary decrease in the density of the stand, a reduction in its overall growth and volume, and underutilization of forest area by root systems of trees. Furthermore, an excessive increase in the sparseness of the stand negatively affects the light regime of the shade-tolerant fir. A simple calculation shows that with an increase in the average distance between trees from 2.3 m to 2.5 m (i.e., when the red cross in
Figure 8 moves to the right), it is necessary to additionally remove about 300 trees from a hectare, which will lead to a decrease in stand density by 0.1 units.
The same cautious approach to tree cutting should be followed in the future. Of course, the parameters of the spatial structure will change, and this is the subject of future research. However, the principles of silvicultural care remain as follows: (a) only obvious outsiders are cut down (trees at the left-hand side of the distribution curve), and (b) tree stand care is carried out using more frequent, low-intensity entries rather than infrequent intensive thinnings.
In this case, the presence of individuals with an increased DBH and indicators of spatial structure exceeding the “threshold” can only be interpreted as a consequence of the influence of a provenance factor. As shown in
Table 10, in seven out of nine trees (78%) with the largest DBH (candidates for plus trees), the average distance to neighbors exceeds 2.3 m. A genetic study of these trees, planned for the future, will allow a more substantiated statement on the validity of this assumption.
Following the methodological approach of Rogozin [
41], we use half the average distance between trees (1.15 m) as the radius of a sufficient “feeding area” [
40,
41] or “growth area” [
38,
39]. As a result, we get that the growth area of each tree is 4.15 m
2, and the density of the stand is 2410 pcs/ha (where it is 3186 pcs/ha at present). It is necessary to approach this indicator, as already mentioned, by selecting the weak intensity according to the grass-root method, removing especially the trees of the 2 and 4 cm diameter classes. In this way: (1) the biology of the species (shade tolerance) is taken into account, (2) the vertical structure is formed, and (3) the gene pool of
A. alba is preserved.
In a situation where a pronounced differentiation is currently taking place in the forest stand (
Table 1), the future of the dominant trees, especially advantage tree candidates, is of interest. Can their growth be improved? It was found that there is an unequal dependence of the change in DBH on the distance to the nearest tree in the direction of the different world sides (
Figure 9).
The starting point for evaluating this dependence is the intersection of the two perpendiculars on the axis of this diagram. The vertical perpendicular corresponds to a DBH of 12 cm, representing the diameter class above the average stand DBH (9.97 cm, [
12]). The horizontal line corresponds to the “threshold” of the spatial structure (see above).
The thinning required to increase the DBH, expressed as the distance to the nearest tree in the various directions, takes the following form (in decreasing order): north, south, west, and east. That is, by focusing on the removal of trees in the rows (north-south direction) and at the same time cutting less intensively in the direction between the rows (west-east), it is possible to influence the increase in DBH of the dominant trees. This opportunity arises from the trees’ initial very dense arrangement in the rows (1 m, and sometimes less,
Table 3).
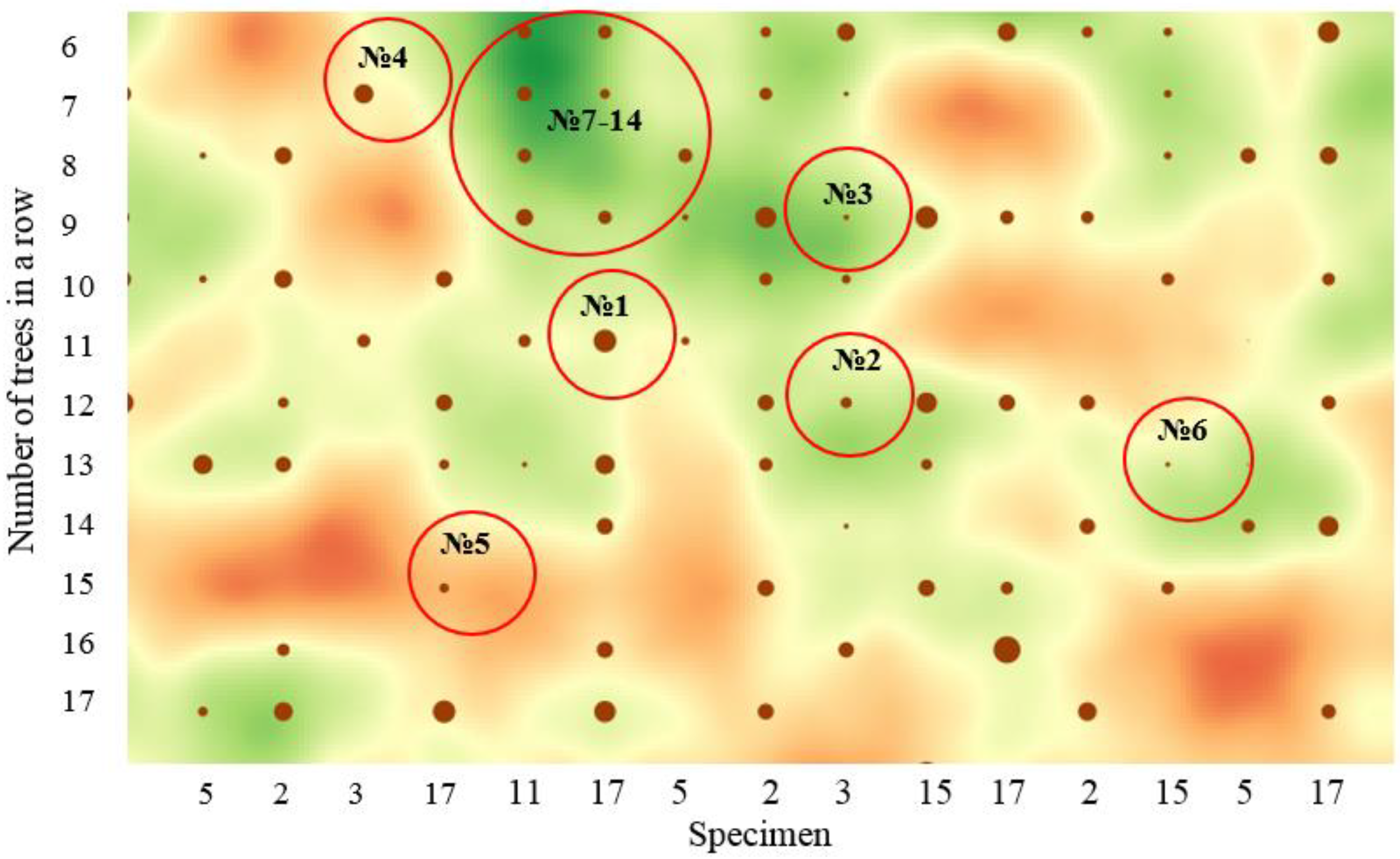
The complexity and ambiguity determining the influence of the spatial structure are illustrated by this analysis of the randomly located sample (
Figure 10).
As can be seen, the planting lines are still visible. However, the initially regular (systematic) placement of trees according to the 1.3 × 1 m design becomes uneven. Biogroups of trees (highlighted in green) and gaps in the forest stand (highlighted in orange) begin to form. Note that from a bird’s eye view, the unevenness of the canopy of the crowns, due to their development, is not yet so apparent (
Figure 2). What may be the cause of the uneven spatial structure of the stand? First of all, we note the possible accretion of fir roots [
52] and the formation, thus, of microgroups, each of which has a common root system. Some plants died due to late spring frosts in the early years (see Korczyk [
53]). Perhaps the negative influence of excessive solar radiation was also uneven in different parts of the plot. In addition, this non-uniformity in the distribution of plants persisted subsequently (we did not find any information in the literature about the addition of crops). After thinning, it may have even increased somewhat under the influence of the subjective factor (selection of trees for cleanings). However, the root causes of this phenomenon (uneven distribution of trees) are not yet known [
41]. Therefore, we can only note the appearance of fundamentally new attributes of the morphogenesis of the stand: biogroups (microcenoses) and gaps in the stand.
From the analysis of the fragment of the spatial model (
Figure 10), it follows that the trees with neighbors that are behind in growth or have a greater distance to them also have a larger DBH (Nos. 1, 4). Conversely, the DBH of the focus trees decreases with decreasing distance to neighboring trees or an increase in the neighboring trees’ DBH (Nos. 2, 3). These two situations are well explained by the weakening and strengthening of intraspecific competition. However, then, do we explain the same DBH of the trees Nos. 7–14 growing in the biogroup? Perhaps there is already the partnership effect (group effect) [
41] (see
Figure 11). In addition, in this context—perhaps there is no need to strive for an even distribution of trees [
54]? In nature,
A. alba is usually found in an admixture of the main tree species in groups because it has relatively heavy seeds [
46]. In addition, as it turns out, it grows better with artificial reproduction in groups; in particular, it is highly resistant to biotic and abiotic factors [
55].
And what influenced the insignificant DBH of the trees Nos. 5 and 6 growing in the gaps? After all, the competition factor is not the limiting influence at present. Perhaps they were suppressed before the late cleaning and still cannot rebuild? Or is this perhaps the result of genetic differentiation? Approximately the same questions arise in analyzing any other fragment of a half-sib plantation.
It must be admitted that the above analysis is objectively a simplified approach that does not take into account other complex influences that are challenging to determine and not consistently observable by managers and scientists (allelopathy, root competition, local specificity of the soil, the state of the plants before planting and late cleaning, etc.). According to Rogozin [
41], the proportion of unknown factors that determine the size of trees in old pine stands can reach 60%.
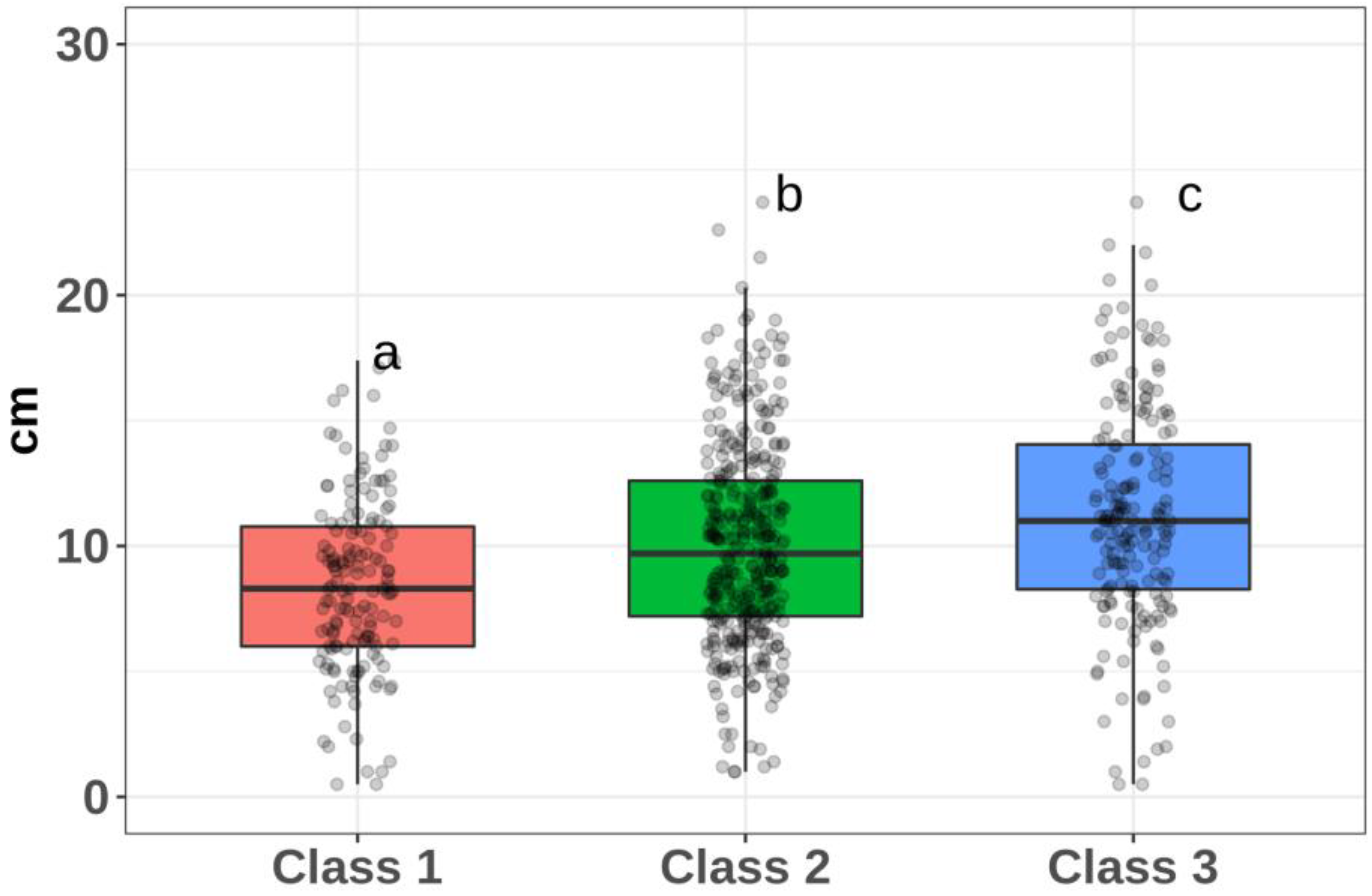
When assessing the influence of the provenance factor on the diameter, it was shown that all three classes are statistically significantly different from each other in terms of the p-level of significance. In addition, there is a pronounced dependence of the DBH on the provenance factor (
Figure 4).
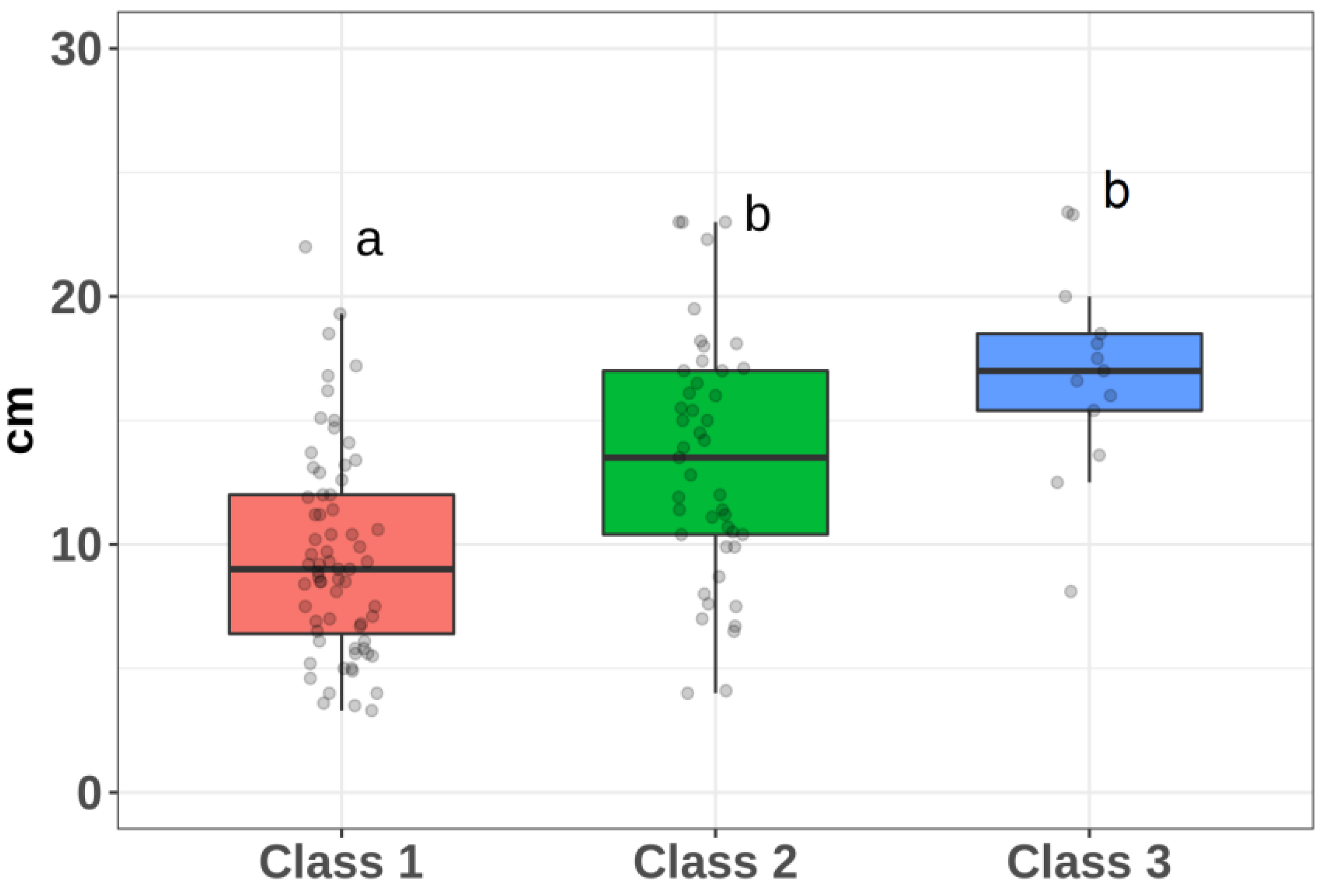
A different situation occurs when assessing the influence of spatial structure. While classes 1 and 2, as well as 1 and 3, differ significantly from each other (
p < 0.01), the difference between classes 2 and 3 is not significantly significant (
p = 0.12). Nevertheless, we note that the tendency to increase in diameter from class 2 to class 3 is visually apparent (
Figure 5).
In light of the above, there is a reason to once again turn to the conclusion illustrated in
Figure 8. The distance between the trees (the density of the stand), which provides them with favorable conditions at a given age is 2.3 m, the middle of the class interval No. 2 (
Table 4). The silvicultural interpretation of the statistically insignificant difference between classes 2 and 3 is the same: a borderline acceptable distance between trees—“threshold”—exists. We found the same result using two different methods, supporting our conclusions.
5. Conclusions
The study results give reason to believe that at this particular age, provenance has a more significant influence on the growth of the trees in DBH throughout the whole range of its variation than has the spatial structure. However, it was also found that the importance of the last factor should not be discounted, especially in the range of small and medium distances between trees (classes 1 and 2), where it can determine their parameters.
Free pollination (father unknown) of a particular mother tree in the Tisovik tract is the basis for the emergence of the half-sibs we studied. In this respect, each of the trees on the plantation of European silver fir, even within the same family, is genetically differentiated and may differ in characteristics to one degree or another. Therefore, to maximize future silvicultural and genetic opportunities, we should preserve as many trees as possible.
The nine best trees were identified, which are the prototypes of plus trees. Their seeds and shoots can already find a practical use for the early creation of a seed base of autochthonous silver fir. By starting this work now, in an experimental stand of about 30 years, we will accelerate the emergence of seed plantations by at least 50 years.
Family No. 17 is of particular interest for further genetic study. A significant number of specimens were identified in this half-sib, with DBH significantly exceeding the average of the plantation. However, this does not mean we should concentrate only on this half-sib. On the contrary, all semi-sibs should be maximally represented in the seed base in order to maintain or even enhance the genetic diversity of future autochthonous silver fir forests.
The actual planting density at the time of the study was 3186 plants/ha, while the threshold value determined by us is 2410 plants/ha. This particular hold can be used in practical forestry when thinning silver fir trees at about 25–30 years. The stand should be “brought” to the threshold value gradually, performing regular fellings of low intensity and removing only obvious outliers.
In light of the catastrophic situation with Norway spruce in Białowieża Forest, studies on various aspects of growing local silver fir as an alternative are of undoubted scientific and practical value.


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}